Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Biomechanical and Molecular Regulation of Bone Remodeling
Alexander G. Robling, Alesha B. Castillo, and Charles H. Turner Annu. Rev. Biomed. Eng :455–98 (a) Sezione sottile frontale, tibia di topo, cartilagine di accrescimento: si vedono osteoclasti attivi (rosa) che riassorbono cartilagine mineralizzata e osso (nero). (b) è illustrato il processo di formazione dell’osso da un gruppo di osteoblasti (frecce bianche). L’osso è nero, i tessuti molli e le cellule sono azzurri. Fra l’osso mineralizzato e la fila di osteoblasti c’è una striscia azzurro chiaro di tessuto (osteoide) che rappresenta matrice appena formata e non ancora mineralizzata. Con l’avanzare dell’osteoide dietro agli osteoblasti, alcuni sono intrappolati (fr. verde) nell’osteoide che poi accumula minerali (fr. rossa) e isola l’osteoblasto in mezzo all’osso formato (fr. gialla); adesso è diventato un osteocita.
 Sezione sottile frontale, tibia di topo, cartilagine di accrescimento: si vedono osteoclasti attivi (rosa) che riassorbono cartilagine mineralizzata e osso (nero). (b) è illustrato il processo di formazione dell’osso da un gruppo di osteoblasti (frecce bianche). L’osso è nero, i tessuti molli e le cellule sono azzurri. Fra l’osso mineralizzato e la fila di osteoblasti c’è una striscia azzurro chiaro di tessuto (osteoide) che rappresenta matrice appena formata e non ancora mineralizzata. Con l’avanzare dell’osteoide dietro agli osteoblasti, alcuni sono intrappolati (fr. verde) nell’osteoide che poi accumula minerali (fr. rossa) e isola l’osteoblasto in mezzo all’osso formato (fr. gialla); adesso è diventato un osteocita.")
2
L’osso è un tessuto attivo metabolicamente che adatta la sua struttura agli stimoli meccanici e ripara danni strutturali attraverso il processo del rimodellamento. Le ossa della maggior parte dei mammiferi hanno quattro superfici – o strati ossei - sulle quali può verificarsi aggiunta o rimozione di osso: strato periosteo, endocorticale, trabecolare e Havesiano (intracorticale). Il riassorbimento e la formazione di osso sono effettuati dalle cellule specifiche, rispettivamente osteoclasti e osteoblasti. Quasi 40 anni fa Frost ha descritto due diversi meccanismi che stanno alla base del modellamento e del rimodellamento dell’osso, per cui tessuto osseo è selettivamente aggiunto e rimosso dalle superfici già formate in modo da ottimizzare la geometria dell’osso. Nel modellamento vi deve essere attivazione di osteoclasti e conseguente riassorbimento di osso, e di osteoblasti, con deposizione di nuovo osso, ma questo non avviene nello stesso posto; questo processo ha termine alla fine dell’accrescimento. Nel rimodellamento invece la sequenza attivazione-riassorbimento-deposizione è contigua: il rimodellamento rimuove e sostituisce pacchetti discreti e misurabili di osso. C’è spesso accoppiamento fra l’attività di osteo -blasti e -clasti, che assicura il bilancio dell’osso. In condizioni patologiche questo bilancio è perduto. Il numero di BMU (basic multicellular unit) in un determinato volume di tessuto dipende da vari fattori. Il primo è la “frequenza di nascita” di nuovi BMU o frequenza di attivazione (Ac.f); un secondo fattore dipende dalla velocità con cui un BMU attraversa l’intero strato osseo ed è chiamato fattore sigma. In media questo è di 120 giorni.
 in un determinato volume di tessuto dipende da vari fattori. Il primo è la frequenza di nascita di nuovi BMU o frequenza di attivazione (Ac.f); un secondo fattore dipende dalla velocità con cui un BMU attraversa l’intero strato osseo ed è chiamato fattore sigma. In media questo è di 120 giorni.")
3
Osteoblasti e osteoclasti si sistemano in strutture anatomiche transitorie chiamate BMU (basic multicellular units), che presentano gruppi di osteoblasti che riassorbono l’osso nel margine del caratteristico cono di taglio e sono seguiti da osteoblasti che si allineano nel cono di chiusura e depositano strati di nuovo osso in direzione centripeta per riempire la galleria scavata dagli osteoclasti. L’osso nuovo non è mineralizzato (osteoide, in blu) e incorpora progressivamente minerale (nero). La riga rossa rappresenta un capillare, che segue la formazione del BMU e fornisce nuovi osteoclasti. b) Micrografia in sezione traversa di osso lungo. Il modellamento produce lo strato esterno di osso primario, indicato dalla stella verde. Il successivo rimodellamento lascia osteoni, a forma di ciambella (frecce gialle) (ingrandito in c). Strutture rimodellate complete sono visibili sulla superficie endocorticale (frecce rosse)
 Micrografia in sezione traversa di osso lungo. Il modellamento produce lo strato esterno di osso primario, indicato dalla stella verde. Il successivo rimodellamento lascia osteoni, a forma di ciambella (frecce gialle) (ingrandito in c). Strutture rimodellate complete sono visibili sulla superficie endocorticale (frecce rosse)")
4
Il gruppo di osteoclasti che si allinea sul cono di taglio deriva da cellule staminali ematopoietiche che si trovano nel midollo e nella milza. L’osteoclastogenesi comincia quando una cellula staminale ematopoietica è stimolata a generare cellule mononucleate che diventano preosteoclasti ed entrano nel torrente circolatorio. Specifici enzimi provocano questa trasformazione. Queste cellule precursori escono dal circolo vicino al sito da riassorbire e si fondono insieme formando un osteoclasto multinucleato immaturo, per la cui formazione è necessario M-CSF (macrophage colony stimulating factor) e il “receptor activator of nuclear factor κB (RANK-L)”, un membro della famiglia dei tumor-necrosis factor. Viene contemporaneamente espressa la “tartrate-resistant acid phosphatase (Trap)”, che sarà alla base del riassorbimento osseo. La trasformazione in osteoclasto maturo avviene sotto il controllo di altri enzimi specifici, ma il RANK-L continua a giocare un ruolo essenziale. L’osteoclasto maturo si attacca alla matrice, pompa ioni H+ che solubilizzano la componente minerale e poi c’è la degradazione proteolitica della matrice organica. Lo sviluppo degli osteoblasti segue una via completamente diversa: comincia con la proliferazione locale di cellule staminali del mesenchima che risiedono nel midollo osseo in loco (e nel periosteo). Sono espressi una serie di fattori di trascrizione che indirizzano le cellule staminali verso il fenotipo osteoblastico, anziché formare adipociti, miociti o condrociti. Comincia ad essere secreto collagene tipo I e osteocalcina (OC) e un enzima fondamentale, la fosfatasi alcalina.
. Sono espressi una serie di fattori di trascrizione che indirizzano le cellule staminali verso il fenotipo osteoblastico, anziché formare adipociti, miociti o condrociti. Comincia ad essere secreto collagene tipo I e osteocalcina (OC) e un enzima fondamentale, la fosfatasi alcalina.")
5
Formazione di osteoclasti e osteoblasti
Formazione di osteoclasti e osteoblasti. (a) gli osteoclasti derivano da precursori ematopoietici nel midollo, milza, fegato. Arrivano con il sangue nel sito da riassorbire e si fondono in un policarion: cominciano ad esprimere TRAP, recettore della calcitonina e integrina beta-3. (b) Gli osteoblasti derivano da cellule staminali mesenchimali; esprimono collagene I e sialoproteina dell’osso. Gli osteoblasti sono incorporati nella matrice e diventano osteociti
 gli osteoclasti derivano da precursori ematopoietici nel midollo, milza, fegato. Arrivano con il sangue nel sito da riassorbire e si fondono in un policarion: cominciano ad esprimere TRAP, recettore della calcitonina e integrina beta-3. (b) Gli osteoblasti derivano da cellule staminali mesenchimali; esprimono collagene I e sialoproteina dell’osso. Gli osteoblasti sono incorporati nella matrice e diventano osteociti.")
6
La formazione di osteoclasti in vitro da cellule stromali richiede il contatto con altre cellule di origine mesenchimale perché queste forniscono RANK-L; si è anche trovato un fattore solubile chiamato osteoprotegerina (OPG), che è un recettore di RANK-L e ne limita la funzione sottraendolo ai recettori sull’osteoclasto. Quindi le cellule stromali possono controllare l’osteoclastogenesi aumentandola con RANK-L o inibendola con OPG. Il sistema RANK-L/OPG è il bersaglio degli ormoni che controllano l’osteogenesi (parathyroid hormone, prostaglandins, interleukins, vitamin D3, corticosteroids) Il carico meccanico ha una profonda influenza sul rimodellamento dell’osso la mancanza di carico provoca un aumento del turnover con prevalenza di riassorbimento, mentre il sovraccarico danneggia il tessuto e stimola il rimodellamento. Il turnover di tessuto osseo ha lo scopo di rimpiazzare e rimuovere il tessuto lesionato: gli osteoclasti si radunano preferibilmente nelle regioni colpite da microtraumi. Se i danni si accumulano più rapidamente di quanto avvenga la riparazione, si formano microfessure che si possono propagare fino a formare fratture. Il rimodellamento è preceduto da morte programmata degli osteociti (apoptosi). Gli effetti del carico sul rimodellamento seguono una curva a U; in un determinato intervallo di carico il rimodellamento è al minimo.
 Il carico meccanico ha una profonda influenza sul rimodellamento dell’osso la mancanza di carico provoca un aumento del turnover con prevalenza di riassorbimento, mentre il sovraccarico danneggia il tessuto e stimola il rimodellamento. Il turnover di tessuto osseo ha lo scopo di rimpiazzare e rimuovere il tessuto lesionato: gli osteoclasti si radunano preferibilmente nelle regioni colpite da microtraumi. Se i danni si accumulano più rapidamente di quanto avvenga la riparazione, si formano microfessure che si possono propagare fino a formare fratture. Il rimodellamento è preceduto da morte programmata degli osteociti (apoptosi). Gli effetti del carico sul rimodellamento seguono una curva a U; in un determinato intervallo di carico il rimodellamento è al minimo.")
7
Il turnover dell’osso (rimodellamento) aumenta in condizioni di disuso (rosso) e di uso ecessivo (arancio). Nell’intervallo fisiologico il rimodellamento è ridotto al minimo. Nel periosteo invece il rimodellamento è assente nel disuso e aumenta molto nel sovraccarico

