Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Modificazione & Restrizione del DNA
Si tratta di un meccanismo di protezione dei batteri contro l’invasione da parte di DNA estraneo, dovuto all’azione specifica di particolari enzimi che tagliano il DNA riconosciuto come estraneo (enzimi di restrizione) osservato inizialmente in due ceppi di E. coli K-12 e E. coli C che si comportavano in modo differente rispetto all’infezione con il fago Lambda i fagi isolati dal ceppo C crescevano bene su ceppi C ma pochissimo su ceppi K-12 I pochissimi fagi sopravvissuti alla prima infezione di fagi K-12 in seguito crescevano perfettamente su K12 e C..
 osservato inizialmente in due ceppi di E. coli K-12 e E. coli C che si comportavano in modo differente rispetto all’infezione con il fago Lambda. i fagi isolati dal ceppo C crescevano bene su ceppi C ma pochissimo su ceppi K-12. I pochissimi fagi sopravvissuti alla prima infezione di fagi K-12 in seguito crescevano perfettamente su K12 e C..")
2
POCHISSIMI lambda sopravvivono
Modificazione/Restrizione in E. coli λ CEPPO C Crescita vigorosa λ POCHISSIMI lambda sopravvivono λ CEPPO K12 CEPPO C λ I POCHISSIMI lambda sopravvissuti infettano K12 con grande efficienza Crescita vigorosa λ CEPPO K12 Crescita vigorosa

3
Interpretazione Cosa era successo ai fagi sopravvissuti alla prima infezione di E. coli K12? E. coli K-12 ha un enzima di restrizione specifico (si chiama Eco K) che degrada il DNA estraneo E.coli C NON possiede questo enzima di restrizione Il DNA dei fagi cresciuti su ceppi K-12 “sfugge” molto raramente alla degradazione perché il sito sul DNA, riconosciuto da Eco K, viene metilato ancor prima di essere tagliato In effetti ogni enzima di restrizione è una coppia di enzimi, una endonucleasi che TAGLIA il DNA e una metilasi che METILA il DNA. La metilazione IMPEDISCE il taglio del DNA, quindi il DNA “sopravvissuto” dà origine a fagi lambda che in una successiva infezione del ceppo di E. coli K12 sono “resistenti” al taglio perché hanno il sito Eco K metilato.
 che degrada il DNA estraneo. E.coli C NON possiede questo enzima di restrizione. Il DNA dei fagi cresciuti su ceppi K-12 sfugge molto raramente alla degradazione perché il sito sul DNA, riconosciuto da Eco K, viene metilato ancor prima di essere tagliato. In effetti ogni enzima di restrizione è una coppia di enzimi, una endonucleasi che TAGLIA il DNA e una metilasi che METILA il DNA. La metilazione IMPEDISCE il taglio del DNA, quindi il DNA sopravvissuto dà origine a fagi lambda che in una successiva infezione del ceppo di E. coli K12 sono resistenti al taglio perché hanno il sito Eco K metilato.")
4
Sistemi Restrizione/Modificazione: Tre tipi principali
Tipo I Tipo II Tipo III ma anche Tipo IIS Tipo IIG Tipo IV Per le tecnologie del DNA ricombinante di gran lunga i più importanti sono il tipo II

5
Sistemi R/M di tipo II • Tipo II: sono gli enzimi di restrizione per antonomasia, quelli usati per le tecnologie del DNA ricombinante. Sono ubiquitari nei procarioti – Sono sempre una coppia di enzimi DISTINTI che funzionano INDIPENDENTEMENTE • Endonucleasi (l’enzima di restrizione che taglia il sito riconosciuto): generalmente un omodimero con una sequenza di riconoscimento di 4-8 bp, di norma - ma non in tutti i casi - palindromica (G/AATTC = EcoRI) • Metiltransferasi (enzima che modifica il sito riconosciuto): monomero che riconosce la stessa sequenza della endonucleasi, metilandola (metiltransferasi di EcoRI = GAA*TTC)
: generalmente un omodimero con una sequenza di riconoscimento di 4-8 bp, di norma - ma non in tutti i casi - palindromica (G/AATTC = EcoRI) • Metiltransferasi (enzima che modifica il sito riconosciuto): monomero che riconosce la stessa sequenza della endonucleasi, metilandola (metiltransferasi di EcoRI = GAA*TTC)")
6
RE (Endonucleasi di restrizione) Tipo II
• Il DNA bersaglio può essere – non-metilato: substrato sia per la MT che per la RE; isolato da un ospite modificazione - – emi-metilato: substrato per la MT ma non per la RE; presente immediatamente dopo la replicazione del DNA in un ospite modificazione + – totalmente metilato: non è substrato né per la MT né per la RE; presente tardivamente dopo la replicazione del DNA in un ospite modificazione +

