Scaricare la presentazione
1
Potenziali di membrana; potenziale di riposo e d'azione.
Premesse indispensabili

2
MEMBRANA PLASMATICA - MEMBRANE DEGLI ORGANELLI
COMPOSIZIONE: bistrato fosfolipidico contenente enzimi, recettori, antigeni; proteine integrali ed estrinseche. LIPIDI: fosfolipidi, colesterolo, glicolipidi PROTEINE: asimmetria funzionale (pompe ioniche) TRASPORTO SENZA ATTRAVERSAMENTO DI MEMEBRANA : Fagocitosi, pinocitosi, esocitosi: formazione di vacuoli, fusione delle membrane anche fra vacuoli (possibile formazione di canali temporanei)
 TRASPORTO. SENZA ATTRAVERSAMENTO DI MEMEBRANA : Fagocitosi, pinocitosi, esocitosi: formazione di vacuoli, fusione delle membrane anche fra vacuoli (possibile formazione di canali temporanei)")
3
TRASPORTO MEDIATO DA PROTEINE
Più rapido; cinetica di saturazione; specificità; inibizione competitiva; possibile inibizione da altri fattori; TRASPORTO (DIFFUSIONE ) FACILITATO: non avviene contro gradiente (non consuma energia), ma è operato da proteine specifiche TRASPORTO ATTIVO: crea gradienti di concentrazione; consuma energia; trasporto attivo secondario (cotrasporto).
 FACILITATO: non avviene contro gradiente (non consuma energia), ma è operato da proteine specifiche. TRASPORTO ATTIVO: crea gradienti di concentrazione; consuma energia; trasporto attivo secondario (cotrasporto).")
4
DIFFUSIONE leggi: 1) (x)2 = 2Dt x = distanza t = tempo D = coefficiente di diffusione il tempo necessario per percorrere la distanza x è proporzionale a D; il tempo di diffusione è inversamente proporzionale al quadrato della distanza.

5
2) D = kT/(6r) (equazione di STOKES EINSTEIN)
kT = energia cinetica media r = raggio molecolare = viscosità del mezzo Il coefficiente di diffusione D è proporzionale alla temperatura e inversamente proporzionale al raggio della molecola e alla viscosità del mezzo. Per molecole con PM (peso molecolare) > a 300 D è inversamente proporzionale a ( 3 PM) e a (2 PM) per le molecole più piccole
 > a 300 D è inversamente proporzionale a ( 3 PM) e a (2 PM) per le molecole più piccole.")
6
DIFFUSIONE ATTRAVERSO UNA MEMBRANA
3) J= DA*(c/ x) (prima legge di FICK) J = velocità netta di diffusione D = coefficiente di diffusione A = area della membrana c = gradiente di concentrazione x = spessore della membrana La velocità netta di diffusione attraverso una membrana è proporzionale al coefficiente di diffusione, all'area della membrana e al gradiente di concentrazione; è inversamente proporzionale allo spessore della membrana.
 J= DA*(c/ x) (prima legge di FICK) J = velocità netta di diffusione. D = coefficiente di diffusione. A = area della membrana. c = gradiente di concentrazione. x = spessore della membrana. La velocità netta di diffusione attraverso una membrana è proporzionale al coefficiente di diffusione, all area della membrana e al gradiente di concentrazione; è inversamente proporzionale allo spessore della membrana.")
7
PERMEABILITA' DELLA MEMBRANA: alle molecole liposolubili; alle molecole idrosolubili.
Le molecole liposolubili sono permeabili in funzione della loro liposolubilità e seguono le leggi della diffusione. Le molecole idrosolubili attraversano la membrana solo se sono molto piccole (acqua), altrimenti la membrana è impermeabile, così come lo è per le molecole ionizzate. Per l'ingresso di queste molecole sono necessari trasportatori, in genere rappresentati da proteine (es. canali ionici, trasportatori di zuccheri e amino acidi).
, altrimenti la membrana è impermeabile, così come lo è per le molecole ionizzate. Per l ingresso di queste molecole sono necessari trasportatori, in genere rappresentati da proteine (es. canali ionici, trasportatori di zuccheri e amino acidi).")
8
PERMEABILITA' DELLA MEMBRANA: all’acqua.
La membrana cellulare è normalmente permeabile all’acqua, perché contiene specifiche proteine (aquaporine) che ne consentono il passaggio. In alcuni casi la permeabilità all’acqua è regolata: es. ormone antidiuretico (ADH)
 che ne consentono il passaggio. In alcuni casi la permeabilità all’acqua è regolata: es. ormone antidiuretico (ADH)")
9
OSMOSI: si verifica quando due ambienti contenenti molecole in soluzione sono separati da una membrana semipermeabile E’ semipermeabile una membrana che consenta il passaggio del solvente, non del soluto Qualunque membrana può comportarsi come semipermeabile nei confronti di alcune molecole e non di altre