8
(a) RANK-L prodotto da cellule dello stroma stimola l’osteoclastogenesi legandosi a recettori RANK sull’osteoclasto. Riduzione di RANK-L o inibizione da parte di OPG blocca la generazione di nuovi osteoclasti e provoca l’apoptosi di quelli esistenti. (b) Le cellule stromali controllano il riassorbimento dell’osso da parte degli osteoclasti in risposta a vari stimoli regolando l’espressione di RANK-L e OPG. La cellula stromale funziona quindi come un reostato nella regolazione dei processi di riassorbimento
 Le cellule stromali controllano il riassorbimento dell’osso da parte degli osteoclasti in risposta a vari stimoli regolando l’espressione di RANK-L e OPG. La cellula stromale funziona quindi come un reostato nella regolazione dei processi di riassorbimento.")
9
Simulazione dell’effetto dell’esercizio sull’ulna di un ratto
Simulazione dell’effetto dell’esercizio sull’ulna di un ratto. Il carico assiale provoca la curvatura laterale dell’osso: la superficie mediale è in compressione e quella laterale in tensione. È eviden-te la formazione di nuovo osso sulle superfici mediale e laterale. Carico assiale applicato per 3 min/day, 3 days/week, for 16 weeks aumenta la resistenza del 64%, con un aumento del 94% dell’energia necessaria a fratturare l’osso, mentre l’aumento di contenuto minerale è solo del 7%

