Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
dell’ Ubiquitina e del Proteosoma 26S
I diversi ruoli dell’ Ubiquitina e del Proteosoma 26S nella vita e nello sviluppo delle piante

2
La regolazione dei processi di proteolisi ha un ruolo essenziale nello sviluppo di tutti gli organismi Una proteina può essere degradata perché non funziona più correttamente, oppure perché il suo folding è scorretto Altre proteine sono degradate per fornire azoto e carbonio sotto forma di aminoacidi Alcune proteine vanno incontro a proteolisi in seguito a specifici stimoli ambientali e/o di sviluppo per controllare la concentrazioni di fondamentali regolatori cellulari Nelle piante questo processo è tanto più cruciale a causa dell’alto grado di plasticità di sviluppo necessaria per la loro vita sessile Il rapido switch da uno stadio di sviluppo ad un altro richiede l’eliminazione di network regolativi preesistenti e l’assemblaggio di nuovi network, processo che spesso dipende dalla proteolisi

3
Due pathways principali:
Protein Degradation Due pathways principali: Attraverso il lisosoma (non specifico, principalmente per proteine extracellulari) Attraverso il pathway del proteasoma/ubiquitina (molto regolato e specifico)
 Attraverso il pathway del proteasoma/ubiquitina (molto regolato e specifico)")
4
Ubiquitina E’ una proteina di 76 aminoacidi altamente conservata
Originariamente isolata da Goldstein nel 1975 (da timo), poi identificata in tutti gli organismi eucarioti ed in tutti i tessuti E’ una proteina di 76 aminoacidi altamente conservata Identificata anche legata covalentemente agli istoni nel 1977 da Goldknopf & Busch Identificata come parte di un sistema proteolitico energia-dipendente da Hershko nel E’ stato dimostrato che substrati ubiquitinati sono instabili da Hershko & Varshavsky nel 1982 e anni successivi La modificazione delle proteine con l’ubiquitina è responsabile anche di processi quali l’endocitosi, la riparazione del DNA, l’attività trascrizionale
, poi identificata in tutti gli organismi eucarioti ed in tutti i tessuti. E’ una proteina di 76 aminoacidi altamente conservata. Identificata anche legata covalentemente agli istoni nel 1977 da Goldknopf & Busch. Identificata come parte di un sistema proteolitico energia-dipendente da Hershko nel E’ stato dimostrato che substrati ubiquitinati sono instabili da Hershko & Varshavsky nel 1982 e anni successivi. La modificazione delle proteine con l’ubiquitina è responsabile anche di processi quali l’endocitosi, la riparazione del DNA, l’attività trascrizionale.")
5
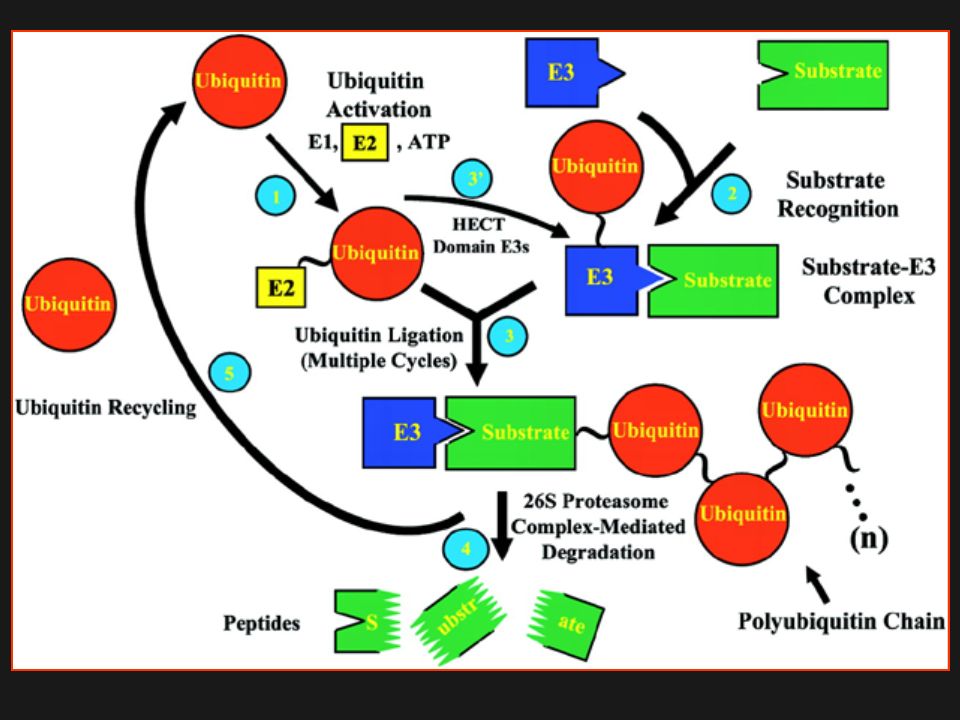
