Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
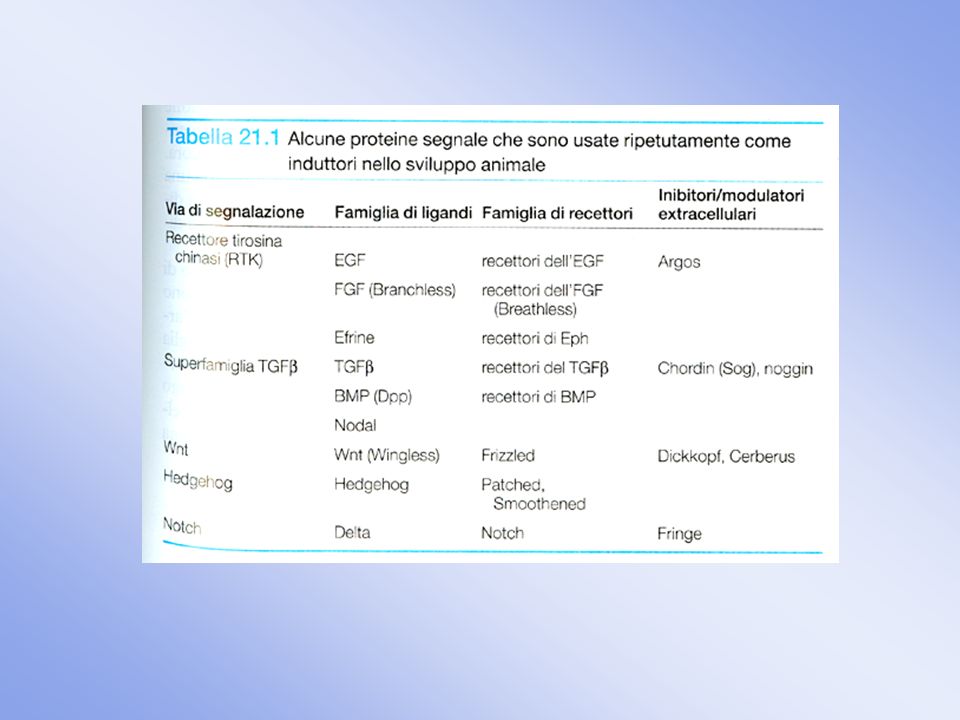
Integrazione dei pathways di segnalazione: networks
Non solo una cellula riceve piu’ segnali, ma un singolo segnale attiva uno schieramento complesso di risposte L’attivazione di un singolo recettore mobilizza piu’ vie di segnalazione Un singolo segnale puo’ generare un numero differente di segnali secondari

2
Integrazione dei pathways di segnalazione: networks
2. Le vie di segnalazione sono interconnesse da sistemi interni, cioe’ molecole che agiscono su componenti di piu’ vie, legando queste stesse vie tra loro 3. L’integrazione delle vie di segnalazione puo’ avvenire a livello della membrana plasmatica, della cascata di eventi intracellulari o nel nucleo FGFR Segnali diversi possono confluire su una stessa via Meccanismi di regolazione di Ras differenti

3
da piu’ vie di segnalazione
Integrazione dei segnali nel nucleo: l’espressione di un gene puo’ essere regolata da piu’ vie di segnalazione L’espressione del gene even skipped nei precursori muscolari di Drosophila richiede sul pro- motore l’assemblaggio di differenti fattori trascrizionali regolati da differenti vie di segnalazione

4
da piu’ vie di segnalazione
Integrazione dei segnali nel nucleo: l’espressione di un gene puo’ essere regolata da piu’ vie di segnalazione Il repressore Groucho regola lo scambio tra le vie di segnalazione mediate da EGF e Notch Gro Hasson P. and Z. Paroush 2006 British J Cancer 94:

5
Il repressore Groucho regola lo scambio tra le vie di segnalazione mediate da EGF e Notch
Overespressione di Groucho non fosforilato Overespressione di Groucho costitutivamente fosforilato

7
Diversita’ cellulare in un organismo multicellulare
In risposta a segnali esterni una cellula di un organismo multicellulare cambia la sua espressione genica