10
Differenti architetture trabecolari nel collo del femore
Differenti architetture trabecolari nel collo del femore. L’angolo fra l’asse e il collo è in genere compreso fra 135° e 130° (b), ma in alcuni soggetti supera i 140° (a) o è inferiore a 110° (c). Le forze dovute al cammino provocano spinte sull’osso trabecolare del collo. Nell’anca normale le forze sono compressive (linea blu) in basso e tensili in alto (linea rossa): gli elementi trabecolari sono allineati con le direzioni principali del carico e più compatte dove esso è maggiore. In (c) le forze tensili e compressive si invertono e gli elementi trabecolari si allineano lungo queste forze. L’osso trabecolare diventa così anisotropo e aumenta la sua capacità di resistere alle forze senza aumentare la massa
, ma in alcuni soggetti supera i 140° (a) o è inferiore a 110° (c). Le forze dovute al cammino provocano spinte sull’osso trabecolare del collo. Nell’anca normale le forze sono compressive (linea blu) in basso e tensili in alto (linea rossa): gli elementi trabecolari sono allineati con le direzioni principali del carico e più compatte dove esso è maggiore. In (c) le forze tensili e compressive si invertono e gli elementi trabecolari si allineano lungo queste forze. L’osso trabecolare diventa così anisotropo e aumenta la sua capacità di resistere alle forze senza aumentare la massa.")
11
Formazione di osso periosteo in funzione del carico applicato e della frequenza della stimolazione meccanica. La soglia per iniziare la deposizione di nuovo osso si riduce aumentando la frequenza e l’accrescimento è molto più ripido in funzione del carico.