10
membrana semipermeabile
soluzione solvente

11
10 mmHg

12
PRESSIONE OSMOTICA (PO) è la pressione che si oppone al passaggio del solvente ed è uguale e contraria alla forza di attrazione delle molecole del soluto su quelle del solvente. La PO è proporzionale alla differenza nel numero assoluto di particelle presenti in ogni ambiente, non al gradiente di concentrazione. A parità di concentrazione (in peso) sono più attive le molecole più piccole (sono più numerose) e quelle dissociate (gli ioni aumentano il numero di particelle).
 sono più attive le molecole più piccole (sono più numerose) e quelle dissociate (gli ioni aumentano il numero di particelle)..")
13
ESEMPIO PRATICO: Una soluzione somministrata in vena deve essere iso-osmotica rispetto al plasma, altrimenti può danneggiare le cellule del sangue. Soluzione salina (NaCl): 0.9% (9 grammi/litro) Soluzione glucosata: 5% (50 gramm/litro)
: 0.9% (9 grammi/litro) Soluzione glucosata: 5% (50 gramm/litro)")
14
Esistono anche soluzioni iperosmotiche, che vengono somministrate molto lentamente per non danneggiare il sangue e servono per riassorbire acqua dagli spazi interstiziali. L’acqua riassorbita verrà eliminata dai reni

15
CONSEGUENZE FISIOLOGICHE DEI FENOMENI OSMOTICI
Le membrane cellulari si comportano come semipermeabili nei confronti delle molecole di cui non consentono il passaggio (sono permeabili all'acqua): in un ambiente povero di soluti (ipoosmotico) le cellule si rigonfiano, mentre si raggrinzano in ambiente iperosmotico.
: in un ambiente povero di soluti (ipoosmotico) le cellule si rigonfiano, mentre si raggrinzano in ambiente iperosmotico.")
16
CONSEGUENZE FISIOLOGICHE DEI FENOMENI OSMOTICI
Nell’organismo avvengono continui ed importanti scambi di acqua. Le forze che spostano l’acqua sono: Pressione idrostatica Pressione osmotica