8
L’espressione genica puo’ essere regolata a livello di molti passaggi nella via DNA-> RNA -> Proteine Segnali extracellulari - > Non solo un gene ha molte proteine regolatrice che lo controllano, ma una sola proteina regolatrice puo’ controllare piu’ geni

9
Controllo combinatorio dei geni durante lo sviluppo
Combinazioni di proteine regolatrici possono generare molti tipi cellulari

10
Memoria cellulare Lo schema di espressione genica specifico di una cellula differenziata deve essere ricordato e passato alla sua progenie attraverso tutte le divisioni cellulari Meccanismi: Circuiti a feedback positivo o negativo diretto o indiretto Compattamento della cromatina regolato (Inattivazione della X) Metilazione ed Imprinting
 Metilazione ed Imprinting.")
11
Destino cellulare Come si studia?

12
Una cellula e’ determinata se e’ specializzata in modo tale da svilupparsi indipen- dentemente dal modo in cui il suo ambiente e’ disturbato Grazie alla memoria cellulare, una cellula puo’ diventare determinata molto prima di mostrare alcun segno di differenziamento Una cellula puo’ essere gia’fortemente specializzata (impegnata), ma non determinata, oppure determinata ma non ancora differenziata
, ma non determinata, oppure. determinata ma non ancora differenziata.")
13
Informazione posizionale: cellule determinate regionalmente
Cellule determinate ma non impegnate Specifica espressione genica Embrioni di pollo a 4 giorni

14
Induzione Il processo attraverso il quale una cellula o una popolazione di cellule guida un programma di espressione genica o di attivita’ proteica in altre cellule Embrione di Xenopus Induzione da parte delle cellule vegetali su quelle animali Specifico per un certo stadio Sostanza diffusibile

15
Induzione a lungo raggio

17
Sonic Hedgehog come induttore a lungo raggio nello sviluppo
dell’arto di pollo Embrione di pollo Tronco e abbozzo ali

18
I morfogeni sono induttori a lungo raggio che esercitano effetti
diversi a seconda della concentrazione

19
Strategie per la generazione di gradienti di sostanze diffusibili
A,B la semplice diffusione non spiega il gradiente C Degradazione del segnale diffusibile D Gradiente opposto di una inibitore diffu- sibile E Il segnale non e’ completamente diffu- sibile ma deve essere trasportato in ma- niera regolata

20
Induttori a corto raggio: Inibizione laterale
Cellule adiacenti inizialmente simili possono scambiare segnali a corto raggio che le spingono a diventare le une diverse dalle altre

21
Induzione sequenziale

22
Cellule sorelle possono nascere diverse per una divisione cellulare
asimmetrica

24
Caenorhabditis elegans
Piccolo nematode Vantaggi: Semplice (5autosomi e 1 cromosoma sessuale) Facile da mantenere in laboratorio Genoma sequenziato Mutanti disponibili Mappatura del destino cellulare Svantaggi: Cambiamenti evolutivi maggiori che in Drosophila Non per lo studio dell’organogenesi
 Facile da mantenere in laboratorio. Genoma sequenziato. Mutanti disponibili. Mappatura del destino cellulare. Svantaggi: Cambiamenti evolutivi maggiori che in Drosophila. Non per lo studio dell’organogenesi.")
25
Il C. elegans e’ anatomicamente semplice
trasparente 959 cellule somatiche e cellule germinali Simmetria bilaterale Due sessi: ermafrodita e maschio L’autofecondazione produce progenie omozigote

26
Sviluppo Sviluppo circa 3 giorni: embriogenesi (558 cellule), 4 stadi larvali e mute Durata vita 3 settimane

27
Caratteristica: estrema precisione nello sviluppo
Schema di interazioni cellula-cellula e sequenza di divisioni cellulari riproducibili

28
ne e la creazione della cellula fondatrice della linea germinale
Divisione asimmetrica dell’uovo per l’organizzazione anteroposteriore dell’embrio- ne e la creazione della cellula fondatrice della linea germinale La maggior parte delle cellule si differenzia in uno stadio tardivo dello sviluppo. Eccezioni: intestino (stadio 8 cells) e gonadi (stadio 16 cells) Par, 6 Geni ad effetto materno, organizzatori del citoscheletro Segregazione dei granuli P (particelle ribonucleproteiche) nella cellula fondatrice della linea germinale
 e gonadi (stadio 16 cells) Par, 6 Geni ad effetto materno, organizzatori del citoscheletro. Segregazione dei granuli P (particelle ribonucleproteiche) nella cellula fondatrice della linea germinale.")
29
Schemi progressivamente piu’ complessi vengono creati da intera-
zioni cellula-cellula Analisi genetica mutanti Notch-delta per Asse dorso-ventrale Wnt per regolare endoderma/mesoderma no Wnt= 2MS + Wnt =2E mesoderma endoderma

