Download presentation
1
La Teoria Chemiosmotica Premio Nobel in Chimica 1978 Prof. Peter Mitchell 1920-1992 Prof. Peter Mitchell 1920-1992 "for his contribution to the understanding of biological energy transfer through the formulation of the chemiosmotic theory"

2
La teoria Chemiosmotica

3
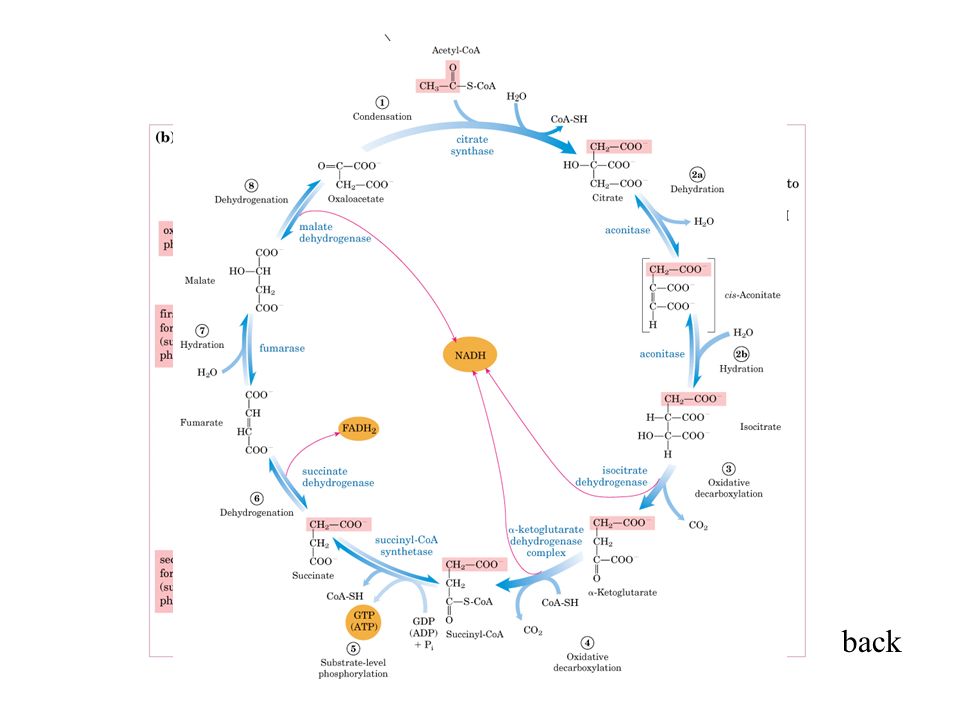
Proprieta’ comuni delle membrane trasducenti l’energia Una pompa protonica primaria, la cui natura dipende dalla fonte di energia utilizzata (irreversibile) Ogni membrana contiene 2 tipi di pompe protoniche distinte: Una pompa protonica secondaria detta ATP sintasi (reversibile)
 Ogni membrana contiene 2 tipi di pompe protoniche distinte: Una pompa protonica secondaria detta ATP sintasi (reversibile)")
4
Se l’ATP Sintasi fosse isolata e appropriatamente ricostituita in una membrana con il sito di binding dell’ATP all’esterno, l’aggiunta di ATP genererebbe un gradiente protonico (lume acido) ricostituzione dell’ATP sintasi ATP rimozione di ATP circuito chemiosmotico accoppiato: un vero e proprio circuito protonico
 ricostituzione dell’ATP sintasi ATP rimozione di ATP circuito chemiosmotico accoppiato: un vero e proprio circuito protonico")
5
Il circuito protonico è analogo ad un circuito elettrico circuito accoppiato circuito disaccoppiato (corto circuito)
")
6
Le membrane che trasducono l’energia contengono pompe protoniche con lo stesso orientamento topologico membrana mitocondriale interna membrana plasmatica batterica membrana plasmatica dei batteri fotosintetici membrana tilacoidale dei cloroplasti Complessi I, III e IV Batteriorodopsina e Centri di Reazione Batteriorodopsina e Centri di Reazione Centri fotosintetici