17
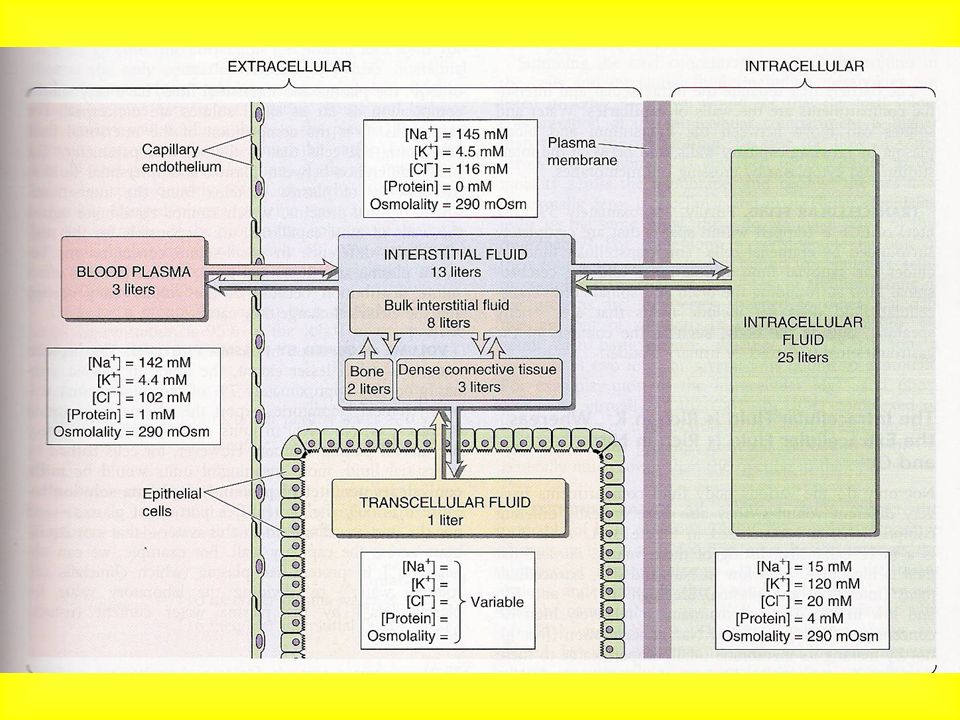
COMPARTIMENTI LIQUIDI DELL’ORGANISMO
Acqua 60% Intracellulare 60% (25 l) Extracellulare (15 l; comprende il plasma – 3.5 l) Transcellulare (membrane speciali: articolazioni, occhio …)
 Extracellulare (15 l; comprende il plasma – 3.5 l) Transcellulare (membrane speciali: articolazioni, occhio …)")
19
Fenomeni da comprendere e imparare:
Potenziali d’equilibrio Potenziali di membrana: Di riposo D’azione Fasi del potenziale d’azione Correnti ioniche durante il potenziale d’azione Mantenimento dell’omeostasi cellulare Conduzione del potenziale d’azione

20
Potenziali di membrana

21
EQUILIBRI IONICI E POTENZIALI TRANSMEMBRANARI POTENZIALE DI EQUILIBRIO
Le cellule viventi sono normalmente cariche elettricamente (differenza di potenziale), con l'esterno positivo rispetto all'interno Potenziale elettrochimico: creato da gradienti di concentrazione di una sostanza ionizzata (carica elettricamente), attraverso una membrana: il gradiente chimico spinge in una direzione e il gradiente elettrico nell'altra; quando le due forze si equivalgono, si raggiunge un POTENZIALE DI EQUILIBRIO
, con l esterno positivo rispetto all interno. Potenziale elettrochimico: creato da gradienti di concentrazione di una sostanza ionizzata (carica elettricamente), attraverso una membrana: il gradiente chimico spinge in una direzione e il gradiente elettrico nell altra; quando le due forze si equivalgono, si raggiunge un. POTENZIALE DI EQUILIBRIO.")
22
- - - - - - - - - - - - - - + + + + + + + + + + + + + + + + + + + +
Gradiente di concentrazione (chimico) Na+ Na+ K+ K+ K+ K+ K+ K+ Gradiente di carica (elettrico)
 Na Na K+ K+ K K+ K+ K+ Gradiente di carica (elettrico)")
23
P- K+ P- K+ P- K+ - + P- K+ + - = Potenziale di equilibrio

24
Il POTENZIALE DI EQUILIBRIO è prevedibile in base alla
EQUAZIONE DI NERNST: Ea-Eb = -(RT/zF)ln([Xa]/[Xb]) Ea-Eb = (RT/zF)ln([Xb]/[Xa]) Ea-Eb = differenza di potenziale R = cost. dei gas, F = cost di Faraday, T= temperatura assoluta, z = valenza. (RT/zF)ln = 60 log;
ln([Xa]/[Xb]) Ea-Eb = (RT/zF)ln([Xb]/[Xa]) Ea-Eb = differenza di potenziale. R = cost. dei gas, F = cost di Faraday, T= temperatura assoluta, z = valenza. (RT/zF)ln = 60 log;")
25
Potenziale di equilibrio Ea-Eb (mV) = 60/z log([Xb]/[Xa])
Il potenziale di equilibrio per uno ione (Ei) è proporzionale ad una costante e al logaritmo del rapporto fra le concentrazioni ioniche ai due lati della membrana
![Potenziale di equilibrio Ea-Eb (mV) = 60/z log([Xb]/[Xa])](http://slideplayer.it/slide/589594/2/images/25/Potenziale+di+equilibrio+Ea-Eb+%28mV%29+%3D+60%2Fz+log%28%5BXb%5D%2F%5BXa%5D%29.jpg "Il potenziale di equilibrio per uno ione (Ei) è proporzionale ad una costante e al logaritmo del rapporto fra le concentrazioni ioniche ai due lati della membrana.")
26
Esistono differenze di concentrazione fra liquido intra- ed extra- cellulare, in particolare:
sodio e cloro all'esterno, proteine e potassio all'interno