12
La massa della tibia di ratti (pallini pieni) e dell’ulna di tacchino (triangoli) aumenta dopo l’applicazione di un carico, ma l’effetto anabolico del carico si satura aumentando il numero di cicli di carico. L’effetto si esaurisce dopo 40 cicli al giorno

13
Formazione di osso sulla superficie endocorticale di tibia di ratto dopo l’applicazione in 4 stadi di 90 cicli di carico ogni secondo giorno, ad intervalli diversi: le formazione di osso aumenta aumentando la distanza fra i cicli, ma l’effetto si satura dopo 4 ore

14
Carichi in progressiva diminuzione (sinistra) o aumento (destra) per gruppi di 5 settimane. Il carico totale è uguale: in entrambi i casi c’è un aumento della resistenza al carico (ordinate), ma l’aumento è 4 volte maggiore per i carichi decrescenti. Questo dimostra che l’influenza maggiore sulla formazione di osso è esercitata dallo stimolo iniziale. In un altro esperimento un gruppo è stato caricato per 15 settimane consecutive mentre un altro ha avuto un intervallo di 5 settimane fra le prime 5 e le ultime 5. Questo gruppo, anche se è stato caricato molto meno (10 sett. anziché 15), ha accumulato più osso. Quindi una pausa nel carico sopprime gli effetti dell’adattamento cellulare e ristabilisce la meccano sensibilità
, ma l’aumento è 4 volte maggiore per i carichi decrescenti. Questo dimostra che l’influenza maggiore sulla formazione di osso è esercitata dallo stimolo iniziale. In un altro esperimento un gruppo è stato caricato per 15 settimane consecutive mentre un altro ha avuto un intervallo di 5 settimane fra le prime 5 e le ultime 5. Questo gruppo, anche se è stato caricato molto meno (10 sett. anziché 15), ha accumulato più osso. Quindi una pausa nel carico sopprime gli effetti dell’adattamento cellulare e ristabilisce la meccano sensibilità.")
15
Sezioni traverse di ossa metacarpali di cane dopo 40 settimane di disuso. Nelle ossa di cani giovani è soppressa la formazione di osso periosteo; in quelle dei cani adulti vi è un’accelerazione del riassorbimento e rimodellamento degli strati endocorticale e trabecolare (non mostrato); è anche aumentata la porosità corticale
; è anche aumentata la porosità corticale.")
16
Variazioni della mineralizzazione ossea dopo 17 settimane di bed rest: la perdita di massa ossea è soprattutto concentrata nelle estremità inferiori: La perdita non è significativa nelle braccia e nel torace, mentre c’è un aumento nel cranio. La distribuzione delle variazioni di massa ossea riflette le variazioni contemporanee della pressione del liquido interstiziale dovute alla posizione supina prolungata.

