Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Struttura primaria Struttura secondaria (alfa-elica, filamento beta, turn) Struttura supersecondaria (motivi strutturali) Struttura terziaria All alpha All beta Alpha and beta Alpha + beta fold classe dominio Struttura quaternaria subunità

2
All- topologies

3
The lone helix There are a number of examples of small proteins (or peptides) which consist of little more than a single helix. A striking example is alamethicin, a transmembrane voltage gated ion channel, acting as a peptide antibiotic.
 which consist of little more than a single helix. A striking example is alamethicin, a transmembrane voltage gated ion channel, acting as a peptide antibiotic.")
4
The helix-turn-helix motif
The simplest packing arrangement of a domain of two helices is for them to lie antiparallel, connected by a short loop. This constitutes the structure of the small (63 residue) RNA-binding protein Rop , which is found in certain plasmids (small circular molecules of double-stranded DNA occurring in bacteria and yeast) and involved in their replication. There is a slight twist in the arrangement as shown.
 RNA-binding protein Rop , which is found in certain plasmids (small circular molecules of double-stranded DNA occurring in bacteria and yeast) and involved in their replication. There is a slight twist in the arrangement as shown.")
5
FASCIO DI QUATTRO ELICHE
Catene laterali idrofile Catene laterali idrofobe

6
Cytochrome c1 ferritin

7
cytokines

8
2 coppie di eliche parallele unite in modo antiparallelo
Fascio di 4 eliche antiparallele

9
domains which bind DNA
A three-helix bundle forms the basis of a DNA-binding domain which occurs in a number of proteins

10
globins

11
Strutture “all alpha” elica solitaria elica-turn-elica
Fascio di 4 eliche Globine (fascio di 8 eliche) Domini ad alfa elica di grandi dimensioni
 Domini ad alfa elica di grandi dimensioni.")
12
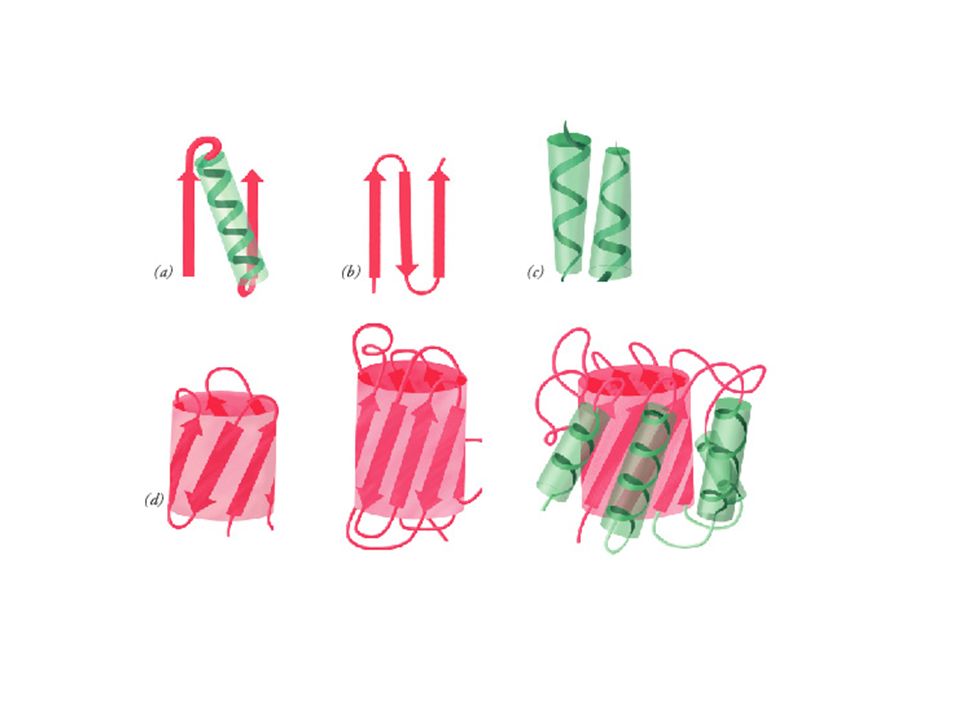
All- topologies

13
The Greek Key topology The Greek Key topology, named after a pattern that was common on Greek pottery, is shown below. Three up-and-down -strands connected by hairpins are followed by a longer connection to the fourth strand, which lies adjacent to the first.

15
Ipotetica modalità di ripiegamento di una struttura a forcina per formare la struttura a chiave greca. I filamenti 2 e 3 si ripiegano sugli altri due in modo che il filamento 2 viene ad essere allineato e antiparallelo al filamento 1.

