Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Immunità innata

2
Immunità innata Risposta immediata Difese attive costitutive
Ampio raggio d’azione Non si espande in maniera esponenziale Può essere sopraffatta da un’elevata carica batterica Non è dotata di memoria

3
Complementarietà dell’immunità innata e acquisita
Sempre “contro” Risposta rapida Non necessita di un primo contatto con l’Ag Può essere sopraffatta Non “on” fino a quando non entra in contatto con un Ag Risposta lenta Risposta intensa Protezione migliore in caso di secondo contatto contatto con l’Ag + - + - + + + - Non è dotata di memoria - Presentazione dell’Antigene Citochine

4
SISTEMA IMMUNITARIO Eventi cellulari e molecolari che caratterizzano le risposte umorali e cellulari MECCANISMI DI DIFESA ASPECIFICI MECCANISMI SPECIFICI PRIMA LINEA DI DIFESA SECONDA LINEA DI DIFESA TERZA LINEA DI DIFESA CUTE SALIVA SECREZIONI MUCOSE RICCHE DI PROTEINE ANTIMICROBICHE MUCO ACIDITA’ LIQUIDI LEUCOCITI FAGOCITARI PROTEINE ANTIMICROBICHE RISPOSTA INFIAMMATORIA IMMUNITA’ UMORALE CELLULARE (LINFOCITI E ANTICORPI)
")
5
Barriere fisiche e fisiologiche
Ruolo: Barriere fisiche impediscono l’ingresso dei microrganismi Barriere fisiologiche inibiscono la replicazione dei microrganismi

6
Cute Prima linea di difesa
impedisce l’ingresso di patogeni nei tessuti Le cellule della cute in superficie producono cheratina Il sebo contiene acido lattico che mantiene il pH acido (~4) Flora batterica normale (organismi commensali o simbionti) Ruolo della flora normale: Competizione con i microrganismi esogeni per sede, nutrienti, O2…(antagonismo microbico) produzione di batteriocine, antibiotici, azione di batteriofagi… Metaboliti (acido lattico abbassamento pH acido-tolleranti)
 Flora batterica normale (organismi commensali o simbionti) Ruolo della flora normale: Competizione con i microrganismi esogeni per sede, nutrienti, O2…(antagonismo microbico) produzione di batteriocine, antibiotici, azione di batteriofagi… Metaboliti (acido lattico abbassamento pH acido-tolleranti)")
7
Superfici delle Mucose Non cheratinizzano Goblet cells: secernono muco

8
pH (~2)
")
9
E’ priva di flora batterica Lacrime (lisozima)
Le regioni epiteliali di alcuni distretti non possono essere classificati né come cute né come mucose La cornea E’ priva di flora batterica Lacrime (lisozima)
")
10
Occhi: Lavaggio lacrimale lisozima Cute: Barriera anatomica Secrezioni antimicrobiche Apparato respiratorio: Muco Epielio ciliato Macrofagi alveolari Apparato gastroenterico: Acidità gastrica Flora batterica intest. Sistema genito-urinario: Lavaggio con l’urina Acidità dell’urina Lisozima Acido lattico vaginale

11
Effettori enzimatici e proteici delle mucose
Antibatterico Soprattutto su gram-positivi, perché idrolizza N-acetilglucosamina (NAG) e acido N-acetilmuramico (NAM) , portando a lisi la cellula batterica Immunostimolante E’ in grado di produrre un quadro di immunostimolazione sistemica, dopo somministrazione orale, sia in modelli sperimentali preclinici sia in trials clinici nell’uomo, indipendentemente dalla sua attività enzimatica. Lisozima o muramidasi E’ un enzima a carattere fortemente basico. Nell’uomo, è presente in diversi distretti tra cui il plasma, la saliva, il liquido lacrimale e il latte materno.
 e acido N-acetilmuramico (NAM) , portando a lisi la cellula batterica. Immunostimolante. E’ in grado di produrre un quadro di immunostimolazione sistemica, dopo somministrazione orale, sia in modelli sperimentali preclinici sia in trials clinici nell’uomo, indipendentemente dalla sua attività enzimatica. Lisozima o muramidasi E’ un enzima a carattere fortemente basico. Nell’uomo, è presente in diversi distretti tra cui il plasma, la saliva, il liquido lacrimale e il latte materno.")
12
Effettori enzimatici e proteici delle mucose
Per distruggere proteine di membrana di virus e batteri

