Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Canali al Ca++ Canali voltaggio-dipendenti Canali attivati da segnali intracellulari (proteine G, IP3, Ca++) Recettori-canale (es. recettore NMDA)
")
2
Canali al Ca++ voltaggio-dipendenti
Funzionalmente e strutturalmente simili a quelli al Na+ Ne esistono 2 classi, in base al potenziale di attivazione: Canali a bassa soglia (canali T): si attivano in seguito a piccole depolarizzazioni e si inattivano molto rapidamente importanti nel controllo dell’eccitabilità. Canali ad alta soglia: si attivano in seguito a forti depolarizzazioni e rimangono aperti più a lungo responsabili dei grandi aumenti di Ca++ intracellulare necessari per l’azione di secondo messaggero del Ca++. Ne esistono diversi sottotipi (L, N, P/Q, R), che differiscono per la sensibilità farmacologica.
: si attivano in seguito a piccole depolarizzazioni e si inattivano molto rapidamente importanti nel controllo dell’eccitabilità. Canali ad alta soglia: si attivano in seguito a forti depolarizzazioni e rimangono aperti più a lungo responsabili dei grandi aumenti di Ca++ intracellulare necessari per l’azione di secondo messaggero del Ca++. Ne esistono diversi sottotipi (L, N, P/Q, R), che differiscono per la sensibilità farmacologica.")
3
I diversi tipi di canale differiscono per la composizione in subunità.
Farmaci che ‘bloccano’ i canali ad alta soglia di tipo L sono utilizzati come antiipertensivi, antiaritmici, nell’angina (i canali di tipo L sono presenti soprattutto nel sistema cardiovascolare). Questi farmaci si legano preferenzialmente ai canali nello stato ‘inattivato’ e ‘stabilizzano’ i canali in questo stato. L’effetto è una diminuzione delle correnti di calcio transmembrana in risposta ad uno stimolo depolarizzante.
. Questi farmaci si legano preferenzialmente ai canali nello stato ‘inattivato’ e ‘stabilizzano’ i canali in questo stato. L’effetto è una diminuzione delle correnti di calcio transmembrana in risposta ad uno stimolo depolarizzante.")
4
Nel cuore, la diminuzione della corrente del calcio determina:
riduzione della forza di contrazione del miocardio; diminuzione della frequenza del pacemaker del nodo del seno; diminuzione della velocità di conduzione del nodo atrioventricolare.

5
Nella muscolatura liscia dei vasi, la diminuzione della corrente del calcio determina rilasciamento riduzione delle resistenze vascolari periferiche (arteriole) diminuzione della pressione arteriosa. Gli effetti totali sono: Diminuzione della pressione arteriosa; Diminuzione del lavoro cardiaco e quindi del fabbisogno di ossigeno da parte del cuore; Diminuzione del plateau del PA e diminuzione della velocità di conduzione AV