17
Le attività quotidiane espongono lo scheletro a carichi ciclici con forze molto inferiori a quelle di rottura, ma l’osso è esposto a danni irreversibili da affaticamento, che provocano microscopiche fessure. Ci sono quattro tipi di lesione: microfessure lineari, piccole rotture a reticolo (tratteggio), danno diffuso e fratture complete delle trabecole. Le lesioni aumentano con l’età soprattutto nelle femmine. Nella figura: microlesioni in una trabecola. Le frecce indicano (da sinistra) una microfessura, tratteggio e danno diffuso
 una microfessura, tratteggio e danno diffuso.")
18
Si formano microlesioni anche per sollecitazioni meccaniche fisiologiche e si accumulano con il continuare delle sollecitazioni: l’osso perde progressivamente resistenza e predispone a fratture. Questa situazione è comune in atleti che subiscono carichi ripetitivi. L’osso però ha una capacità innata di riparare il danno da fatica attraverso rimodellamento coordinato da parte di BMU: gli osteoclasti rimuovono osso danneggiato e gli osteoblasti lo sostituiscono con nuovo osso. La riparazione delle lesioni non avviene a caso ma è indirizzata ai siti di lesione Una microfessura interstiziale (due frecce) si trova nel cammino di osteoclasti sul cono di taglio di un BMU. Questo si sposta nella direzione della microfessura ad indicare che è indirizzato alla rimozione della microlesione
 si trova nel cammino di osteoclasti sul cono di taglio di un BMU. Questo si sposta nella direzione della microfessura ad indicare che è indirizzato alla rimozione della microlesione.")
19
L’osso può resistere a 4-10 milioni di cicli di carico; pertanto non basta il danno osseo per spiegare il tempo relativamente breve in cui si formano frattura da fatica. Quindi si ritiene che il processo di rimodellamento di per se sia una concausa delle fratture perché provoca un aumento transitorio della porosità, aggravando le conseguenze del danno se il sovraccarico continua. Nella figura: Osso corticale di ratto adulto: associazione spaziale fra (a) microlesione (Mdx) e osteociti in apoptosi e (b) una cavità di riassorbimento con osteociti apoptotici, che indica una relazione fra l’apoptosi provocata dal danno tessutale e l’inizio di un nuovo BMU
 microlesione (Mdx) e osteociti in apoptosi e (b) una cavità di riassorbimento con osteociti apoptotici, che indica una relazione fra l’apoptosi provocata dal danno tessutale e l’inizio di un nuovo BMU.")
20
Modello fisiologico della frattura da fatica
Modello fisiologico della frattura da fatica. Quando un osso lungo è caricato in compressione longitudinale, il carico si trasmette lungo lo strato corticale e provoca una spinta (stress) cui consegue deformazione (strain), che è inversamente proporzionale al modulo elastico (resistenza meccanica) dell’osso. In condizioni di carico fisiologico, il danno e la sua riparazione sono in equilibrio. Ma se la formazione del danno aumenta improvvisamente, per aumento improvviso della frequenza e dell’entità del carico, il danno si accumula interrompendo la normale funzione della rete degli osteociti (via a), e questi entrano in apoptosi. L’apoptosi degli osteociti provoca ulteriore rimodellamento. Il transitorio aumento della porosità che ne consegue, insieme al danno stesso, riducono la resistenza e aumentano la deformazione, aumentando il danno (via b). Se il sovraccarico continua può verificarsi un aumento rapido e incontrollato del danno e del rimodellamento, che si alimentano a vicenda fino a provocare la frattura dell’osso
 cui consegue deformazione (strain), che è inversamente proporzionale al modulo elastico (resistenza meccanica) dell’osso. In condizioni di carico fisiologico, il danno e la sua riparazione sono in equilibrio. Ma se la formazione del danno aumenta improvvisamente, per aumento improvviso della frequenza e dell’entità del carico, il danno si accumula interrompendo la normale funzione della rete degli osteociti (via a), e questi entrano in apoptosi. L’apoptosi degli osteociti provoca ulteriore rimodellamento. Il transitorio aumento della porosità che ne consegue, insieme al danno stesso, riducono la resistenza e aumentano la deformazione, aumentando il danno (via b). Se il sovraccarico continua può verificarsi un aumento rapido e incontrollato del danno e del rimodellamento, che si alimentano a vicenda fino a provocare la frattura dell’osso.")
21
Risposta generale non lineare di un sistema che simula lo sviluppo delle fratture da carico: il danno (in figura), la frequenza di attivazione, la porosità e la deformazione seguono tutti gli stessi profili (i numeri si riferiscono a cicli per giorno). Sopra un livello critico (321), con l’aggiunta di un solo ciclo (322) il modello diventa instabile. Quindi ci può essere un carico soglia al di sopra del quale le fratture da carico si sviluppano rapidamente.