13
Effettori enzimatici e proteici delle mucose
Peptidi anti-microbici Si inseriscono nel doppio strato fosfolipidico di membrana e distruggono la permeabilità di virus e batteri gram-negativi defensine α-defensine β-defensine

14
Effettori enzimatici e proteici delle mucose
Proteine che sequestrano il Ferro libero presente nell’organismo Lattoferrina nelle secrezioni delle mucose (saliva, lacrime, latte) e nei granuli dei granulociti neutrofili Protegge i neonati da infezioni all'apparato gastrointestinale Entrambe sottraggono il ferro ai microbi, inibendone le loro attività enzimatiche. Legando lo ione ferrico inibiscono l'attività e le capacità aggregative dei batteri gram-negativi e dei virus verso le membrane cellulari. Poiché alcuni batteri richiedono Fe per poter effettuare la replicazione cellulare, ne impedisce la proliferazione Protegge i neonati da infezioni all'apparato gastrointestinale. N.B. Alcuni batteri (Esc. Coli) possiedono chelanti del ferro che permettono al microrganismo di procurarselo anche in presenza di lattoferrina. Transferrina nel sangue
 e nei granuli dei granulociti neutrofili. Protegge i neonati da infezioni all apparato gastrointestinale. Entrambe sottraggono il ferro ai microbi, inibendone le loro attività enzimatiche. Legando lo ione ferrico inibiscono l attività e le capacità aggregative dei batteri gram-negativi e dei virus verso le membrane cellulari. Poiché alcuni batteri richiedono Fe per poter effettuare la replicazione cellulare, ne impedisce la proliferazione. Protegge i neonati da infezioni all apparato gastrointestinale. N.B. Alcuni batteri (Esc. Coli) possiedono chelanti del ferro che permettono al microrganismo di procurarselo anche in presenza di lattoferrina. Transferrina. nel sangue.")
15
Complemento E’ un complesso sistema multifattoriale, costituito da oltre 30 proteine, che ha la funzione di modulare ogni fase della risposta immunitaria. E’ un costituente importante dell’immunità innata e dell’immunità adattativa; è uno dei primi sistemi attivati in risposta ad una infezione E’ tra le forme di difesa più primitive contro le infezioni e filogeneticamente più antico Le molecole del complemento si trovano nel sangue o nei liquidi biologici, in forma inattiva (proenzimi o zimogeni) finché non sono attivati. Il meccanismo di attivazione è un meccanismo a cascata, in cui il primo elemento agisce sul successivo , rendendolo attivo e quindi capace di agire sull’elemento successivo. Il meccanismo terminale dell’azione del complemento è la formazione di un “complesso litico”, detto complesso d’attacco alla membrana (MAC), costituito dai fattori che vanno dal C5 al C9, che è in grado di eliminare i microbi. -2Il sistema del complemento consiste di circa trenta proteine, sia solubili (plasmaproteine) che di membrana, presenti in tutti i liquidi organici, che vengono indicate con la lettera C seguita da numeri progressivi. In condizioni fisiologiche queste proteine sono inattive. Circa il 90% delle proteine del complemento vengono sintetizzate nel fegato mentre il restante 10% viene sintetizzato principalmente dai monociti, macrofagi e fibroblasti. Come dimostrato dagli esperimenti di Bordet, le proteine del complemento a differenza degli anticorpi, sono proteine poco stabili e degradate dalla temperatura (termolabili). Il complemento può essere attivato direttamente dai microbi, costituendo un sistema di difesa dell’immunità innata, oppure dagli anticorpi attaccati al microbo, costituendo un braccio effettore dell’immunità adattativa. Il sistema del complemento agisce con una filosofia del tutto particolare, ovvero con un meccanismo a cascata finemente regolato, in cui un piccolo numero di proteine attivate sono in grado di produrre un gran numero di molecole effettrici. In condizioni fisiologiche, la specificità della risposta delle proteine