7
RE Tipo II • Meccanismo enzimatico della endonucleasi
– idrolizza lo scheletro dei fosfati- desossiribosi sulla catena del DNA – l’idrolisi avviene sul “lato” 5’ del gruppo fosfato dando origine a estremità 5’PO4 e 3’-OH – le estremità sono saldabili ad opera della(e) DNA ligasi
 DNA ligasi.")
8
RE Tipo II • Meccanismo enzimatico della metiltransferasi
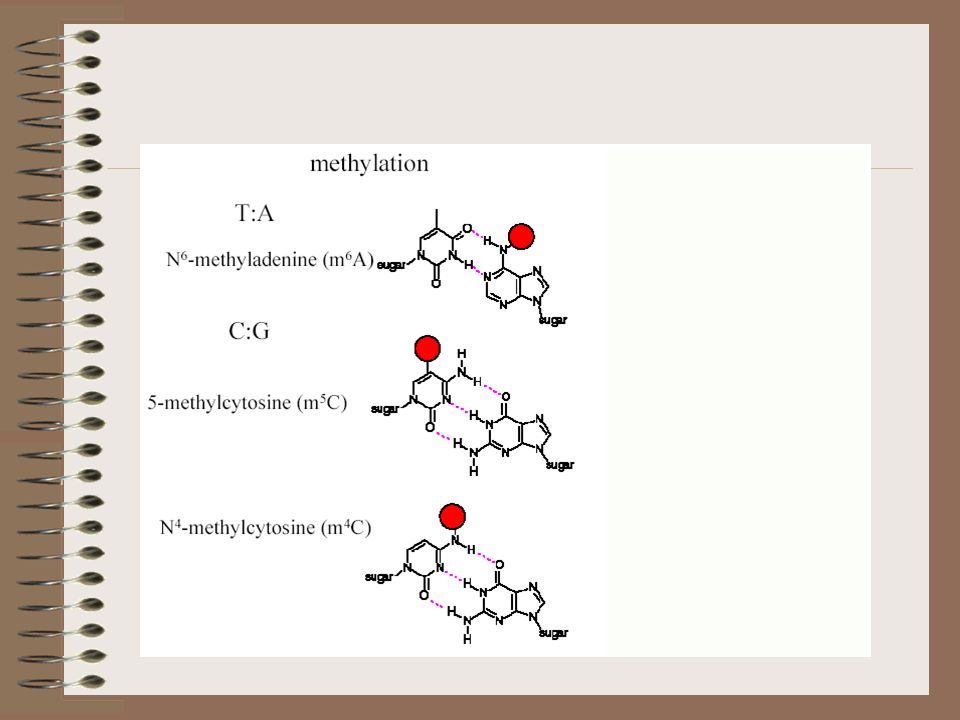
– trasferisce un metile dalla S-adenosil Metionina (SAM) a uno specifico nucleotide del sito di riconoscimento – La metliazione può dare origine a N6-metA, 5-metC, N4- metC – I gruppi metilici sono posizionati sul solco maggiore della doppia elica e non interferiscono con gli appaiamenti tra le basi del DNA – presumibilmente la metilazione impedisce il taglio da parte della RE mediante ingombro sterico o ostacolando il binding stesso della RE al sito
 a uno specifico nucleotide del sito di riconoscimento. – La metliazione può dare origine a N6-metA, 5-metC, N4- metC. – I gruppi metilici sono posizionati sul solco maggiore della doppia elica e non interferiscono con gli appaiamenti tra le basi del DNA. – presumibilmente la metilazione impedisce il taglio da parte della RE mediante ingombro sterico o ostacolando il binding stesso della RE al sito.")
10
Hsd, mcr e mrr mcrC mcrB hsdS hsdM hsdR mrr
I sistemi di restrizione in E. coli sono integrati in una “immigration control region” di circa 14 Kb DH1 e DH5 sono mutanti hsdR (EcoKI- ma mk+) DP50 è hsdS (quindi è EcoKI- sia rk- che mk-). Nota Bene: mutanti hsdS aboliscono sia la endonucleasi che la metilasi, mutanti hsdR solo la metilasi Alcuni ceppi come E coli C e HB101 (E coli B) sono Δ(mcrC-mrr) Hsd = Host Specificity for DNA Mcr/mrr = methylation dependent restriction endonucleases mcrC mcrB hsdS hsdM hsdR mrr
 DP50 è hsdS (quindi è EcoKI- sia rk- che mk-). Nota Bene: mutanti hsdS aboliscono sia la endonucleasi che la metilasi, mutanti hsdR solo la metilasi. Alcuni ceppi come E coli C e HB101 (E coli B) sono Δ(mcrC-mrr) Hsd = Host Specificity for DNA. Mcr/mrr = methylation dependent restriction endonucleases. mcrC mcrB hsdS hsdM hsdR mrr.")
11
Nomenclatura degli enzimi di restrizione
Per convenzione viene usato un acronimo di 3 lettere basato sul nome dell’organismo di provenienza. 1a lettera = genere, 2a e 3a = specie Dopo l’acronimo seguono una lettera che denota il ceppo e un numero romano che fa riferimento all’ordine cronologico di scoperta es. EcoRV (“eco erre cinque”) = Escherichia coli, ceppo R, 5° enzima scoperto
 = Escherichia coli, ceppo R, 5° enzima scoperto.")
12
Esempi di nomi di RE • BamHI =Bacillus amyloliquefaciens H, 1° enz
• DpnI = Diplococcus pneumoniae, 1° enz • SphI = Streptomyces phaeochromogenes, 1° enz • HindIII = Haemophilus influenzae Rd, 3° enz • BglII = Bacillus globigii, 2° enz • PstI = Providencia stuartii 164, 1° enz • Sau3A = Staphylococcus aureus 3A • KpnI =Klebsiella pneumoniae OK8, 1° enz