7
Basi storiche e sperimentali La ricerca di un intermedio organico ad alta energia che collegasse la pompa primaria a quella secondaria è stata lo stimolo centrale per la ricerca in bioenergetica negli anni ’50-’60. L’enigma centrale: questo ipotetico intermedio si è rivelato alquanto elusivo e non è mai stato identificato. Ogni teoria deve essere consistente con osservazioni sperimentali di base Nonostante l’enigma centrale le principali vie della trasduzione energetica erano note nei mitocondri

8
Osservazioni sperimentali Le catene di trasporto elettronico sono costituite da una serie di trasportatori di elettroni, con 3 regioni separate in cui l’energia redox può essere conservata nella sintesi di ATP La velocità della respirazione è controllata dalla richiesta di ATP (Controllo Respiratorio o RCR) L’accoppiamento fra respirazione e sintesi di ATP può essere interrotto (disaccoppiamento) mediante l’uso di composti noti come disaccoppianti che abbassano l’RCR (stimolano la respirazione in assenza di ATP o stimolano l’idrolisi dell’ATP) L’antibiotico oligomicina inibisce sia la sintesi che l’idrolisi dell’ATP L’energia respiratoria puo’ essere accoppiata anche al trasporto di Ca 2+ o alla riduzione del NAD(P) + Quest’ultimi processi possono essere indotti dall’idrolisi dell’ATP
 L’accoppiamento fra respirazione e sintesi di ATP può essere interrotto (disaccoppiamento) mediante l’uso di composti noti come disaccoppianti che abbassano l’RCR (stimolano la respirazione in assenza di ATP o stimolano l’idrolisi dell’ATP) L’antibiotico oligomicina inibisce sia la sintesi che l’idrolisi dell’ATP L’energia respiratoria puo’ essere accoppiata anche al trasporto di Ca 2+ o alla riduzione del NAD(P) + Quest’ultimi processi possono essere indotti dall’idrolisi dell’ATP")
9
Queste osservazioni erano consistenti con vie della trasduzione energetica che originavano da un intermedio ad alta energia comune

10
Postulati (requisiti) di Mitchell Le catene respiratorie e fotosintetiche trasportano protoni (O 2 pulse, uncouplers, H/e, 200 mv, SMP) L’ATP sintasi funziona come una ATPasi traslocante protoni Le membrane che trasducono l’energia hanno una conduttanza protonica effettiva molto bassa (grasso bruno) Le membrane che trasducono l’energia possiedono trasportatori specifici di scambio (carrier), che permettono la permeazione di metaboliti, il mantenimento della stabilità osmotica in presenza di un potenziale di membrana alto
 di Mitchell Le catene respiratorie e fotosintetiche trasportano protoni (O 2 pulse, uncouplers, H/e, 200 mv, SMP) L’ATP sintasi funziona come una ATPasi traslocante protoni Le membrane che trasducono l’energia hanno una conduttanza protonica effettiva molto bassa (grasso bruno) Le membrane che trasducono l’energia possiedono trasportatori specifici di scambio (carrier), che permettono la permeazione di metaboliti, il mantenimento della stabilità osmotica in presenza di un potenziale di membrana alto")
11
back

13
Batteriorodopsina da Halobacterium halobium Citocromo c ossidasi Paracoccus denitrificans back

15
Il trasporto di ioni attraverso le membrane trasducenti l’energia 1.Il trasporto ha luogo direttamente attraverso la membrana o e’ catalizzato da una proteina? 2.Il trasporto e’ passivo o accoppiato direttamente al metabolismo (trasporto attivo)? 3.Il trasporto coinvolge uno ione o metabolita singoli oppure il flusso di piu’ ioni o metaboliti sono direttamente accoppiati? 4.Il trasporto determina un trasferimento di carica attraverso la membrana? Qualsiasi processo di trasporto e’ descrivibile mediante 4 criteri
. 3.Il trasporto coinvolge uno ione o metabolita singoli oppure il flusso di piu’ ioni o metaboliti sono direttamente accoppiati. 4.Il trasporto determina un trasferimento di carica attraverso la membrana. Qualsiasi processo di trasporto e’ descrivibile mediante 4 criteri.")
16
Il trasporto ha luogo direttamente attraverso la membrana o e’ catalizzato da una proteina?