6
Proteolisi e proteasi: fisiologia, patologia, farmacologia
La proteolisi è il processo di degradazione idrolitica delle proteine (idrolisi del legame peptidico). E’ catalizzata da enzimi detti proteasi E’ una reazione irreversibile
. E’ catalizzata da enzimi detti proteasi. E’ una reazione irreversibile.")
7
Le proteasi Esistono 2 tipi di proteasi: Endopeptidasi: idrolizzano un legame peptidico interno Esopeptidasi: idrolizzano un legame petidico terminale (carbossipeptidasi, aminopeptidasi); alcune ‘staccano’ 2 o 3 aminoacidi terminali (di- o tri-peptidil-peptidasi)
; alcune ‘staccano’ 2 o 3 aminoacidi terminali (di- o tri-peptidil-peptidasi)")
8
Ruoli fisiologici della proteolisi
Turn-over delle proteine (catabolismo): proteine la cui presenza deve essere temporalmente controllata; proteine parzialmente alterate Formazione di proteine funzionali da precursori (es. pre-pro-albumina pro-albumina albumina) Attivazione di proteine funzionali (es. enzimi) da precursori inattivi Formazione di trasmettitori (neuropeptidi, ormoni ecc.) da precursori inattivi (‘maturazione’) Degradazione intracellulare di proteine provenienti dall’esterno (esogene o endogene) Le proteasi svolgono un ruolo nel processo di apoptosi
: proteine la cui presenza deve essere temporalmente controllata; proteine parzialmente alterate. Formazione di proteine funzionali da precursori (es. pre-pro-albumina pro-albumina albumina) Attivazione di proteine funzionali (es. enzimi) da precursori inattivi. Formazione di trasmettitori (neuropeptidi, ormoni ecc.) da precursori inattivi (‘maturazione’) Degradazione intracellulare di proteine provenienti dall’esterno (esogene o endogene) Le proteasi svolgono un ruolo nel processo di apoptosi.")
9
Regolazione del numero di recettori:
internalizzazione riciclaggio degradazione Proteasi di membrana: Degradazione di peptidi attivi (es. bradichinina) Attivazione di peptidi inattivi (es. angiotensina I) Proteasi extracellulari: Controllo della matrice extracellulare Coagulazione ecc.
 Attivazione di peptidi inattivi (es. angiotensina I) Proteasi extracellulari: Controllo della matrice extracellulare. Coagulazione ecc.")
10
Regolazione delle proteasi
L’attività delle proteasi è strettamente controllata, per evitare processi di degradazione indiscriminata: Le proteasi sono sintetizzate come precursori inattivi, che vengono attivati in condizioni particolari (es. pH acido) o in risposta a specifici segnali Segregazione: le proteasi sono in genere segregate in particolari compartimenti cellulari (es. lisosomi) Inibitori: sono presenti in eccesso rispetto alle proteasi
 o in risposta a specifici segnali. Segregazione: le proteasi sono in genere segregate in particolari compartimenti cellulari (es. lisosomi) Inibitori: sono presenti in eccesso rispetto alle proteasi.")
11
Proteasi nella patologia. 1. Ischemia e necrosi
Proteasi Ca++-dipendenti: le calpaine Sono ubiquitarie Degradano un grande numero di proteine Sono regolate dall’inibitore calpastatina Ruolo patologico: aumento eccessivo e/o prolungato di Ca++ intracellulare (cellule ischemiche) attivazione generalizzata delle calpaine proteolisi indiscriminata necrosi.
 attivazione generalizzata delle calpaine proteolisi indiscriminata necrosi.")
12
I bloccanti dei canali del Ca++ sono inefficaci nella protezione dal danno ischemico (anzi sono dannosi) Attualmente non esistono inibitori delle calpaine sufficientemente specifici da poter essere utilizzati per la terapia.

13
Proteasi e patologia. 2. Proteasi esogene
Agenti infettivi intracellulari possono: Introdurre nella cellula proteasi estranee, la cui attività non può essere controllata dalla cellula (es. virus, P. falciparium ecc.). Queste proteasi possono essere necessarie per il ciclo vitale dell’agente infettivo e/o danneggiare la cellula ospite. Introdurre inibitori delle proteasi cellulari
. Queste proteasi possono essere necessarie per il ciclo vitale dell’agente infettivo e/o danneggiare la cellula ospite. Introdurre inibitori delle proteasi cellulari.")
14
Inibitori delle proteasi nella terapia dell’AIDS
Una proteasi virale, che scinde i precursori in proteine funzionali, è necessaria alla maturazione del virus. Gli inibitori di questa proteasi (saquinavir, indinavir ecc.), utilizzati insieme ad altri farmaci, hanno rivoluzionato la terapia anti-AIDS.
, utilizzati insieme ad altri farmaci, hanno rivoluzionato la terapia anti-AIDS.")
15
Representation of HIV polyprotein with protease enzyme approaching polyprotein (top left). Polyprotein inserted into the active cleft of protease enzyme (top right). Polyprotein cut (bottom left). The HIV polyprotein with protease inhibitor occupying enzymatic cleft of protease enzyme, preventing insertion of polyprotein (bottom right).
. Polyprotein cut (bottom left). The HIV polyprotein with protease inhibitor occupying enzymatic cleft of protease enzyme, preventing insertion of polyprotein (bottom right)..")
16
Il loro sviluppo è stato il frutto di una progettazione della molecola del farmaco (drug design) basato sulla conoscenza della struttura tridimensionale del sito attivo dell’enzima e del suo funzionamento. Un problema è rappresentato dalle mutazioni virali, che portano alla formazione di proteasi meno sensibili.