del complemento è assicurata dal fatto che le proteine attivate si legano in modo covalente alla membrana del microorganismo da eliminare; in questo modo l’eliminazione della cellula è limitata al sito di attivazione. Inoltre, le cellule dell’organismo producono proteine in grado di inibire l’attivazione delle proteine del complemento, chiamate inibitori, in modo che l’attività proteolitica degli enzimi del complemento venga limitata e non porti al danneggiamento delle cellule dell’organismo. La mancanza di alcuni di questi inibitori porta alla formazione di patologie anche molto gravi.
 finché non sono attivati. Il meccanismo di attivazione è un meccanismo a cascata, in cui il primo elemento agisce sul successivo , rendendolo attivo e quindi capace di agire sull’elemento successivo. Il meccanismo terminale dell’azione del complemento è la formazione di un complesso litico , detto complesso d’attacco alla membrana (MAC), costituito dai fattori che vanno dal C5 al C9, che è in grado di eliminare i microbi. -2Il sistema del complemento consiste di circa trenta proteine, sia solubili (plasmaproteine) che di membrana, presenti in tutti i liquidi organici, che vengono indicate con la lettera C. seguita da numeri progressivi. In condizioni fisiologiche queste proteine sono inattive. Circa. il 90% delle proteine del complemento vengono sintetizzate nel fegato mentre il restante. 10% viene sintetizzato principalmente dai monociti, macrofagi e fibroblasti. Come. dimostrato dagli esperimenti di Bordet, le proteine del complemento a differenza degli. anticorpi, sono proteine poco stabili e degradate dalla temperatura (termolabili). Il. complemento può essere attivato direttamente dai microbi, costituendo un sistema di difesa. dell’immunità innata, oppure dagli anticorpi attaccati al microbo, costituendo un braccio. effettore dell’immunità adattativa. Il sistema del complemento agisce con una filosofia. del tutto particolare, ovvero con un meccanismo a cascata finemente regolato, in cui un. piccolo numero di proteine attivate sono in grado di produrre un gran numero di molecole. effettrici. In condizioni fisiologiche, la specificità della risposta delle proteine del complemento è. assicurata dal fatto che le proteine attivate si legano in modo covalente alla membrana del. microorganismo da eliminare; in questo modo l’eliminazione della cellula è limitata al sito di. attivazione. Inoltre, le cellule dell’organismo producono proteine in grado di inibire. l’attivazione delle proteine del complemento, chiamate inibitori, in modo che l’attività. proteolitica degli enzimi del complemento venga limitata e non porti al danneggiamento. delle cellule dell’organismo. La mancanza di alcuni di questi inibitori porta alla formazione di. patologie anche molto gravi.")
16
Complemento Funzioni : Lisi : delle membrane
cellulari , batteriche e dell’involucro lipoproteico di alcuni virus per azione del complesso litico (C5b-C9) Opsonizzazione : delle cellule capaci di fagocitosi (C3b,C4b) Risposta infiammatoria : mediante liberazione di sostanze ad azione anafilattica (vasoattiva) e chemiotattica, che favoriscono il richiamo locale di cellule fagocitanti (C3a, C5a) Rimozione degli immunocomplessi: avviene tramite il legame tra un frammento del complemento (C3b) e i recettori presenti su alcune cellule.
 Opsonizzazione : delle cellule capaci di fagocitosi (C3b,C4b) Risposta infiammatoria : mediante liberazione di sostanze ad azione anafilattica (vasoattiva) e chemiotattica, che favoriscono il richiamo locale di cellule fagocitanti (C3a, C5a) Rimozione degli immunocomplessi: avviene tramite il legame tra un frammento del complemento (C3b) e i recettori presenti su alcune cellule.")
17
Funzioni Biologiche del
Complemento B CELL ACTIVATION regolazione dell’attivazione dei linfociti B e della risposta anticorpale