L’ubiquitinazione di una proteina target (T) comporta una serie di attività enzimatiche a cascata, che portano alla formazione di un legame isopeptidico fra il residuo G76 di Ub ed il gruppo NH2 di un residuo di L nella proteina T, formando un legame tioestere con G76. Questo processo inizia con un enzima Ub-activating (E1), responsabile del legame tioestere. Questo processo non sembra avere un ruolo regolativo e gli organismi eucariotici hanno solo poche isoforme E1. L’Ub è poi trasferita ad un enzima Ub-conjugating (E2) di nuovo mediante un legame tioestere. E2 porta l’ Ub attivata all’Ub ligasi (E3), che permette il trasferimento dell’Ub da E2 al residuo di Lys nella proteina T. E3 è generalmente considerato il più importante dei 3 enzimi nel controllo della specificità del T, perché è responsabile di recrutare il T e nel posizionarlo per il transfer di Ub da E2. Ulteriori round di coniugazione aggiungono residui di Ub al T, ed il tipo di catena di Ub formata è determinante per il destino del T. Dopo degradazione del T, i residui di Ub sono riciclati mediante azione di enzimi di de-ubiquitinazione (Dub).
 di nuovo mediante un legame tioestere. E2 porta l’ Ub attivata all’Ub ligasi (E3), che permette il trasferimento dell’Ub da E2 al residuo di Lys nella proteina T. E3 è generalmente considerato il più importante dei 3 enzimi nel controllo della specificità del T, perché è responsabile di recrutare il T e nel posizionarlo per il transfer di Ub da E2. Ulteriori round di coniugazione aggiungono residui di Ub al T, ed il tipo di catena di Ub formata è determinante per il destino del T. Dopo degradazione del T, i residui di Ub sono riciclati mediante azione di enzimi di de-ubiquitinazione (Dub).")
6
Il Pathway dell’ Ubiquitina/Proteosoma

7
Il Ciclo dell’ Ubiquitina/Proteosoma

9
La natura gerarchica del processo mediato da Ubiquitina
26S proteasome substrati

10
Alcune proteine regolative degradate dal sistema Ubiquitina/Proteosoma
Tipo Regolazione Ruolo della degradazione Esempi I positiva limite della durata Cicline (fasi G1,S,M), TFs II negativa inizio del processo Inibitori Cdk III positiva attivazione x stabilizzazione p53 o negativa tumor suppressor Dalla conferenza del Premio Nobel Avram Herscko’s
, TFs. II negativa inizio del processo Inibitori Cdk. III positiva attivazione x stabilizzazione p53. o negativa tumor suppressor. Dalla conferenza del Premio Nobel Avram Herscko’s.")
11
Perché il malfunzionamento del sistema ubiquitina-proteosoma può causare malattie?
Substrato Proteico Malattia Livello steady state ridotto state aumentato Diminuzione Degradazione Aumento Proteosoma/ Ubiquitina Substrato Proteico Funzione normale Livello steady state normale Degradazione normale mutazione