, con l’aggiunta di un solo ciclo (322) il modello diventa instabile. Quindi ci può essere un carico soglia al di sopra del quale le fratture da carico si sviluppano rapidamente..")
22
Il carico meccanico stimola risposte da parte degli osteociti, che probabilmente non sono dirette ma sono dovute a variazoni del flusso di liquidi causate dal carico. Evidenziazione di mRNA: (a) dopo applicazione di carico (b) scarico
 dopo applicazione di carico (b) scarico.")
23
Il carico meccanico non influenza direttamente gli osteociti, ma provoca spostamenti di liquido extracellulare contenuto nelle lacune ed è questo movimento che stimola le cellule Modello della meccano trasduzione nell’osso. Lo scivolamento di liquido sugli osteociti (OCY) provoca un ingresso di Ca2+ attraverso canali voltaggio (V) e forse meccano (M) sensibili, aumenta anche la liberazione di ATP, che si lega a recettori purinergici (P2X, P2Y), portando alla liberazione di PGE-2, che legandosi a recettori EP stimola la formazione di osso. È coinvolto anche PTH ma non si sa come. È importante anche la via Wnt-catenina. La pressione nella cavità midollare o le forze di taglio sulle cellule stromali (MSC) stimolano NOS e liberazione di NO, che è un forte inibitore del riassorbimento osseo e probabilmente inibisce RANK-L
 provoca un ingresso di Ca2+ attraverso canali voltaggio (V) e forse meccano (M) sensibili, aumenta anche la liberazione di ATP, che si lega a recettori purinergici (P2X, P2Y), portando alla liberazione di PGE-2, che legandosi a recettori EP stimola la formazione di osso. È coinvolto anche PTH ma non si sa come. È importante anche la via Wnt-catenina. La pressione nella cavità midollare o le forze di taglio sulle cellule stromali (MSC) stimolano NOS e liberazione di NO, che è un forte inibitore del riassorbimento osseo e probabilmente inibisce RANK-L.")
24
The Effect of Exercise Training Programs on Bone Mass: A Metaanalysis of Published Controlled Trials in Pre- and Postmenopausal Women I. Wolff, J. J. van Croonenborg, H. C. G. Kemper, P. J. Kostense and J. W. R. Twisk Osteoporos Int (1999) 9:1–12 Programmi di allenamento fisico hanno la possibilità di prevenire o di invertire la perdita di osso nella colonna lombare, nel collo del femore in donne pre o post menopausa. L’effetto è modesto, ma costante e di importanza clinica, ammontando a circa 0,9% per anno di attività. Effects of Exercise Training on Bone Remodeling, Insulin-Like Growth Factors, and Bone Mineral Density in Postmenopausal Women With and Without Hormone Replacement Therapy L. A. Milliken, S. B. Going, L. B. Houtkooper, H. G. Flint-Wagner, A. Figueroa, L. L. Metcalfe, R. M. Blew, S. C. Sharp, T. G. Lohman Calcif Tissue Int (2003) 72:478–484 La terapia ormonale sostitutiva (HRT) migliora la densità ossea (BMD) e la combinazione con l’esercizio la migliora ulteriormente (non al collo del femore). L’esercizio nelle donne senza HRT cambia poco BMD. Il rimodellamento dell’osso si blocca con HRT, anche senza esercizio, mentre l’esercizio da solo non modifica il rimodellamento.
 9:1–12. Programmi di allenamento fisico hanno la possibilità di prevenire o di invertire la perdita di osso nella colonna lombare, nel collo del femore in donne pre o post menopausa. L’effetto è modesto, ma costante e di importanza clinica, ammontando a circa 0,9% per anno di attività. Effects of Exercise Training on Bone Remodeling, Insulin-Like Growth Factors, and Bone Mineral Density in Postmenopausal Women With and Without Hormone Replacement Therapy. L. A. Milliken, S. B. Going, L. B. Houtkooper, H. G. Flint-Wagner, A. Figueroa, L. L. Metcalfe, R. M. Blew, S. C. Sharp, T. G. Lohman. Calcif Tissue Int (2003) 72:478–484. La terapia ormonale sostitutiva (HRT) migliora la densità ossea (BMD) e la combinazione con l’esercizio la migliora ulteriormente (non al collo del femore). L’esercizio nelle donne senza HRT cambia poco BMD. Il rimodellamento dell’osso si blocca con HRT, anche senza esercizio, mentre l’esercizio da solo non modifica il rimodellamento.")
25
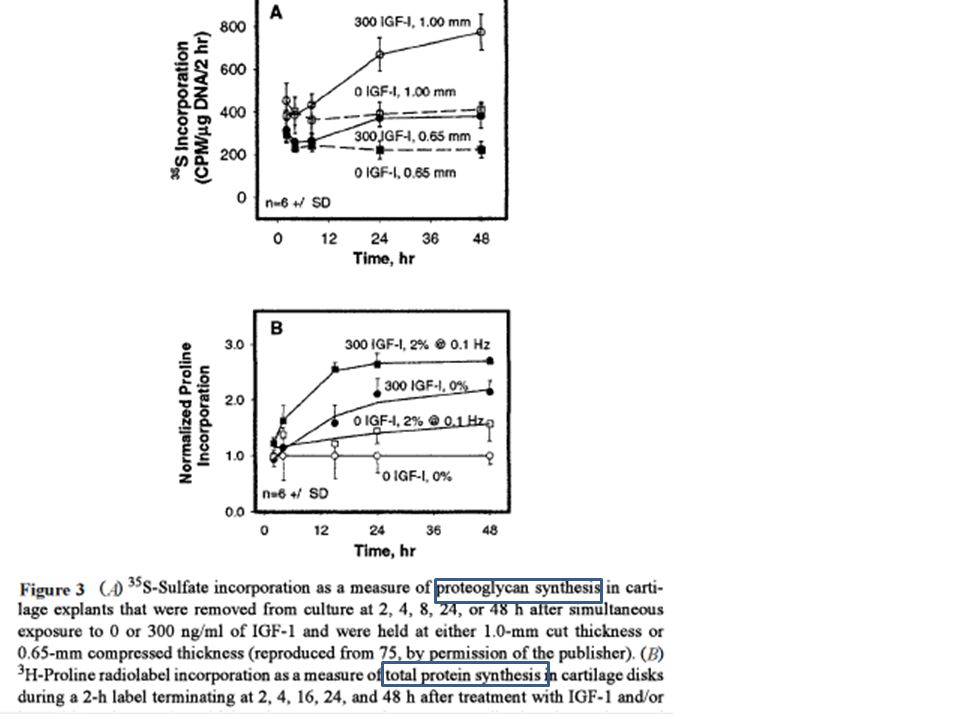
Applicazione di compressione meccanica a colture di tessuto cartilagineo: se è statica inibisce la sintesi di PG e proteine; se è dinamica stimola la produzione di matrice. Il carico applicato si trasmette a tutti gli elementi del tessuto. In un modello di dischi cartilagini di vitello neonato sottoposti ad una compressione del 3% la sintesi di PG diminuiva del 45% in 2 ore, mentre l’aggiunta di IGF-1 raddoppiava la sintesi di PG in ore. L’applicazione combinata portava ad un’iniziale inibizione della sintesi seguita da un incredibile aumento dopo 24 ore. Quindi gli effetti della compressione sono bimodali. Il ricupero dopo la rimozione della compressione è lento.

