Scaricare la presentazione
1
Figura 14.2 Russell, Pearson
Esperimento di Stern: relazione fra scambio di materiale cromosomico e ricombinazione genetica in Drosophila Esperimenti di incrocio nei quali i cromosomi parentali differivano per marcatori genetici e citologici. Figura 14.2 Russell, Pearson

3
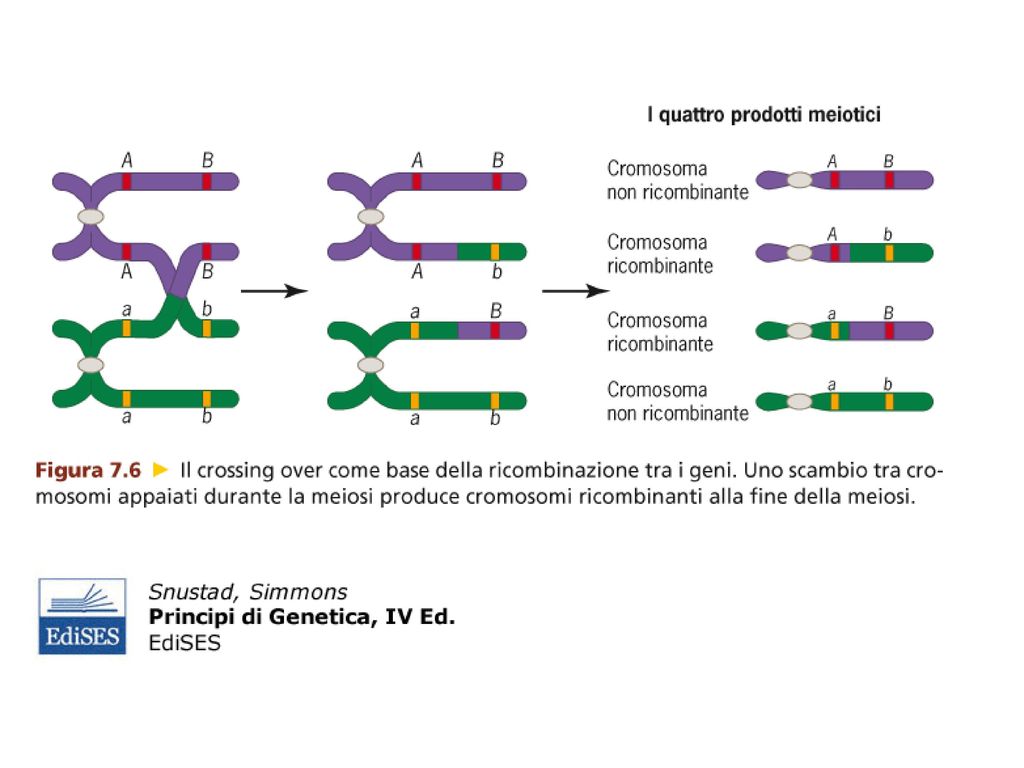
Questo esperimento ha dimostrato che la ricombinazione genetica (formazione di fenotipi ricombinanti o non parentali) ha origine dal crossing-over che determina uno scambio fisico tra cromosomi.
 ha origine dal crossing-over che determina uno scambio fisico tra cromosomi.")
4
Esperimento di Barbara McClintock (1931) in Zea mais
C: seme colorato, c: non colorato Wx: amido normale, wx: amido ceroso

5
Reincrocio ed esame della progenie ricombinante
Questi dati suggerirono in modo convincente che la ricombinazione era causata da uno scambio fisico o crossing over tra cromosomi appaiati. X C Wx c wx

6
Peter J Russell, Genetica © 2010 Pearson Italia S.p.A
Figura 14.7 Peter J Russell, Genetica © 2010 Pearson Italia S.p.A

7
Peter J Russell, Genetica © 2010 Pearson Italia S.p.A
Figura 14.8 Peter J Russell, Genetica © 2010 Pearson Italia S.p.A

8
Interferenza Se i crossing over, CO, fossero indipendenti:
P(doppio CO) = P(CO p-j) x P(CO j-r) = = 0.1 x 0.2 = 0.02 Doppi crossing over attesi, DCA = 500x0,02 = 10 Doppi crossing over osservati, DCO = 6 cc = coeff. di coincidenza = DCO/DCA = 0.6 I = interferenza = 1 – cc = 0.4
 = P(CO p-j) x P(CO j-r) = = 0.1 x 0.2 = Doppi crossing over attesi, DCA = 500x0,02 = 10. Doppi crossing over osservati, DCO = 6. cc = coeff. di coincidenza = DCO/DCA = 0.6. I = interferenza = 1 – cc = 0.4.")
10
Meccanismo molecolare della ricombinazione (Modello di Holliday)
Sono mostrati solo i due cromatidi che partecipano al crossing-over Rottura nel DNA a doppio filamento di uno dei cromatidi che prendono parte al crossing-over A partire dal punto di rottura il DNA viene eroso nel filamento 5’ di ciascuna estremità rotta lasciando entrambe le estremità 3’ a singolo filamento. Uno di questi filamenti invade il DNA dell’altro cromatidio cioè penetra all’interno dell’elica e si appaia alla sua sequenza omologa.

11
L’estremità del filamento che ha invaso utilizza la sequenza a cui si è appaiato come stampo per la nuova polimerizzazione che procede allontanando i due filamenti residenti dell’elica. Le estremità replicate vengono saldate e il risultato finale è una struttura strana con due giunzioni chiamate giunzioni di Holliday. Eventi di rottura e riunione portano a un completo crossing over

12
La maggior parte dei ceppi batterici può essere infettata da fagi specifici. Per Escherichia coli: T2, T4, T6 e λ. lisi batterica placca strato batterico continuo

13
Mutanti del fago T4 a lisi rapida r producono placche ampie con margini netti.
Il fago T4 selvatico produce placche piccole con margini frastagliati.

14
I mutanti rII Un tipo di mutanti r di T4 (i mutanti rII) sono letali condizionali. I mutanti rII possono crescere su alcuni ceppi di E. coli, come il ceppo B, ma non possono farlo (sono letali) su altri, come K12(),cellule K12 che contengono un profago .
 su altri, come K12(),cellule K12 che contengono un profago .")
15
Benzer e i suoi collaboratori isolarono un numero elevato di mutanti rII:
1) a causa della caratteristica morfologia di placca, i mutanti r sono facilmente distinguibili dai fagi selvatici 2) solo i mutanti rII sono incapaci di crescere su E. coli K12().
 a causa della caratteristica morfologia di placca, i mutanti r sono facilmente distinguibili dai fagi selvatici. 2) solo i mutanti rII sono incapaci di crescere su E. coli K12().")
16
Mappatura delle mutazioni rII

17
Frequenza di ricombinazione= 2X numero di ricombinanti selvatici
Numero totale della progenie








. La dominanza (primo principio di Mendel).>")