17
ACE-inibitori L’ACE (Angitensin Converting Enzyme) è una esopeptidasi che ‘stacca’ il dipeptide terminale dell’angiotensina I, formando l’angiotensina II, biologicamente attivo. L’enzima idrolizza anche la bradichinina, inattivandola. La bradichinina produce, tra l’altro, anche vasodilatazione.
 è una esopeptidasi che ‘stacca’ il dipeptide terminale dell’angiotensina I, formando l’angiotensina II, biologicamente attivo. L’enzima idrolizza anche la bradichinina, inattivandola. La bradichinina produce, tra l’altro, anche vasodilatazione.")
18
Gli inibitori dell’ACE (ACE-inibitori) hanno un ruolo fondamentale nella terapia di ipertensione e insufficienza cardiaca. Sono stati sintetizzati in base alla conoscenza della struttura del sito attivo dell’enzima. Più recentemente sono stati sintetizzati degli antagonisti del recettore per l’angiotensina. I loro effetti sono sovrapponibili a quelli degli ACE-inibitori.

19
Farmacologia del sistema nervoso periferico
La sezione afferente porta al sistema nervoso centrale (SNC) le informazioni provenienti dal corpo. La sezione efferente distribuisce al corpo le informazioni provenienti dal SNC. Vi sono organi (es. fegato) o tessuti (es. tessuto adiposo) la cui funzionalità non è regolata dal sistema nervoso
 le informazioni provenienti dal corpo. La sezione efferente distribuisce al corpo le informazioni provenienti dal SNC. Vi sono organi (es. fegato) o tessuti (es. tessuto adiposo) la cui funzionalità non è regolata dal sistema nervoso.")
20
Il SNA agisce sotto il controllo del SNC, che elabora le informazioni provenienti dalla periferia.

21
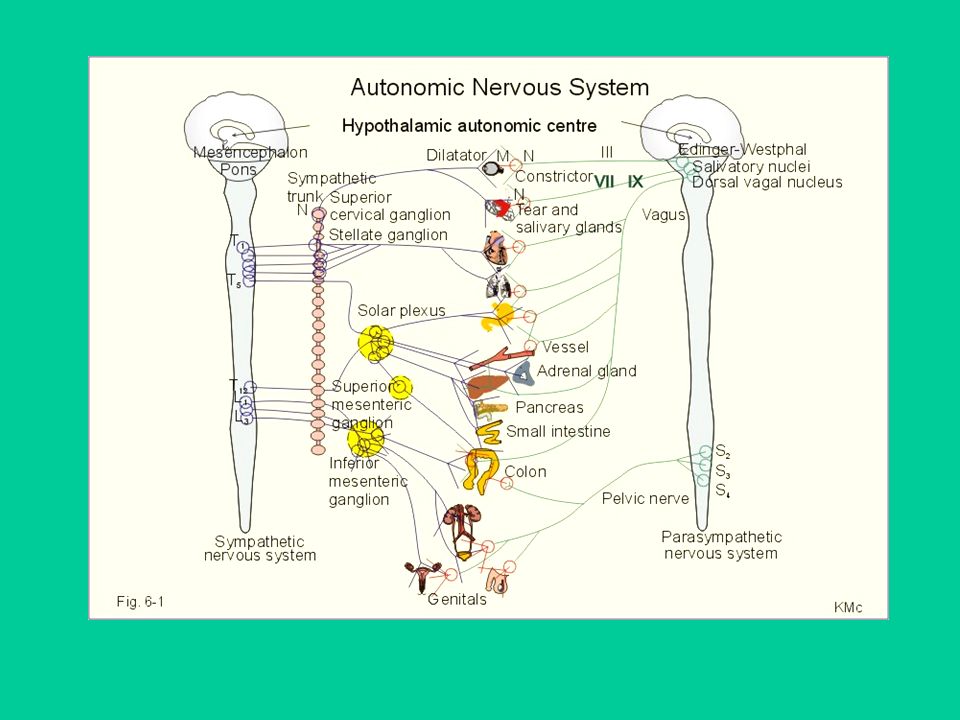
Diversità di organizzazione anatomica del sistema autonomo e del sistema somatico