26
Rappresentazione schematica dei carichi sulla cartilagine articolare
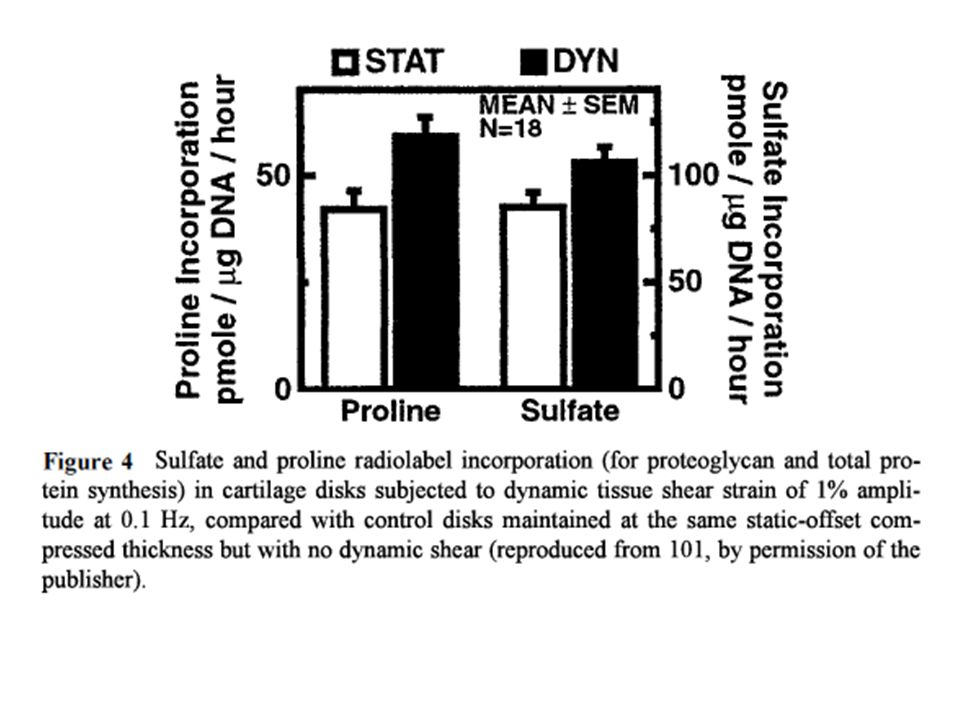
Rappresentazione schematica dei carichi sulla cartilagine articolare. La compressione dinamica della matrice extracellulare deforma le cellule e la matrice e provoca gradienti di pressione idrostatica e flusso di liquido interstiziale. La convezione di liquido e la separazione di controioni dai gruppi fissi di cariche sui proteoglicani fa sorgere correnti ioniche e differenze di potenziale. Tutte queste modificazioni non si producono con la deformazione del tessuto per scivolamento.