30
Geni eterocronici controllano i tempi di sviluppo
Meccanismi interni alla cellula, insieme a segnali passati e presenti ricevuti, dettano sia sia la sequenza dei suoi cambiamenti biochimici che i tempi delle divisioni I prodotti dei geni etero- cronici sono spesso microRNAi

31
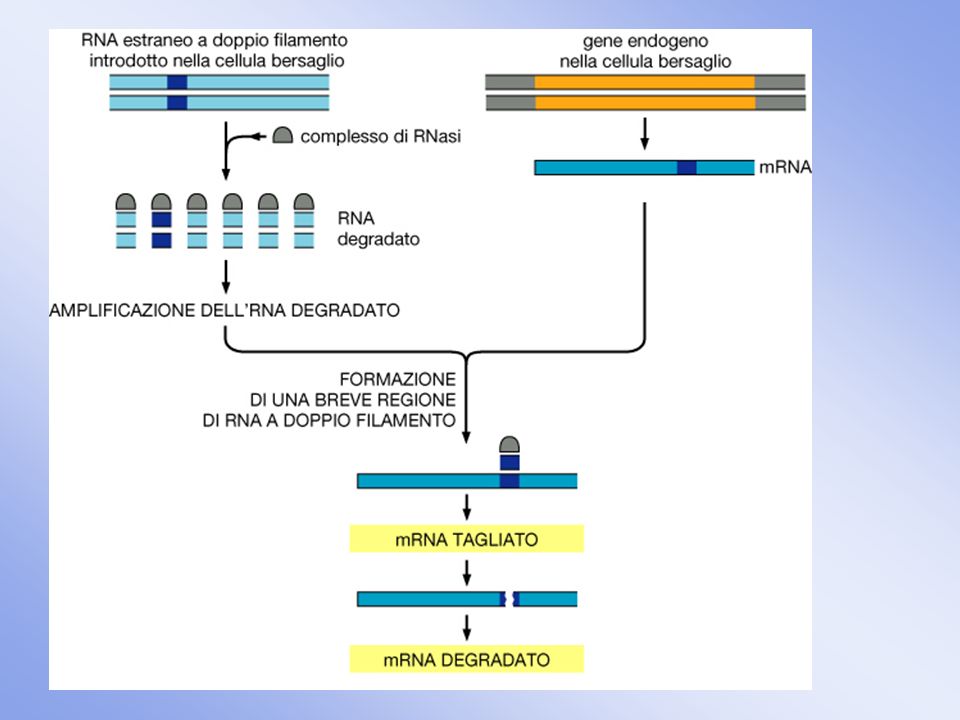
Rnai per studiare la funzione di un gene
RNA Interference Scoperto in C.elegans 3 metodi: 1) microiniezione dsRNA; 2) nel cibo batteri che producono il dsRNA di interesse; 30 immersione del verme direttamente nella soluzione con dsRNA Rnai per studiare la funzione di un gene Screening per RNAi per identificare componenti di vie di segnalazione, processi metabolici, di sviluppo etc.)
 microiniezione dsRNA; 2) nel cibo. batteri che producono il dsRNA di interesse; 30 immersione del verme direttamente nella. soluzione con dsRNA. Rnai per studiare la funzione di un gene. Screening per RNAi per identificare componenti di vie di segnalazione, processi. metabolici, di sviluppo etc.)")
33
Cellule selezionate muoiono per apoptosi come parte del programma di sviluppo
1030 nuclei sono generati, ma 131 cellule muoiono L’apoptosi dipende dall’espressione dei geni ced-3 e ced-4 in assenza di espressione di ced-9 Ced cell death abnormal Conservati tra le specie (ced-9= Bcl) Iperplasia ma non letalita’
 Iperplasia ma non letalita’")
Presentazioni simili