22
Il sistema nervoso somatico porta le informazioni ai muscoli scheletrici.
Il sistema nervoso autonomo (SNA) distribuisce le informazioni a molti organi e tessuti:
 distribuisce le informazioni a molti organi e tessuti:")
23
Il sistema nervoso autonomo (SNA)
Regola (es. diametro pupillare) o modula (es. frequenza cardiaca) le funzioni degli organi innervati, di momento in momento. L’organizzazione anatomica delle 2 branche, simpatico e parasimpatico, è molto diversa e si riflette in una diversa regolazione delle funzioni periferiche.
 o modula (es. frequenza cardiaca) le funzioni degli organi innervati, di momento in momento. L’organizzazione anatomica delle 2 branche, simpatico e parasimpatico, è molto diversa e si riflette in una diversa regolazione delle funzioni periferiche.")
24
Il sistema nervoso enterico
E’ formato da neuroni i cui corpi cellulari sono localizzati nei plessi intestinali. Questi neuroni innervano l’intestino e sono interconnessi tra loro; vi sono inoltre neuroni afferenti sensoriali che veicolano informazioni dalla parete intestinale ai plessi, dando origine ad archi riflessi locali. Il sistema enterico regola in modo autonomo la motilità e in parte le secrezioni intestinali. Utilizza molti trasmettitori, diversi da quelli del SNA. Le funzioni intestinali sono modulate anche dal SNA: neuroni simpatici e parasimpatici innervano direttamente gli organi effettori (fibre muscolari, ghiandole ecc.) o i neuroni enterici.
 o i neuroni enterici.")
26
La stimolazione simpatica genera una risposta diffusa, quella parasimpatica una risposta localizzata. I due sistemi agiscono in modo coordinato per regolare, di momento in momento, funzioni specifiche. Alcune funzioni sono controllate solo dal simpatico (es. pressione ematica), altre solo dal parasimpatico (secrezione lacrimale).
, altre solo dal parasimpatico (secrezione lacrimale).")
27
Neuroni, neurotrasmettitori, recettori delle vie nervose autonome
Le vie autonome sono costituite da 2 neuroni: un neurone pre-gangliare e uno post-gangliare Il neurone pre-gangliare è sempre colinergico: rilascia acetilcolina (ACh) come neurotrasmettitore I recettori gangliari del neurone post-gangliare sono sempre recettori colinergici nicotinici gangliari. I recettori nicotinici sono recettori-canale, la cui apertura lascia entrare ioni Na+ potenziale d’azione post-sinaptico.
 come neurotrasmettitore. I recettori gangliari del neurone post-gangliare sono sempre recettori colinergici nicotinici gangliari. I recettori nicotinici sono recettori-canale, la cui apertura lascia entrare ioni Na+ potenziale d’azione post-sinaptico.")
28
I recettori nicotinici gangliari si differenziano da quelli muscolari per la composizione in subunità; questa differenza conferisce diversa sensibilità farmacologica: antagonisti del recettore nicotinico gangliare (es. trimetafano) non sono attivi a livello muscolare. Antagonisti del recettore nicotinico muscolare (es. tubocurarina) non sono attivi a livello gangliare. Esistono composti in grado di legarsi ad entrambi i tipi di recettore nicotinico.