12
Degradazione delle proteine mediata da Ubiquitina
Perché studiarla nelle piante? Saccharomices Arabidopsis 5% del proteoma di Arabidopsis thaliana (1300 geni) è coinvolto nel pathway ubiquitina/ proteasoma 26S, rendendolo così uno dei meccanismi di regolazione più elaborato nelle piante.
 è coinvolto nel pathway ubiquitina/ proteasoma 26S, rendendolo così uno dei meccanismi di regolazione più elaborato nelle piante.")
13
Organizzazione e struttura di diverse E3
in associazione con l’intermedio Ub-E2 HECT Ring/U-box SCF APC HECT: singoli polipeptidi caratterizzati dal dominio HECT, contenente il dominio per il complesso Ub-E2. Proteine > 100kDa, contenenti motivi di interazione proteina-proteina nella regione N-ter, importanti per il riconoscimento del target. In lievito ed Arabidopsis ci sono solo 5 e 7 E3 di questo tipo, mentre in uomo ce ne sono >50 APC: è il complesso più sofisticato tra le E3. Contiene 12 o più subunità, compresa una Cullin e un ring finger, che formano il core dell’attività ligasica. In piante sembra esista solo un numero limitato di isoforme APC

14
Ring/U-box: il motivo Ring-E3 è formato da un ottetto di C e H che legano un atomo di Zn in un motivo H2 o HC. In Arabidopsis ci sono >400 Ring E3 potenziali. All’N-ter ci sono motivi di riconoscimento del target (WW,WD40). Il gruppo più piccolo delle E3 U-box (37 in Arabidopsis) hanno una struttura Ring finger-like, detta U-box. Il motivo ring/U-box non partecipa direttamente alla reazione con Ub ma serve da piattaforma con il Ring finger che interagisce con Ub-E2 in modo da attivare allostericamente il trasferimento di Ub alla Lys del substrato SCF (Skp, F-box, Cullin): complessi di 4 polipeptidi con attività di Ub ligasi. Una delle subunità, Rbx, contiene un ring finger di tipo H2 che lega E2-Ub. Anche SCF funziona da ponte che mette in contatto l’intermedio Ub-E2 con i target e promuove il transfer senza formare un intermedio E3-Ub. La specificità al complesso è conferita dall’F-box, che contiene diversi motivi di interazione proteina-proteina al C-ter ed un dominio F-box all’N-ter. L’F-box lega Skp. L’organizzazione combinatoriale di SCF fornisce un meccanismo per riconoscere molti substrati solo cambiando la subunità F-box. Il numero di complessi SCF nei diversi organismi eucariotici, aumenta all’aumentare della complessità dell’organismo. S.cerevisiae ne ha 14, C.elegans 337 e A.thaliana almeno 694. Molti targets dei complessi SCF devono esser fosforilati prima del riconoscimento da parte dell’ F-box.
: complessi di 4 polipeptidi con attività di Ub ligasi. Una delle subunità, Rbx, contiene un ring finger di tipo H2 che lega E2-Ub. Anche SCF funziona da ponte che mette in contatto l’intermedio Ub-E2 con i target e promuove il transfer senza formare un intermedio E3-Ub. La specificità al complesso è conferita dall’F-box, che contiene diversi motivi di interazione proteina-proteina al C-ter ed un dominio F-box all’N-ter. L’F-box lega Skp. L’organizzazione combinatoriale di SCF fornisce un meccanismo per riconoscere molti substrati solo cambiando la subunità F-box. Il numero di complessi SCF nei diversi organismi eucariotici, aumenta all’aumentare della complessità dell’organismo. S.cerevisiae ne ha 14, C.elegans 337 e A.thaliana almeno 694. Molti targets dei complessi SCF devono esser fosforilati prima del riconoscimento da parte dell’ F-box.")
15
E3 ligasi di tipo RING finger
Multi-subunità Subuntià singola Un esempio:Mdm2 Un esempio : SCF