16
Gamma-crystallin Gamma-crystallin has two domains each of which is an eight- stranded -barrel-type structure composed of two Greek keys. In fact, the structure is more accurately described as consisting of two -sheets, one consisting of strands 2,1,4,7 (white) and the other of strands 6,5,8,3 (red) as indicated in the diagram.
 and the other of strands 6,5,8,3 (red) as indicated in the diagram.")
17
Aligned and orthogonal sandwiches

18
Diagram of this -sheet arrangement in the Lipocalin family, which binds small molecules between the sheets of the sandwich.

19
barrels

20
Strutture alfa/beta

21
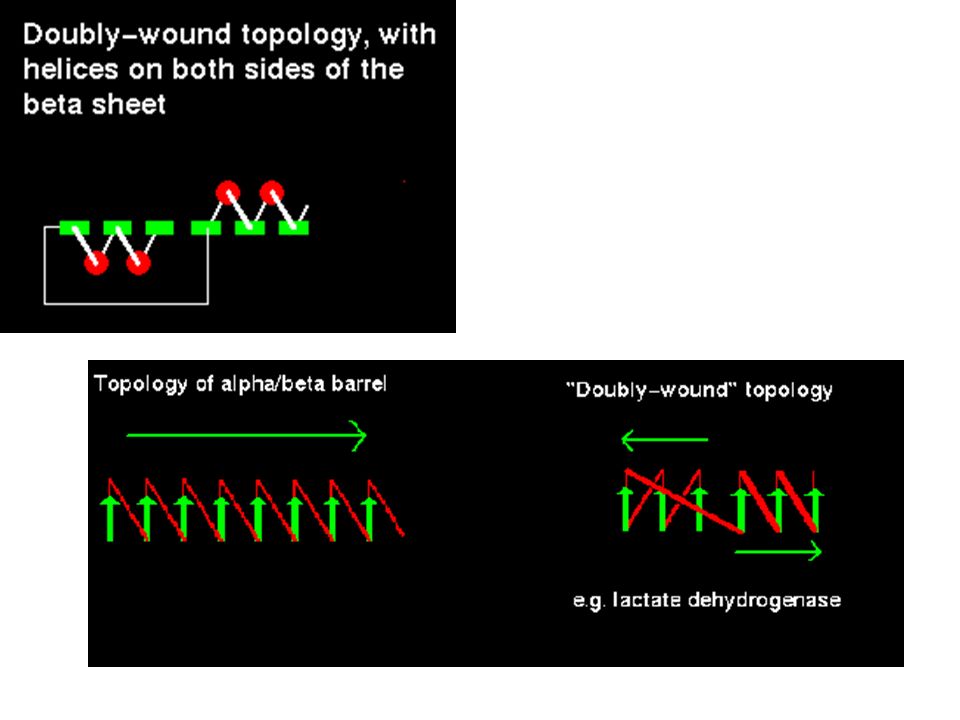
/ topologies The most regular and common domain structures consist of repeating -- supersecondary units, such that the outer layer of the structure is composed of helices packing against a central core of parallel -sheets. These folds are called / , or wound . Many enzymes, including all those involved in glycolysis , are / structures. Most / proteins are cytosolic. The -- is always right-handed. In / structures, there is a repetition of this arrangement, giving a ---.....etc sequence. The strands are parallel and hydrogen bonded to each other, while the helices are all parallel to each other, and are antiparallel to the strands. Thus the helices form a layer packing against the sheet. The ---- subunit, often present in nucleotide-binding proteins, is named the Rossman Fold, after Michael Rossman (Rao and Rossman,1973).
.")
22
Beta-alfa-beta destrorsa
Beta-alfa-beta sinistrorsa

23
Elica di collegamento Alfa-eliche sul medesimo piano del foglietto beta Alfa-eliche su piani opposti del foglietto beta

24
/ barrels Consider a sequence of eight - motifs
If the first strand hydrogen bonds to the last, then the structure closes on itself forming a barrel-like structure.

26
Enzima triosofosfato isomerasi
Enzima lattato deidrogenasi Struttura a “TIM barrel” Rossman Fold

28
In tutte le strutture / a botte il sito attivo si trova in una tasca formata dalle regioni loop che collegano le estremità carbossiliche dei filamenti beta con le adiacenti alfa eliche.