17
Il trasporto e’ passivo o accoppiato direttamente al metabolismo?

18
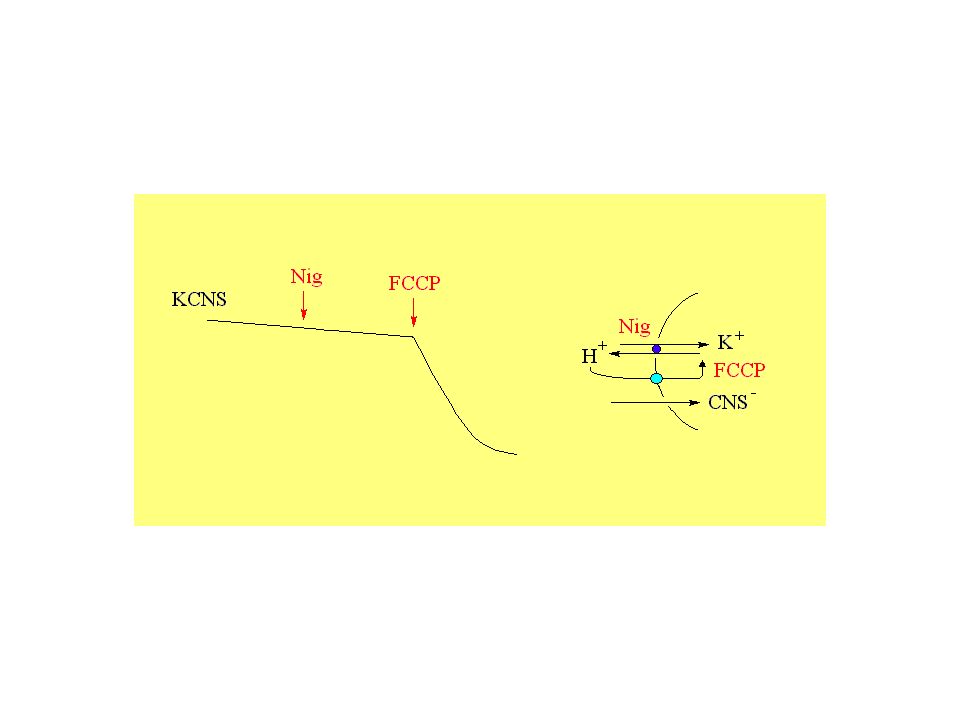
Il trasporto coinvolge uno ione o metabolita singoli oppure il flusso di piu’ ioni o metaboliti sono direttamente accoppiati? uniporto: trasporto del calcio nei mitocondri, protonofori simporto: trasporto del fosfato nei mitocondri antiporto: Na + /H + mitocondriale o K + /H + (nigericina)
.")
19
Il trasporto determina un trasferimento di carica attraverso la membrana?

20
L’ATP sintasi il trasporto di protoni è catalizzato da una proteina il trasporto è accoppiato al metabolismo (trasporto attivo) il trasporto è un uniporto il trasporto è elettrogenico L’ATP sintasi il trasporto di protoni è catalizzato da una proteina il trasporto è accoppiato al metabolismo (trasporto attivo) il trasporto è un uniporto il trasporto è elettrogenico Esempi La nigericina il trasporto è mediato dal doppio strato il trasporto è passivo il trasporto è un antiporto il trasporto è elettroneutro La nigericina il trasporto è mediato dal doppio strato il trasporto è passivo il trasporto è un antiporto il trasporto è elettroneutro
 il trasporto è un uniporto il trasporto è elettrogenico L’ATP sintasi il trasporto di protoni è catalizzato da una proteina il trasporto è accoppiato al metabolismo (trasporto attivo) il trasporto è un uniporto il trasporto è elettrogenico Esempi La nigericina il trasporto è mediato dal doppio strato il trasporto è passivo il trasporto è un antiporto il trasporto è elettroneutro La nigericina il trasporto è mediato dal doppio strato il trasporto è passivo il trasporto è un antiporto il trasporto è elettroneutro")
21
Trasporto mediato dalla membrana Il doppio strato idrofobico è una barriera per il passaggio di ioni (energia di attivazione alta) La membrana presenta una elevata resistenza elettrica e può sopportare campi elettrici giganteschi (con 200 mV si ha un campo elettrico di 300,000 V/cm) Molecole non cariche possono attraversare liberamente il doppio strato (H 2 O, O 2, CO 2, acidi e basi deboli) Trasporto diretto Trasporto mediato da ionofori La bassissima permeabilità ionica della membrana può essere aumentata mediante l’uso di ionofori (naturali o artificiali). Possiedono una parte idrofobica e una parte idrofilica Formano navette mobili nella membrana o canali e sono spesso molto selettivi per lo ione