18
Funzioni del Complemento
Benefiche per l’ospite: -lisi dei batteri e delle cellule infettate -opsonizzazione e fagocitosi -regolazione dell’attivazione dei linfociti B e della risposta anticorpale -clearance dei complessi immuni -clearance di cellule apoptotiche Dannose per l’ospite: -infiammazione -anafilassi

19
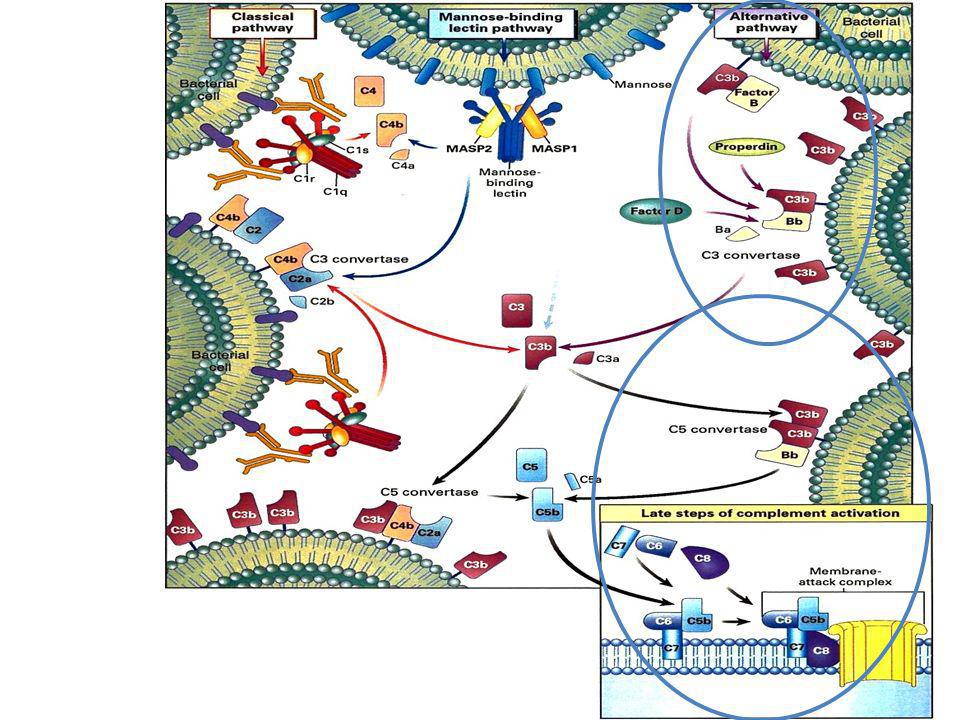
VIE DI ATTIVAZIONE DEL COMPLEMENTO

20
Attivazione dei linfociti B
VIA della LECTINA Figure 2-19 Attivazione dei linfociti B Solubilizzazione degli immunocomplessi