16
La famiglia delle ubiquitin ligasi cullin-RING (CRLs)
Albero filogenetico delle CULLINs di Arabidopsis insieme a quelle di altri eucarioti. Tutte le E3 ligasi contenenti CULLINs richiedono una subunità di tipo RING-H2 che interagisce con E2. Inoltre ogni E3 porta un adattatore che recruta il substrato in modo specifico al complesso core.

17
Struttura della ubiquitina ligasi SCF

18
Ruolo dell’Ubiquitina e del Proteosoma 26S
nelle piante Molti processi vegetali sono regolati mediante ubiquitinazione e degradazione proteica Il sistema Ub/26S è attivo sia nel nucleo che nel citoplasma Molti dei mutanti di sviluppo finora studiati, presentano mutazioni in geni codificanti per F-box o per proteine con motivi Ring finger In Arabidopsis ci sono: 2 geni E1 46 geni E2 ed E2-like più di 1200 geni E3 Di questi ultimi, nel genoma di Arabidopsis sono stati identificati circa 700 geni codificanti putative F-box. Queste proteine possono essere classificate in 19 gruppi sulla base della struttura dell’ F-box

19
Il Proteosoma 26S è un complesso proteico di circa 700kDa isolato sia da nucleo che da citoplasma sia di piante che di animali In assenza di ATP, il complesso si dissocia in due subunità: Il complesso 20S proteasi core (CP) costituito da un cilindro formato da 4 anelli di 7 subunità a e b, contenente il dominio catalitico ATP-indipendente Il complesso regolativo 19S (RP), che può essere ulteriormente diviso in “lid” e “base”. E’ legato al complesso 20s ad una o ad entrambe le estremità. RP conferisce sia ATP-dipendenza che specificità di substrato Il subcomplesso “base” consiste di 6 subunità ATPasi e 3 non ATPasi Il subcomplesso “lid” consiste di 8 subunità non ATPasi DUBs: sono enzimi che deubiquitinano, rimuovendo in modo specifico le Ubs legate covalentemente ai targets. In Arabidopsis sono presenti 32 DUBs, organizzate in subfamilies. Alcune sono coinvolte nell’eliminazione di proteine danneggiate, altre nel recycling delle catene di poliUb.
 costituito da un cilindro formato da 4 anelli di 7 subunità a e b, contenente il dominio catalitico ATP-indipendente. Il complesso regolativo 19S (RP), che può essere ulteriormente diviso in lid e base . E’ legato al complesso 20s ad una o ad entrambe le estremità. RP conferisce sia ATP-dipendenza che specificità di substrato. Il subcomplesso base consiste di 6 subunità ATPasi e 3 non ATPasi. Il subcomplesso lid consiste di 8 subunità non ATPasi. DUBs: sono enzimi che deubiquitinano, rimuovendo in modo specifico le Ubs legate covalentemente ai targets. In Arabidopsis sono presenti 32 DUBs, organizzate in subfamilies. Alcune sono coinvolte nell’eliminazione di proteine danneggiate, altre nel recycling delle catene di poliUb.")
20
Il Target poliUb è riconosciuto dalla RP 19S
. Il Target poliUb è riconosciuto dalla RP 19S. La proteina è poi defoldata attraverso la particella core “base”, e poi degradata da 5 diverse proteasi all’interno della particella 20S . Il COP9 signalosome (CSN) è un complesso multiproteico altamente conservato, inizialmente identificato come repressore della luce, importante in molti processi di sviluppo delle piante. Consiste di 8 subunità e mostra notevole somiglianza con il complesso del “lid” 19S
 è un complesso multiproteico altamente conservato, inizialmente identificato come repressore della luce, importante in molti processi di sviluppo delle piante. Consiste di 8 subunità e mostra notevole somiglianza con il complesso del lid 19S.")
22
Controllo dei Pathways dei Fitormoni in Arabidopsis mediante SCF E3