27
Ogni cellula avrà un potenziale di riposo (Em) che risulta dalla somma dei potenziali di equilibrio per ogni ione, calcolabile in base all'equazione di Nernst, ma corretto per la permeabilità (gi) della membrana allo stesso ione
 che risulta dalla somma dei potenziali di equilibrio per ogni ione, calcolabile in base all equazione di Nernst, ma corretto per la permeabilità (gi) della membrana allo stesso ione")
28
PERMEABILITA’ CONDUTTANZA (G)
Capacità di una membrana di far passare una determinata molecola (inverso = impermeabilità) CONDUTTANZA (G) Flusso effettivo di una molecola attraverso una membrana (dipende dalla permebilità e ne costituisce una misura)
 CONDUTTANZA (G) Flusso effettivo di una molecola attraverso una membrana (dipende dalla permebilità e ne costituisce una misura)")
29
Potenziale di membrana: equazione di GOLDMAN
Em = (gK/gt) EK + (gNa/gt) ENa + (gcl/gt) ECl; gt = gK+gNa+gCl Nella cellula a riposo gK è circa 10 volte maggiore di gNa e pertanto il potenziale è vicino a EK.
 EK + (gNa/gt) ENa + (gcl/gt) ECl; gt = gK+gNa+gCl. Nella cellula a riposo gK è circa 10 volte maggiore di gNa e pertanto il potenziale è vicino a EK.")
30
Le differenze di concentrazione sono generate da pompe metaboliche (in particolare Na/K-ATPasi), che possono essere parzialmente elettrogeniche (creano differenze di potenziale)
, che possono essere parzialmente elettrogeniche (creano differenze di potenziale)")
31
I canali ionici più importanti per i potenziali d’azione sono quello per il Na+ e quello per il K+.
In particolare, il canale per il Na+ è voltaggio-dipendente e si chiude progressivamente con la polarizzazione della membrana (negativa all’interno), quindi gNa è piccola a riposo
, quindi gNa è piccola a riposo.")
32
Poiché la pompa Na+/K+ è elettrogenica (porta fuori più Na+ e porta dentro meno K+), a mano a mano che si instaurano i gradienti di concentrazione, la membrana si polarizza diventando negativa all’interno (ci sono meno cariche positive). Questo fa chiudere i canali per il Na+ voltaggio-dipendenti e gNa è piccola

33
I canali per il K+ sono in larga misura non voltaggio-dipendenti e rimangono aperti nella membrana polarizzata, quindi gK è elevata (nella membrana a riposo)
")
34
POTENZIALE D'AZIONE E' la prima espressione dell'eccitamento della cellula eccitabile. E' un fenomeno con soglia E' un fenomeno tutto-o-nulla E' un fenomeno che si propaga rigenerandosi E' un fenomeno con un preciso decorso temporale.