21
Attivatori del complemento
Superfici microbiche cariche Carboidrati batterici Sistema delle chinine Fibrina (cascata della coagulazione) Complessi Ag-Ab Prot. Fase Acuta Attivazione del Complemento Altri meccanismi di attivazione del complemento La proteina C reattiva La proteina C reattiva (CRP) è una proteina sierica prodotta dagli epatociti. Nel 1974 M.H. Kaplan e J.E. Volanakis dimostrarono che la CRP era in grado di attivare la cascata del complemento in risposta ad un componente della superficie batterica chiamato polisaccaride C o sostanza C. Si ritiene che il meccanismo con cui CRP attivi il complemento sia simile a quello della via classica, in quanto l’attivazione indotta dalla CRP richiede il componente complementare C1q. L’interazione della CRP con il C1q dipende dalla composizione biochimica della membrana bersaglio, in quanto modificazioni di essa influenzano il grado di attivazione del complemento. Il sistema della callicreina/callidina Il sistema della callicreina comprende un gruppo di proteine sieriche coinvolte nella generazione di callicreina e bradichinina, due peptidi vasoattivi importanti per la regolazione della risposta infiammatoria. Esperimenti condotti nel 1980 da numerosi gruppi di ricerca dimostrarono che la callicreina poteva fungere da attivatore del complemento, evento che si verifica mediante attivazione diretta del fattore B da parte della callicreina. Inoltre, nel 1981, R.C. Wiggins dimostrò che l’incubazione di callicreina purificata con il componente complementare C5 induceva la produzione della anafilotossina C5a, suggerendo che la callicreina poteva attivare il C5. HIV E’ probabile che la proteina gp120, una volta che si è legata sulle superfici delle cellule dell’ospite, sia in grado di legare il componente C1 o il C4 ed in tal modo dare inizio alla via classica. Altri attivatori Complemento (inattivo) Complemento (attivo)
 Complessi. Ag-Ab. Prot. Fase Acuta. Attivazione. del. Complemento. Altri meccanismi di attivazione del complemento. La proteina C reattiva. La proteina C reattiva (CRP) è una proteina sierica prodotta dagli epatociti. Nel 1974 M.H. Kaplan e J.E. Volanakis dimostrarono che la CRP era in grado di attivare la cascata del complemento in risposta ad un componente della superficie batterica chiamato polisaccaride C o sostanza C. Si ritiene che il meccanismo con cui CRP attivi il complemento sia simile a quello della via classica, in quanto l’attivazione indotta dalla CRP richiede il componente complementare C1q. L’interazione della CRP con il C1q dipende dalla composizione biochimica della membrana bersaglio, in quanto modificazioni di essa influenzano il grado di attivazione del complemento. Il sistema della callicreina/callidina. Il sistema della callicreina comprende un gruppo di proteine sieriche coinvolte nella generazione di callicreina e bradichinina, due peptidi vasoattivi importanti per la regolazione della risposta infiammatoria. Esperimenti condotti nel 1980 da numerosi gruppi di ricerca dimostrarono che la callicreina poteva fungere da attivatore del complemento, evento che si verifica mediante attivazione diretta del fattore B da parte della callicreina. Inoltre, nel 1981, R.C. Wiggins dimostrò che l’incubazione di callicreina purificata con il componente complementare C5 induceva la produzione della anafilotossina C5a, suggerendo che la callicreina poteva attivare il C5. HIV. E’ probabile che la proteina gp120, una volta che si è legata sulle superfici delle cellule dell’ospite, sia in grado di legare il componente C1 o il C4 ed in tal modo dare inizio alla via classica. Altri attivatori. Complemento. (inattivo) Complemento. (attivo)")
22
(anticorpo indipendente)
VIA DELLA LECTINA (anticorpo indipendente) MBL (Mannan Binding Lectin) e’ una collectina presente nel siero, omologa al C1q Si lega ai gruppi di mannosio presente sulle superfici batteriche Interagisce con MASP1 e MASP2 (Mannan Associated Serine Protease), omologhe a C1r e C1s, che attivano C4 e C2 portando alla formazione della C4bC2b, la C3 convertasi della Via Classica, che scinde il C3 in C3a e C3b
 MBL (Mannan Binding Lectin) e’ una collectina presente nel siero, omologa al C1q. Si lega ai gruppi di mannosio presente sulle superfici batteriche. Interagisce con MASP1 e MASP2 (Mannan Associated Serine Protease), omologhe a C1r e C1s, che attivano C4 e C2 portando alla formazione della C4bC2b, la C3 convertasi della Via Classica, che scinde il C3 in C3a e C3b.")
23
FATTORI DI INNESCO DELLA VIA ALTERNATIVA
Molti ceppi di batteri gram-positivi e gram-negativi LPS dei batteri gram- L’acido teicoico della parete dei gram+ Funghi e la parete dei lieviti (zymosan) Alcuni virus e cellule infettate da virus Alcune cellule tumorali Parassiti (tripanosomi) -
 Alcuni virus e cellule infettate da virus. Alcune cellule tumorali. Parassiti (tripanosomi) -")
26
Esistono tre diverse vie attraverso cui può essere attivato il complemento. Due di
queste, note come la via alternativa e quella lectinica, sono iniziate dai microorganismi in assenza di anticorpi. La via classica, invece, viene attivata dai complessi formati dal microorganismo ricoperto dagli anticorpi. Solo alcuni isotipi anticorpali (IgM, IgG1 e IgG3) sono in grado di attivare il complemento.
 sono in grado di attivare il complemento.")
27
Effettori cellulari dell’immunità innata