27
La compressione statica deforma direttamente le cellule
La compressione statica deforma direttamente le cellule. L’inibizione della biosintesi durante compressione statica comincia dopo solo un’ora e riprende due ore dopo la rimozione della compressione. In presenza di IGF-1 però la biosintesi riprende dopo l’iniziale inibizione. La compressione dinamica provoca correnti di liquido e campi elettrici che stimolano la biosintesi in maniera coerente con la direzione degli stimoli. Nell’osteoartrite le forze di compressione sulle cartilagini sono alterate, provocando perdita di proteine e in particolare di collagene e ritenzione di acqua. Le proprietà fisiche dei proteoglicani non sono compromesse, ma la capacità delle cellule di riparare le lesioni è ridotta.

30
La compressione statica stimola la produzione di mRNA ma blocca la produzione di proteine: le relazioni sono quindi complesse

31
La compressione statica altera profondamente gli organelli intracellulari e l’apparato di Golgi. Questo spiega l’alterazione della produzione proteica (collagene ed enzimi). a) basso e b) alto ingrandimento di un condrocita sotto compressione del 20%: morfologia anisotropa ordinata del reticolo endoplasmatico rugoso, mentre senza compressione la distribuzione è meno anisotropa e più disordinata. c) visualizzazione dell’apparato di Golgi con colorazione di proteine specifiche (sulfotransferasi).
 basso e b) alto ingrandimento di un condrocita sotto compressione del 20%: morfologia anisotropa ordinata del reticolo endoplasmatico rugoso, mentre senza compressione la distribuzione è meno anisotropa e più disordinata. c) visualizzazione dell’apparato di Golgi con colorazione di proteine specifiche (sulfotransferasi).")
32
Nell’osteoartrite la composizione della matrice cartilaginea è molto alterata, tanto che il tessuto si indebolisce sicché l’attrito meccanico dei movimenti articolari erode le superfici. Molte sono le famiglie di proteasi responsabili di queste aletrazioni: metalloproteinasi, proteinasi seriniche e aggrecanasi. È possibile che le vie enzimatiche siano modificate dalla compressione meccanica. Studi sperimentali su dischi di cartilagine sottoposti a compressione controllata rivelano che lo stimolo meccanico altera l’ECM, compromettendo acutamente la funzione del collagene e accelerando il degrado proteico da parte di enzimi liberati dalle cellule. Determinati livelli di compressione danno inizio all’apoptosi cellulare, indebolendo la capacità della cartilagine di rigenerarsi

Presentazioni simili


 (Ito) (IKr, IKs) (ICa-L) (INa).>")

 formazione del centro di ossificazione b) accrescimento c) modellamento e mineralizzazione Es.: ossa del cranio,>")








>")


, specializzato per la distribuzione di: gas.>")