29
Nei domini con struttura / aperta ruotata il sito attivo si trova in una fessura localizzata esternamente all’estremità carbossilica del filamenti . Questa fessura è formata da due regioni loop adiacenti che collegano i due filamenti con eliche presenti su facce opposte del foglietto

30
flavodossina Adenilato chinasi Punti di inversione topologica (topological switch points)
")
31
Strutture alfa/beta I) TIM barrel
II) a)Struttura / aperta e ruotata (di tipo parallelo o misto) b) Rossmann fold III) / horseshoe (ferro di cavallo)
 a)Struttura / aperta e ruotata (di tipo parallelo o misto) - b) Rossmann fold. III) / horseshoe (ferro di cavallo)")
32
Alpha+Beta Topologies
Ribonuclease-H

33
Structural Classification of Proteins (SCOP)
Authors. Alexey G. Murzin, Loredana Lo Conte, Bartlett G. Ailey, Steven E. Brenner, Tim J. P. Hubbard, and Cyrus Chothia. Reference: Murzin A. G., Brenner S. E., Hubbard T., Chothia C. (1995). SCOP: a structural classification of proteins database for the investigation of sequences and structures. J. Mol. Biol. 247, Classes: All alpha proteins (151) All beta proteins (111) Alpha and beta proteins (a/b) (117) Mainly parallel beta sheets (beta-alpha-beta units) Alpha and beta proteins (a+b) (212) Mainly antiparallel beta sheets (segregated alpha and beta regions) Multi-domain proteins (alpha and beta) (39) Folds consisting of two or more domains belonging to different classes Membrane and cell surface proteins and peptides (12) Does not include proteins in the immune system Small proteins (59) Usually dominated by metal ligand, heme, and/or disulfide bridges Coiled coil proteins (5) Not a true class Low resolution protein structures (17) Not a true class Peptides (95) Peptides and fragments. Not a true class Designed proteins (36) Experimental structures of proteins with essentially non-natural sequences. Not a true class
. SCOP: a structural classification of proteins database for the investigation of sequences and structures. J. Mol. Biol. 247, Classes: All alpha proteins (151) All beta proteins (111) Alpha and beta proteins (a/b) (117) Mainly parallel beta sheets (beta-alpha-beta units) Alpha and beta proteins (a+b) (212) Mainly antiparallel beta sheets (segregated alpha and beta regions) Multi-domain proteins (alpha and beta) (39) Folds consisting of two or more domains belonging to different classes. Membrane and cell surface proteins and peptides (12) Does not include proteins in the immune system. Small proteins (59) Usually dominated by metal ligand, heme, and/or disulfide bridges. Coiled coil proteins (5) Not a true class. Low resolution protein structures (17) Not a true class. Peptides (95) Peptides and fragments. Not a true class. Designed proteins (36) Experimental structures of proteins with essentially non-natural sequences. Not a true class.")
35
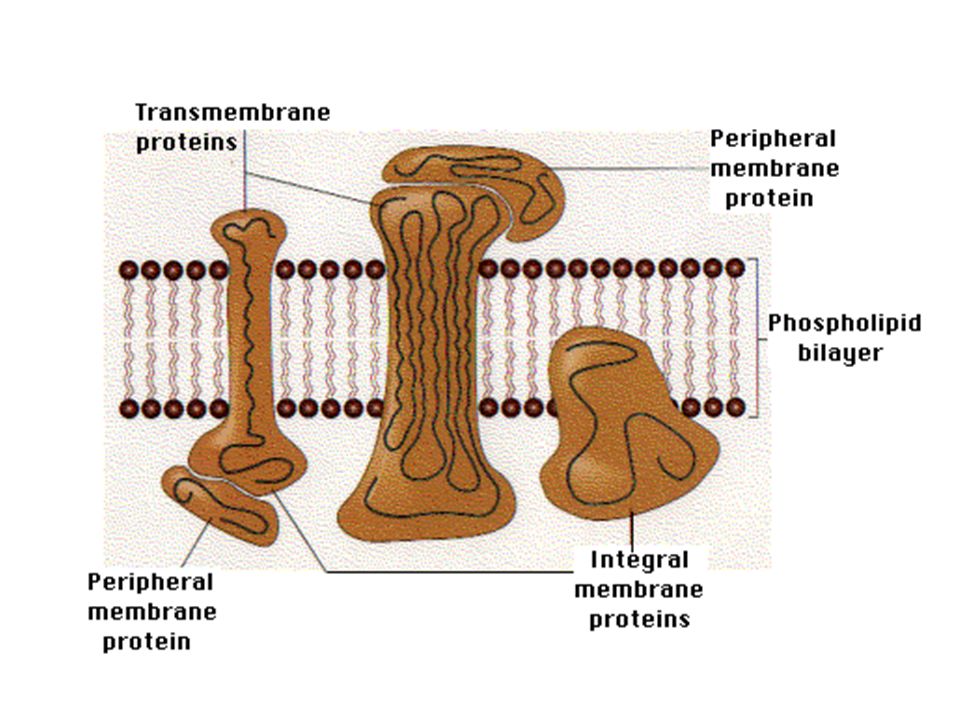
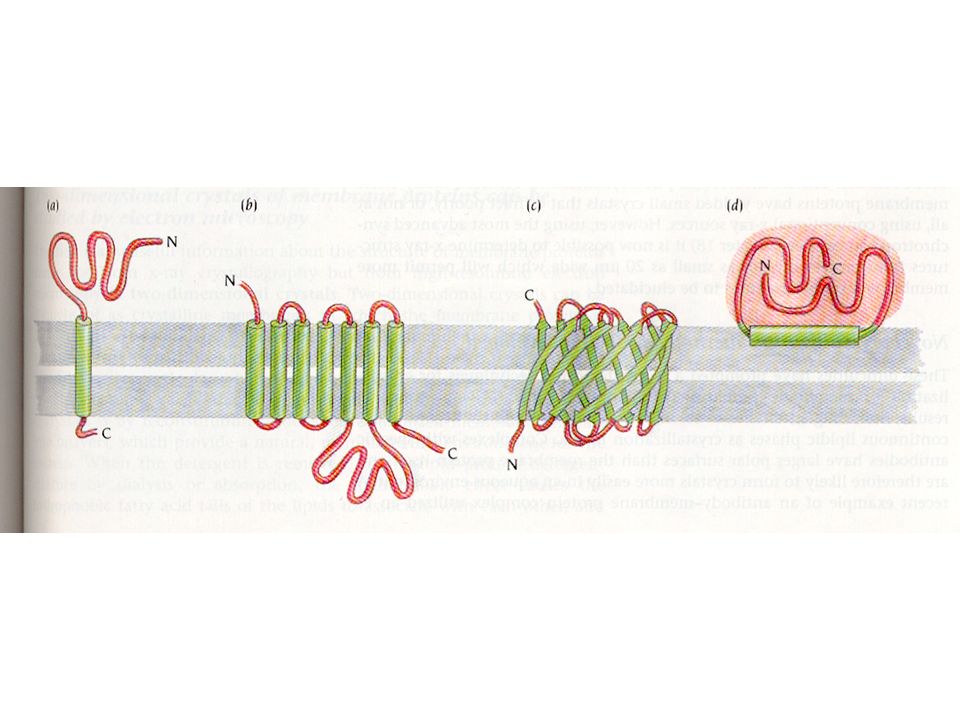
Membrane protein topology