28
Effettori cellulari dell’immunità innata

29
Effettori cellulari dell’immunità innata

30
La trasduzione del segnale è necessaria per l’attivazione cellulare

31
Meccanismi di riconoscimento
Recettori endocitici : servono a stimolare l’endocitosi e la fagocitosi (es: recettore per il mannosio- CR3 del complemento) Collettine Solubili: sono pattern recognition receptors (conglutinina; CL-43: MBL; surfattanti A e D “SP-A e SP-D) sono specifiche per i carboidrati ad lato contenuto di Mannosio. Sono in grado di legare il virus dell’influenza e di inibirne la capaictà infettiva Lectine di tipo C: molecole di membrana sulle cellule DC e di Langherans servono a catturare l’Ag e di internalizzarlo. Mediano l’interazione di DC e LC con altre cellule Toll-like receptors : servono ad interagire con prodotti microbici denominati pathogen-associated molecular patterns (PAMPs)
 Collettine Solubili: sono pattern recognition receptors (conglutinina; CL-43: MBL; surfattanti A e D SP-A e SP-D) sono specifiche per i carboidrati ad lato contenuto di Mannosio. Sono in grado di legare il virus dell’influenza e di inibirne la capaictà infettiva. Lectine di tipo C: molecole di membrana sulle cellule DC e di Langherans servono a catturare l’Ag e di internalizzarlo. Mediano l’interazione di DC e LC con altre cellule. Toll-like receptors : servono ad interagire con prodotti microbici denominati pathogen-associated molecular patterns (PAMPs)")
32
Pathogen-associated molecular pattern (PAMP)
Innate immune recognition components di pareti batteriche Gram-negative bacteria Gram-positive bacteria

33
Recettore Ligando (interno/esterno) TLR 1 TLR 2 TLR 3 TLR 4 TLR 5 TLR 6 TLR 7 TLR 8 TLR 9 TLR 10 TLR 11 TLR12 TLR13 Regola il TLR 2 Lipoarabinomannano (micobatteri) Acido lipoteicoico (batteri gram +) Peptidoglicano (batteri gram -) Doppia elica RNA LPS (batteri gram -) Flagellina (numerosi batteri) Regola il TLR2 Singola elica RNA Sequenze specializzate CpG del DNA ? Toxoplasma profilin
 Acido lipoteicoico (batteri gram +) Peptidoglicano (batteri gram -) Doppia elica RNA. LPS (batteri gram -) Flagellina (numerosi batteri) Regola il TLR2. Singola elica RNA. Sequenze specializzate CpG del DNA. Toxoplasma profilin.")
35
Localizzazione cellulare dei TLRs
I TLRs che riconoscono gli acidi nucleici si trovano nel cytosol