13
Enzimi tipo II e siti di riconoscimento con simmetria

14
I siti di restrizione degli enzimi tipo II sono
generalmente palindromici “I TOPI NON AVEVANO NIPOTI” “O MORDO TUA NUORA O ARO UN AUTODROMO” Una palindrome è una sequenza con un DOPPIO asse di simmetria: Bam H1 site: 5’-GGATCC-3’ 3’-CCTAGG-5’

15
Isoschizomeri - Neoschizomeri
Enzimi di restrizione differenti che riconoscono la stessa sequenza e la tagliano allo stesso modo Stesso sito riconosciuto: es. Sau3A, MboI, DpnI riconoscono tutti la seq /GATC Esistono però anche enzimi (neoschizomeri) che riconoscono lo stesso sito ma fanno un taglio diverso tra di loro: SmaI CCC/GGG = estremità piatte XmaI C/CCGGG = estremità 5’ sporgenti
 che riconoscono lo stesso sito ma fanno un taglio diverso tra di loro: SmaI CCC/GGG = estremità piatte. XmaI C/CCGGG = estremità 5’ sporgenti.")
16
Siti di riconoscimento e estremità compatibili
Tutte queste estremità “appiccicose” sono compatibili tra di loro BglII 5’A-G-A-T-C-T T-C-T-A-G-A 5’ Sau3A 5’G-A-T-C C-T-A-G 5’ BamHI 5’ G-G-A-T-C-C C-C-T-A-G-G 5’ Isoschizomeri: due o più enzimi differenti riconoscono lo stesso sito. (es. MboI taglia GATC, quindi è un isoschizomero di Sau3A)
")
17
Temperatura e attività degli enzimi RE tipo II