35
Stimolo: fenomeno (generalmente elettrico), naturale o artificiale che può eccitare la cellula (se raggiunge la soglia), e provoca comunque una variazione del suo potenziale di riposo
, naturale o artificiale che può eccitare la cellula (se raggiunge la soglia), e provoca comunque una variazione del suo potenziale di riposo")
36
Stimolo sotto soglia: provoca variazioni locali del potenziale, che si propagano con decremento e diminuiscono nel tempo (costanti di spazio e di tempo).
.")
37
Stimolo = iniezione cariche positive
+30 -60 -80 mv depolarizzazione Tempo (millisecondi) Cellula a riposo Sotto soglia soglia Tutto-o-nulla Sopra soglia
 Cellula a riposo. Sotto soglia. soglia. Tutto-o-nulla. Sopra soglia.")
38
FASI DEL POTENZIALE D'AZIONE
potenziale di riposo stimolo efficace (talvolta chiamato prepotenziale; spesso non distinguibile) salita rapida con inversione del potenziale (spike) ripolarizzazione Iperpolarizzazione postuma.
 salita rapida con inversione del potenziale (spike) ripolarizzazione. Iperpolarizzazione postuma.")
39
Le fasi del potenziale d’azione
-60 -80 +30 mv Tempo (millisecondi) ENa EK Potenziale a punta (spike) ripolarizzazione prepotenziale iperpolarizzazione Potenziale di riposo Potenziale di riposo
 ENa. EK. Potenziale a punta. (spike) ripolarizzazione. prepotenziale. iperpolarizzazione. Potenziale di riposo. Potenziale di riposo.")
40
MECCANISMI IONICI PREVALENTI
a riposo: membrana (quasi) impermeabile al sodio (corrente di K+ verso l’esterno e verso l’interno) durante il prepotenziale: aumento della permeabilità al sodio (corrente di Na+ verso l’interno), compensato dall'aumento della corrente di K+ verso l’esterno durante lo spike: aumento esponenziale della permeabilità al sodio (e della corrente di Na+), che cessa bruscamente durante la ripolarizzazione: aumento (tardivo) della permeabilità al potassio (corrente di K+ verso l’esterno).
 impermeabile al sodio (corrente di K+ verso l’esterno e verso l’interno) durante il prepotenziale: aumento della permeabilità al sodio (corrente di Na+ verso l’interno), compensato dall aumento della corrente di K+ verso l’esterno. durante lo spike: aumento esponenziale della permeabilità al sodio (e della corrente di Na+), che cessa bruscamente. durante la ripolarizzazione: aumento (tardivo) della permeabilità al potassio (corrente di K+ verso l’esterno).")
41
-60 -80 +30 mv Tempo (millisecondi) ENa EK Correnti ioniche nel potenziale d’azione Potenziale di riposo Canali Na+ chiusi; corrente di K+ ripolarizzazione Chiusura canali Na+; apertura canali K+ (rettificazione); uscita K+ per gradiente elettrico e chimico Potenziale a punta (spike) Apertura canali Na+ voltaggio-dipendenti; aumento esponenziale INa Accumulo K+ extracellulare Potenziale postumo negativo (rallentamento) Potenziale postumo positivo (iperpolarizzazione) Pompa elettrogenica Na/K accelerata La pompa Na/K ristabilisce l’omeostasi prepotenziale Aumenta INa, ma anche Ik Potenziale di riposo