36
Delivery of LPS to TLR4 by lipid transfer proteins
L’LPS richiede la partecipazione di altre proteine (co-recettori): CD14, CD55 e MD-2
: CD14, CD55. e MD-2.")
38
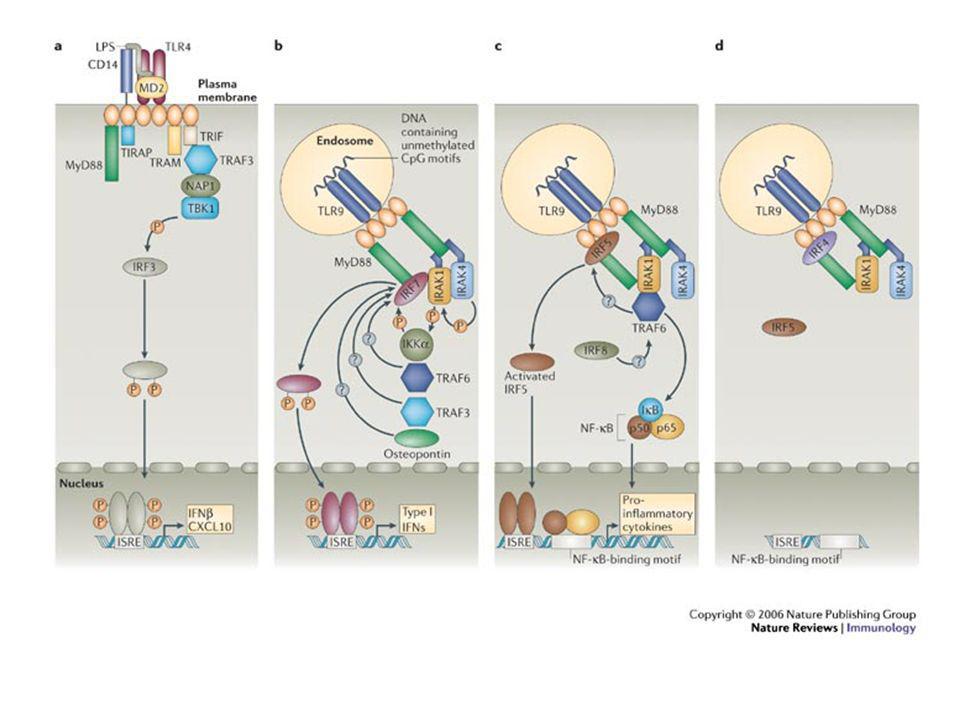
Una volta che il TLR riconosce il suo ligando trasmette segnali di attivazione

39
Il legame con il ligando induce la dimerizzazione di TLR
La via meglio caratterizzata è quella dell’NF-κB Il legame con il ligando induce la dimerizzazione di TLR Reclutamento di altre molecole del segnale Multimerizzazione dei TIR (domini di morte) presente nella porzione C-terminale di MyD88 e N-terminale di IRAK
 presente. nella porzione C-terminale. di MyD88 e N-terminale di. IRAK.")
41
Risposte innate nei confronti delle infezioni virali
Nelle infezioni virali il più importante meccanismo d’interferenza è mediata dagli INTERFERONI

42
IFN-alfa (detto anche IFN leucocitario) IFN-beta (IFN dei fibroblasti)
IFN sono una delle prime linee di difesa contro i virus in quanto indotta prima che qualsiasi altro meccanismo di difesa inizi a comparire (ad es. anticorpi, celllule T etc..) IFN-alfa (detto anche IFN leucocitario) IFN-beta (IFN dei fibroblasti) IFN-gamma(anche noto come IFN immune) L’ IFN-α e IFN-β sono anche detti IFN di Tipo I, L’ IFN-γ è detto IFN di tipo II. Ci sono 20 sottotipi di IFN-α, ma solo 1 per IFN-β e 1 per IFN-γ . IFN-κ (cheratinociti)
 IFN-alfa (detto anche IFN leucocitario) IFN-beta (IFN dei fibroblasti) IFN-gamma(anche noto come IFN immune) L’ IFN-α e IFN-β sono anche detti IFN di Tipo I, L’ IFN-γ è detto IFN di tipo II. Ci sono 20 sottotipi di IFN-α, ma solo 1 per IFN-β e 1 per IFN-γ . IFN-κ (cheratinociti)")
43
DAI: inibitore della traduzione attivato dalla doppia elica

44
Risposta Immunitaria Risposta Sistemica

Presentazioni simili








 SPECIFICA/SELETTIVA:>")



>")





 Pontieri - Elementi di Patologia Generale (ed. Piccin) Majno-Joris - Cellule,>")