18
Frequenza di taglio degli enzimi RE tipo II
Sau 3A (GATC) taglia (1/4)(1/4)(1/4)(1/4) = ogni 256 bp (assumendo G/C = A/T, cosa generalmente non vera) BamH1 (GGATCC) taglia (1/4)(1/4)(1/4)(1/4)(1/4)(1/4) = ogni 4096 bp HindII (GTPyPuAC) taglia (1/4)(1/4)(1/2)(1/2)(1/4)(1/4) = ogni ~1Kb NotI (GC/GGCCGC) taglia (1/4)8 = ogni ~66Kbb I-CeuI (CGTAACTATAACGGTCCTAAGGTAGCGAA –9/-13) taglia ….. ogni qualche cromosoma ….. Esempio: il batterio Wolbachia ha un DNA con il 70% circa di A/T. Con che frequenza taglierà EcoRI?
 taglia (1/4)(1/4)(1/4)(1/4) = ogni 256 bp (assumendo G/C = A/T, cosa generalmente non vera) BamH1 (GGATCC) taglia (1/4)(1/4)(1/4)(1/4)(1/4)(1/4) = ogni 4096 bp. HindII (GTPyPuAC) taglia (1/4)(1/4)(1/2)(1/2)(1/4)(1/4) = ogni ~1Kb. NotI (GC/GGCCGC) taglia (1/4)8 = ogni ~66Kbb. I-CeuI (CGTAACTATAACGGTCCTAAGGTAGCGAA –9/-13) taglia ….. ogni qualche cromosoma ….. Esempio: il batterio Wolbachia ha un DNA con il 70% circa di A/T. Con che frequenza taglierà EcoRI")
19
Frequenza di taglio degli enzimi RE tipo II
Il batterio Wolbachia ha un DNA con il 70% circa di A/T. Ogni quanto taglierà EcoRI in media? Il sito di EcoRI è G/AATTC In Wolbachia A = T = 35% e C = G = 15% La possibilità che in prima base ci sia G non è più il 25% (1/4) ma 15%, cioè Analogamente che ci sia A in seconda base ha una probabilità del 35%, cioè 0.35 Considerato che queste probabilità sono indipendenti il calcolo è in conclusione 0.15 x 0.35 x 0.35 x 0.35 x 0.35 x 0.15 = Che equivale a 1/2961, UN TAGLIO OGNI 2961 bp
 ma 15%, cioè Analogamente che ci sia A in seconda base ha una probabilità del 35%, cioè Considerato che queste probabilità sono indipendenti il calcolo è in conclusione x 0.35 x 0.35 x 0.35 x 0.35 x 0.15 = Che equivale a 1/2961, UN TAGLIO OGNI 2961 bp.")
20
Estremità “adesive” o “sticky ends”
5’ sporgente (EcoRI) 5’-GAATTC-3’ ⇒ 5’-G-OH PO4-AATTC-3’ 3’-CTTAAG-5’ ’-CTTAA-PO HO-G-5’ + 3’ sporgente (PstI) 5’-CTGCAG-3’ ⇒ 5’-CTGCA-OH PO4-G-3’ 3’-GACGTC-5’ ’-G-PO HO-ACGTC-5’ + Estremità “piatte” o “blunt ends” SmaI 5’-CCC|GGG-3’ ⇒ 5’-CCC-OH PO4-GGG-3’ 3’-GGG|CCC-5’ ’-GGG-PO HO-CCC-5’ +
 5’-GAATTC-3’ ⇒ 5’-G-OH PO4-AATTC-3’ 3’-CTTAAG-5’ 3’-CTTAA-PO4 HO-G-5’ + 3’ sporgente (PstI) 5’-CTGCAG-3’ ⇒ 5’-CTGCA-OH PO4-G-3’ 3’-GACGTC-5’ 3’-G-PO4 HO-ACGTC-5’ + Estremità piatte o blunt ends SmaI. 5’-CCC|GGG-3’ ⇒ 5’-CCC-OH PO4-GGG-3’ 3’-GGG|CCC-5’ 3’-GGG-PO4 HO-CCC-5’ +")
21
RE tipo II - Utilizzi

22
_ + Elettroforesi su gel di agarosio
Il DNA ha una carica netta negativa che gli deriva dai fosfati e che lo fa “migrare” verso il polo positivo in un campo elettrico Gel di agarosio + Il DNA si visualizza grazie all’intercalante Bromuro di Etidio che assorbe UV e restituisce una fluorescenza nell’arancio (solo se è legato al DNA)
")
23
Figure 3.15 Partial unwinding of the DNA double helix by EtBr intercalation between adjacent base pairs. The normal DNA molecule shown on the left is partially unwound by taking up four EtBr molecules, resulting in the ‘stretched’ structure on the right. Gene Cloning and DNA Analysis by T.A. Brown. © 2006 T.A. Brown.

24
La velocità di migrazione del DNA è inversamente proporzionale al log delle dimensioni della molecola La quantità di carica negativa su una molecola di DNA è proporzionale alla sua lunghezza, MA il rapporto carica/massa è costante Dato che la densità di carica di un qualunque DNA è sempre la stessa, la separazione dei DNA in elettroforesi avviene in base alle loro dimensioni Le dimensioni di un DNA ignoto si possono ricavare confrontandone la migrazione su gel rispetto a quella di un DNA a dimensione nota

25
Le estremità compatibili sono saldabili
con la DNA ligasi DNA umano tagliato con EcoRI DNA batterico tagliato con EcoRI + 5’-C-G-G-T-A-C-T-A-G-OH 3’-G-C-C-A-T-G-A-T-C-T-T-A-A-PO4 PO4-A-A-T-T-C-A-G-C-T-A-C-G-3’ HO-G-T-C-G-A-T-G-C-5’ Le estremità si associano grazie alla complementarietà 5’-A-C-G-G-T-A-C-T-A-G A-A-T-T-C-A-G-C-T-A-C-G-3’ 3’-T-G-C-C-A-T-G-A-T-C-T-T-A-A G-T-C-G-A-T-G-C-5’ + DNA Ligasi, + ATP 5’-A-C-G-G-T-A-C-T-A-G-A-A-T-T-C-A-G-C-T-A-C-G-3’ 3’-T-G-C-C-A-T-G-A-T-C-T-T-A-A-G-T-C-G-A-T-G-C-5’ Molecola di DNA ricombinante

26
Saldatura mediante ligasi
Fagi - ATP T4 DNA ligasi E.Coli/Eucar - NAD Nicotinamide adenin dinucleotide

Presentazioni simili








 sottoposte a un campo elelttrico.>")







. Chimica del DNA i). Forze che influenzano la stabilità della doppia elica del DNA interazioni idrofobiche - stabilizzano dentro idrofobiche e fuori.>")