23
Controllo dei Pathways dei Fitormoni in Arabidopsis mediante SCF E3
JA: è il primo ormone di cui si è dimostrato il controllo da parte di SCF-like. L’attività del proteosoma richiede COI1, che è parte del complesso SCF , i cui partner proteici non sono anocara noti. GA: sia in Arabidopsis che in riso sono presenti F-box coinvolte nel signalling delle GA, in particolare nella degradazione dei regolatori negativi della risposta alle GA, le proteine DELLA Ethylene: nel signalling dell’etilene, il TF EIN3 si lega a EBF1/2 sia in lievito (two-hybrid), che in vitro in esperimenti di pull-down assay
, che in vitro in esperimenti di pull-down assay.")
24
L’auxina induce variazioni rapide dell’espressione genica e due famiglie proteiche sono coinvolte in questa risposta: ARF (auxin response factors) e le proteine AUX/IAA. Le AUX/IAA reprimono l’espressione genica mediata da auxina inibendo l’attività delle ARFs, e un alto livello di auxina accellera la loro degradazione. TIR1 (Transport Inhibitor Response1) contiene una F-box ed è parte di un complesso SCF che interagisce con le proteine Aux/IAA
 contiene una F-box ed è parte di un complesso SCF che interagisce con le proteine Aux/IAA.")
25
Nel modello ABC di sviluppo fiorale AP3 e PI sono geni di classe B codificanti FT che funzionano come eterodimeri. La quantità di questi TF è regolata da UFO e LFY. UFO codifica per una proteina F-box che attiva/mantiene l’attività trascrizionale di PI. La possibilità che UFO formi una Ub ligasi E3 di tipo SCF è supportata dal fatto che UFO interagisce con omologhi di SKP. Inoltre è stato dimostrato che CSN interagisce con UFO in vivo. E’ probabile che il target di UFO sia un repressore di AP3.

26
skotomorfogenesi fotomorfogenesi
Tra i geni importanti per il de-eziolamento ci sono i geni COP/DET/FUS, necessari per la repressione della fotomorfogenesi al buio. COP1 interagisce con HY5, HYH e LAF TF che promuovono la foomorfogenesi. La degradazione di HY5 al buio coinvolge il proteosoma 26S e COP1. L’attività di ubiquitinazione di COP1 su HY5 e LAF1 è stata recentemente dimostrata in vitro skotomorfogenesi fotomorfogenesi

27
Variazioni sul Tema dell’Ubiquitinazione
I ruoli alternativi dell’ubiquitinazione sono stati finora dimostrati solo in lievito e nei mammiferi, ma l’alta conservazione di questi processi lascia supporre che siano presenti anche nelle piante

28
L’Ub è sintetizzata come proteina di fusione di monomeri (poliUb), o con piccole subunità ribosomali, che sono poi processate per taglio della gly C-ter. Dopo la degradazione dei targets (S:substrates), l’Ub deve essere liberata e disassemblata. Gli enzimi DUBs revertano anche l’attività di E3, rimuovendo così l’Ub dai substrati.
, l’Ub deve essere liberata e disassemblata. Gli enzimi DUBs revertano anche l’attività di E3, rimuovendo così l’Ub dai substrati..")
29
Diverse funzioni per diverse interazioni dell’Ub
Diverse funzioni per diverse interazioni dell’Ub. Schema delle diverse lys dell’Ub e delle diverse funzioni. Le funzioni delle catene legate a lys11 e lys29 sono ancora sconosciute. Le catene legate a lys48 mediano la degradazione via proteosoma, mentre le catene su lys63 hanno diverse funzioni.

Presentazioni simili













>")