; uscita K+ per gradiente elettrico e chimico. Potenziale a punta. (spike) Apertura canali Na+ voltaggio-dipendenti; aumento esponenziale INa. Accumulo K+ extracellulare. Potenziale postumo negativo (rallentamento) Potenziale postumo positivo (iperpolarizzazione) Pompa elettrogenica Na/K accelerata. La pompa Na/K ristabilisce l’omeostasi. prepotenziale. Aumenta INa, ma anche Ik. Potenziale di riposo.")
42
RUOLO DEI CANALI IONICI: si tratta di proteine intrinseche di membrana, il cui comportamento (che regola la permeabilità) dipende dal voltaggio della membrana (canali voltaggio-dipendenti). A riposo (-90 mV) i canali per il potassio sono prevalentemente aperti e quelli per il sodio prevalentemente chiusi. Esistono anche canali regolati chimicamente, attraverso recettori: ROC (receptor-operated channels)
")
43
La soglia per l’insorgenza del potenziale d’azione
-60 -80 mv ma INa Canali voltaggio dipendenti (esponenziale) IK Equilibrio di Nernst (lineare) Voltaggio soglia
 IK. Equilibrio di Nernst (lineare) Voltaggio soglia.")
44
TUTTO-O-NULLA: il potenziale d'azione per una stessa cellula, a parità di condizioni ambientali, è sempre uguale. Per questa ragione viene definito "impulso": la funzione della cellula dipende dalla presenza o assenza del potenziale d'azione, non dalla sua ampiezza e la funzione è modulata dal numero di impulsi nell'unità di tempo (frequenza di scarica).
..")
45
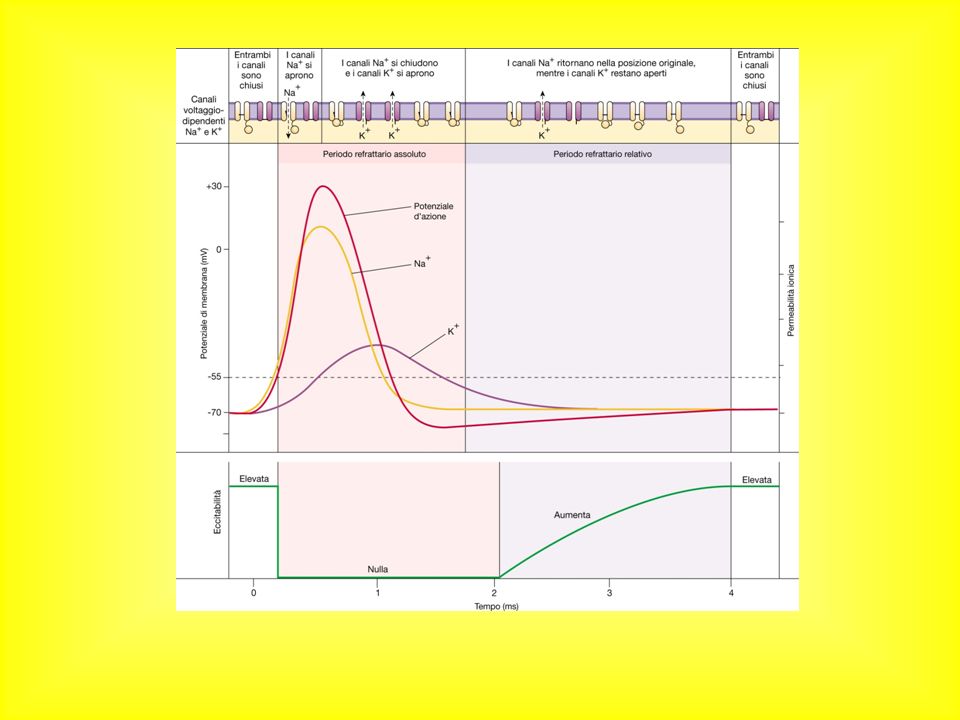
VARIAZIONI DI ECCITABILITA' DURANTE IL P.d'A.
una volta superata la soglia e fino a circa i 2/3 della ripolarizzazione la cellula non può essere nuovamente eccitata (PERIODO REFRATTARIO ASSOLUTO) In seguito può rispondere a stimoli di intensità maggiore (PERIODO REFRATTARIO RELATIVO).
 In seguito può rispondere a stimoli di intensità maggiore (PERIODO REFRATTARIO RELATIVO).")
46
Significato: In un preparato sperimentale: si aumenta l’intensità dello stimolo Nella realtà fisiologica: l’eccitazione proveniente da altre cellule (attraverso sinapsi) deve essere maggiore
 deve essere maggiore.")
47
EK -60 -80 +30 mv Tempo (millisecondi) ENa Periodo refrattario assoluto Periodo refrattario relativo Periodo supernormale Eccitabilità normale

48
ECCEZIONI ALLE REGOLE SOPRA ESPOSTE: effetto di variazioni del potenziale di riposo molto lente: accomodazione. I canali Na+ voltaggio dipendenti si inattivano prima di aprirsi; lo spike diminuisce e la cellula può diventare ineccitabile.

49
I canali Na+ voltaggio dipendenti possono trovarsi in tre diverse condizioni:
Attivi ma chiusi (a riposo, polarizzazione) Aperti (spike) Inattivi (apice dello spike; periodo refrattario assoluto)
 Aperti (spike) Inattivi (apice dello spike; periodo refrattario assoluto)")
50
Modello di funzionamento dei canali voltaggio-dipendenti
polarizzazione Fattore di attivazione E Fattore di inattivazione I depolarizzazione

51
Canale Na+ voltaggio-dipendente.
Fattore di attivazione e di inattivazione