29
Neuroni post-gangliari
Neuroni simpatici: rilasciano nor-adrenalina come neurotrasmettitore (neuroni adrenergici). I recettori degli organi effettori possono essere (1 e 2) o (ß1, ß2, ß3). Neuroni parasimpatici: rilasciano ACh come neurotrasmettitore (neuroni colinergici). I recettori degli organi effettori sono sempre recettori colinergici muscarinici (M1, M2, M3, M4, M5). Sia i recettori adrenergici ( e ß) che quelli muscarinici sono recettori accoppiati a proteine G
. I recettori degli organi effettori possono essere (1 e 2) o (ß1, ß2, ß3). Neuroni parasimpatici: rilasciano ACh come neurotrasmettitore (neuroni colinergici). I recettori degli organi effettori sono sempre recettori colinergici muscarinici (M1, M2, M3, M4, M5). Sia i recettori adrenergici ( e ß) che quelli muscarinici sono recettori accoppiati a proteine G.")
30
Caratteristiche della trasmissione nel SNA
Velocità di trasmissione. I recettori nicotinici gangliari sono recettori-canale; il tempo intercorrente tra la stimolazione recettoriale e l’evento cellulare evocato (potenziale d’azione) è molto breve (millisecondi). Questo tipo di sinapsi è detto sinapsi ‘veloce’. Nelle sinapsi ‘lente’ i recettori sono recettori accoppiati a proteine G. I recettori degli organi innervati dai neuroni autonomi sono tutti accoppiati a proteine G; a questo livello la trasmissione è ‘lenta’.
 è molto breve (millisecondi). Questo tipo di sinapsi è detto sinapsi ‘veloce’. Nelle sinapsi ‘lente’ i recettori sono recettori accoppiati a proteine G. I recettori degli organi innervati dai neuroni autonomi sono tutti accoppiati a proteine G; a questo livello la trasmissione è ‘lenta’.")
31
Modulazioni sinaptiche
Modulazioni sinaptiche. Il rilascio di un neurotrasmettitore ed il suo effetto sulla cellula post-sinaptica possono essere modulati dallo stesso o da altri trasmettitori o modulatori. Interazioni pre-sinaptiche: Feedback auto-inibitorio (inibizione omotropa): il neurotrasmettitore stimola recettori presinaptici (situati sullo stesso neurone da cui viene rilasciato il trasmettitore); la stimolazione del recettore pre-sinaptico determina diminuzione del rilascio del trasmettitore. L’esempio più noto è l’inibizione del rilascio della nor-adrenalina prodotta dall’interazione della nor-adrenalina stessa con i recettori 2 pre-sinaptici.
: il neurotrasmettitore stimola recettori presinaptici (situati sullo stesso neurone da cui viene rilasciato il trasmettitore); la stimolazione del recettore pre-sinaptico determina diminuzione del rilascio del trasmettitore. L’esempio più noto è l’inibizione del rilascio della nor-adrenalina prodotta dall’interazione della nor-adrenalina stessa con i recettori 2 pre-sinaptici.")
32
Interazioni eterotrope
Interazioni eterotrope. In diverse popolazioni di neuroni parasimaptici, la nor-adrenalina ( e l’adrenalina), stimolando recettori pre-sinaptici dei neuroni colinergici, inibisce il rilascio di ACh. A livello cardiaco, esiste una inibizione reciproca. Altri trasmettitori e modulatori possono inibire il rilascio di ACh o NA (es. peptidi oppioidi).
, stimolando recettori pre-sinaptici dei neuroni colinergici, inibisce il rilascio di ACh. A livello cardiaco, esiste una inibizione reciproca. Altri trasmettitori e modulatori possono inibire il rilascio di ACh o NA (es. peptidi oppioidi).")
33
Meccanismi delle interazioni pre-sinaptiche
I recettori pre-sinaptici che inibiscono il rilascio del neurotrasmettitore sono accoppiati a proteine G. La loro stimolazione causa: Fosforilazione dei canali Ca++ inibizione della loro apertura in seguito a depolarizzazione diminuito ingresso di Ca++ diminuzione del rilascio vescicolare. Apertura dei canali K+ iperpolarizzazione diminuita suscettibilità allo stimolo depolarizzante