22
Gli ionofori mobili Trasportano gli ioni per diffusione attraverso la membrana Tipicamente catalizzano il trasporto di 1000 ioni/s La selettività e discriminazione fra ioni e’ altissima Sono influenzati dalla fluidità della membrana

23
La Valinomicina: un uniporto elettrogenico per il potassio antibiotico naturale da Streptomyces è un depsipeptide ciclico (legami peptidici ed esterei) catalizza l’uniporto elettrogenico di Cs +, Rb +, K +, NH 4 + La capacità di trasportare il Na + è 10000 inferiore rispetto al K +
 catalizza l’uniporto elettrogenico di Cs +, Rb +, K +, NH 4 + La capacità di trasportare il Na + è inferiore rispetto al K +")
25
Struttura della valinomicina

26
Gli ionofori formanti canali Trasportano gli ioni per conduzione diretta attraverso lo ionoforo che forma un canale Catalizzano il trasporto di 10 7 ioni/s La selettività e discriminazione fra ioni non e’ alta Sono influenzati dalla fluidità della membrana

27
Struttura della gramicidina polipeptide costituito da L- e D-amminoacidi alternati, forma un elica sinistrorsa con 6.4 res/giro L-Val-Gly-L-Ala-D-Leu-L-Ala-D-Val-L-Val-D-Val-L-Trp-D-Leu- L-Trp-D-Leu-L-Trp forma un tubo lungo 26 Å: 2 polipeptidi con associaziobe testa- coda

28
Struttura della nigericina

29
Il meccanismo ad antiporto elettroneutro

30
I Protonofori Sono acidi deboli organici lipofilici La base coniugata è stabilizzata per risonanza e la solubilità nelle membrane è dovuta alla delocalizzazione della carica all’interno della molecola. Trasportano protoni mediante un meccanismo a navetta (uniporto elettrogenico) Sono acidi deboli organici lipofilici La base coniugata è stabilizzata per risonanza e la solubilità nelle membrane è dovuta alla delocalizzazione della carica all’interno della molecola. Trasportano protoni mediante un meccanismo a navetta (uniporto elettrogenico)
 Sono acidi deboli organici lipofilici La base coniugata è stabilizzata per risonanza e la solubilità nelle membrane è dovuta alla delocalizzazione della carica all’interno della molecola. Trasportano protoni mediante un meccanismo a navetta (uniporto elettrogenico).")
31
Carbonilcianuro-p-trifluorometossifenilidrazone

32
Il 2,4-dinitrofenolo

33
Funzionano mediante un meccanismo ad uniporto elettrogenico

34
I mitocondri sono organelli osmoticamente sensibili I mitocondri “scoppiano” se non posti in una soluzione isotonica con la matrice mitocondriale Spesso si utilizza una soluzione tamponata di saccarosio 0.25 M (non permeante) In presenza di un soluto permeante i mitocondri si “rigonfiano” (swelling) per il passaggio di acqua all’interno della matrice Esiste un metodo semplice per lo studio dello swelling basato sulla diffusione della luce
 In presenza di un soluto permeante i mitocondri si rigonfiano (swelling) per il passaggio di acqua all’interno della matrice Esiste un metodo semplice per lo studio dello swelling basato sulla diffusione della luce")
35
Lo Swelling

36
Il movimento coordinato di ioni attraverso le membrane Per osservare il rigonfiamento dei mitocondri, 2 condizioni debbono essere soddisfatte 1.Sia il catione che l’anione del componente osmoticamente significativo deve essere permeabile. 2.Il bilancio di carica transmembrana deve essere rispettato

37
Lo swelling non avviene finche’ non e’ presente un controione permeante