Membrane proteins come in numerous types with a few different suggested classifications. One of the most commonly used to date is the classification method suggested by JS Singer: Type I proteins have a single TM stretch of hydrophobic residues, with the portion of the polypeptide on the NH2-terminal side of the TM domain exposed on the exterior side of the membrane and the COOH-terminal portion exposed on the cytoplasmic side. The proteins are subdivided into types Ia (cleavable signal sequences) and Ib (without cleavable signal sequence). Most eukaryotic mebrane proteins with single spanning regions are of Type Ia. Type II membrane proteins are similar to the type I class in that they span the membrane only once, but they have their amino terminus on the cytoplasmic side of the cell and the carboxy terminus on the exterior. Type III membrane proteins have multiple transmembrane domains in a single polypeptide chain (See also figure). They are also sub divided into a and b: Type IIIa molecules have cleavable signal sequences while type IIIb have their amino termini exposed on the exterior surface of the membrane, but do not have a cleavable signal sequences. Type IIIa proteins include the M and L peptides of the photoreaction center. Type IIIb proteins include e.g. cytochrome P450, and leader peptidase of E. coli.
 and Ib (without cleavable signal sequence). Most eukaryotic mebrane proteins with single spanning regions are of Type Ia. Type II membrane proteins are similar to the type I class in that they span the membrane only once, but they have their amino terminus on the cytoplasmic side of the cell and the carboxy terminus on the exterior. Type III membrane proteins have multiple transmembrane domains in a single polypeptide chain (See also figure). They are also sub divided into a and b: Type IIIa molecules have cleavable signal sequences while type IIIb have their amino termini exposed on the exterior surface of the membrane, but do not have a cleavable signal sequences. Type IIIa proteins include the M and L peptides of the photoreaction center. Type IIIb proteins include e.g. cytochrome P450, and leader peptidase of E. coli.")
37
Paradosso di Levinthal
• Proteina di 100 residui • Due gradi di liberta’ torsionali/residuo (phi,psi) • 3 conformazioni accessibili per ogni grado di liberta’ torsionale • 32x100 possibili conformazioni conformazioni esplorate/sec • Tempo richiesto per esplorare tutte le conformazioni: t = 20 x 109 anni ! • Le proteine si devono ripiegare seguendo un cammino definito, caratterizzato da conformazioni via via piu’ stabili (diminuizione di G)
 • 3 conformazioni accessibili per ogni grado di. liberta’ torsionale. • 32x100 possibili conformazioni conformazioni esplorate/sec. • Tempo richiesto per esplorare tutte le. conformazioni: t = 20 x 109 anni ! • Le proteine si devono ripiegare seguendo un. cammino definito, caratterizzato da conformazioni. via via piu’ stabili (diminuizione di G)")
Presentazioni simili


















>")
>")