34
Modulazione post-sinaptica
L’effetto di un trasmettitore dipende dagli effetti cellulari concomitanti di un altro trasmettitore; il ‘secondo’ trasmettitore può aumentare (sinergismo) o diminuire (antagonismo) la risposta al ‘primo’ trasmettitore. La stimolazione prolungata di un recettore può provocare la diminuzione della risposta cellulare (tolleranza), con diversi meccanismi: Diminuzione del numero dei recettori (down-regulation) Diminuzione dell’affinità del recettore per l’agonista Disaccoppiamento della stimolazione recettoriale con i meccanismi di trasduzione
 o diminuire (antagonismo) la risposta al ‘primo’ trasmettitore. La stimolazione prolungata di un recettore può provocare la diminuzione della risposta cellulare (tolleranza), con diversi meccanismi: Diminuzione del numero dei recettori (down-regulation) Diminuzione dell’affinità del recettore per l’agonista. Disaccoppiamento della stimolazione recettoriale con i meccanismi di trasduzione.")
35
Trasmissione Non-Adrenergica Non-Colinergica (NANC)
Trasmissione Non-Adrenergica Non-Colinergica (NANC). I neuroni autonomi possono utilizzare, oltre a ACh o nor-adrenalina, anche altri trasmettitori (ATP, neuropeptidi, ossido d’azoto ecc.). Spesso un neurone utilizza più di un trasmettitore o modulatore. In questi casi si parla di ‘co-trasmissione’. I co-trasmettitori possono: avere cinetiche di risposta diverse (es. ACh e neuropeptidi) modulare l’uno la risposta dell’altro (es. NPY e nor-adrenalina) essere rilasciati a diverse frequenze di stimolazione (potenziali d’azione nell’unità di tempo).
. I neuroni autonomi possono utilizzare, oltre a ACh o nor-adrenalina, anche altri trasmettitori (ATP, neuropeptidi, ossido d’azoto ecc.). Spesso un neurone utilizza più di un trasmettitore o modulatore. In questi casi si parla di ‘co-trasmissione’. I co-trasmettitori possono: avere cinetiche di risposta diverse (es. ACh e neuropeptidi) modulare l’uno la risposta dell’altro (es. NPY e nor-adrenalina) essere rilasciati a diverse frequenze di stimolazione (potenziali d’azione nell’unità di tempo).")
36
Siti d’azione di farmaci attivi sul SNA
Livello pre-sinaptico: Captazione dei precursori Accumulo del neurotrasmettitore nelle vescicole Degradazione citosolica del trasmettitore Trasmissione del potenziale d’azione Ingresso Ca++ Liberazione del trasmettitore Interazione con i recettori pre-sinaptici (autotropi ed eterotropi) modulatori del rilascio del trasmettitore
 modulatori del rilascio del trasmettitore.")
37
Interazione a livello dello spazio sinaptico
Degradazione del trasmettitore Livello post-sinaptico Interazione con i recettori post-sinaptici Interazioni con le risposte cellulari attivate dal trasmettitore Livello centrale. Modulazione dei neuroni del SNC che regolano l’attività dei neuroni autonomi (es. simpaticolitici centrali)
")
38
Farmaco-tossicologia della trasmissione colinergica
L’ACh è il trasmettitore dei neuroni pre-sinaptici simpatici e parasimpatici, dei neuroni efferenti somatici, dei neuroni colinergici centrali. Sostanze che agiscono a livello pre-sinaptico sono potenzialmente in grado di interferire con l’attività di tutti questi neuroni mancanza di selettività. Sostanze in grado di inibire il trasporto della colina nella cellula (emicolinio), il suo immagazzinamento nelle vescicole (vesamicolo), la liberazione per esocitosi (tossina botulinica) hanno interesse sperimentale e tossicologico. La tossina botulinica ha limitati usi clinici)
, il suo immagazzinamento nelle vescicole (vesamicolo), la liberazione per esocitosi (tossina botulinica) hanno interesse sperimentale e tossicologico. La tossina botulinica ha limitati usi clinici)")
39
Farmaci che interferiscono a livello post-sinaptico, legandosi ai recettori colinergici, sono più selettivi. Recettori muscarinici: sono i recettori degli organi effettori; sono anche presenti nel SNC. Gli agenti che si legano a questi recettori possono avere o meno effetti centrali in funzione del loro passaggio attraverso la barriere emato-encefalica. Esistono 5 sottotipi di recettori muscarinici. I farmaci hanno in genere scarsa selettività per i sottotipi.

40
Recettori nicotinici: sono presenti a livello centrale, gangliare e nella giunzione neuromuscolare. Tuttavia, le differenze tra i diversi sottotipi (muscolare, neuronali) determinano una diversa sensibilità farmacologica relativa selettività. Alla selettività contribuisce anche la barriera emato-encefalica.
 determinano una diversa sensibilità farmacologica relativa selettività. Alla selettività contribuisce anche la barriera emato-encefalica..")
41
Alcuni farmaci che agiscono inibendo la degradazione del trasmettitore (spazio sinaptico) sono relativamente selettivi per la giunzione neuromuscolare (nervi efferenti somatici); composti ionizzati non attraversano la barriera emato-encefalica.
 sono relativamente selettivi per la giunzione neuromuscolare (nervi efferenti somatici); composti ionizzati non attraversano la barriera emato-encefalica.")
42
Farmaci attivi sui recettori muscarinici
I 5 sottotipi di recettori muscarinici sono tutti accoppiati a proteine G. M1, M3 e M5 agiscono tramite Gq produzione InsP3 e DAG. I sottotipi M2 e M4 agiscono tramite Gi inibizione dell’adenilato ciclasi e apertura canali K+). I vari sottotipi hanno anche diverse localizzazioni. Agonisti ed antagonisti hanno scarsa selettività per un sottotipo. Gli agonisti possono quindi produrre tutti gli effetti di una stimolazione parasimpatica, gli antagonisti quello di un blocco parasimpatico generalizzato. I vari organi hanno tuttavia diversa sensibilità; gli effetti a livello dei vari organi dipendono quindi dalla concentrazione del farmaco.
. I vari sottotipi hanno anche diverse localizzazioni. Agonisti ed antagonisti hanno scarsa selettività per un sottotipo. Gli agonisti possono quindi produrre tutti gli effetti di una stimolazione parasimpatica, gli antagonisti quello di un blocco parasimpatico generalizzato. I vari organi hanno tuttavia diversa sensibilità; gli effetti a livello dei vari organi dipendono quindi dalla concentrazione del farmaco.")
Presentazioni simili
















 sono state usate nella medicina cinese Il principio attivo.>")
 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")


