Scaricare la presentazione
1
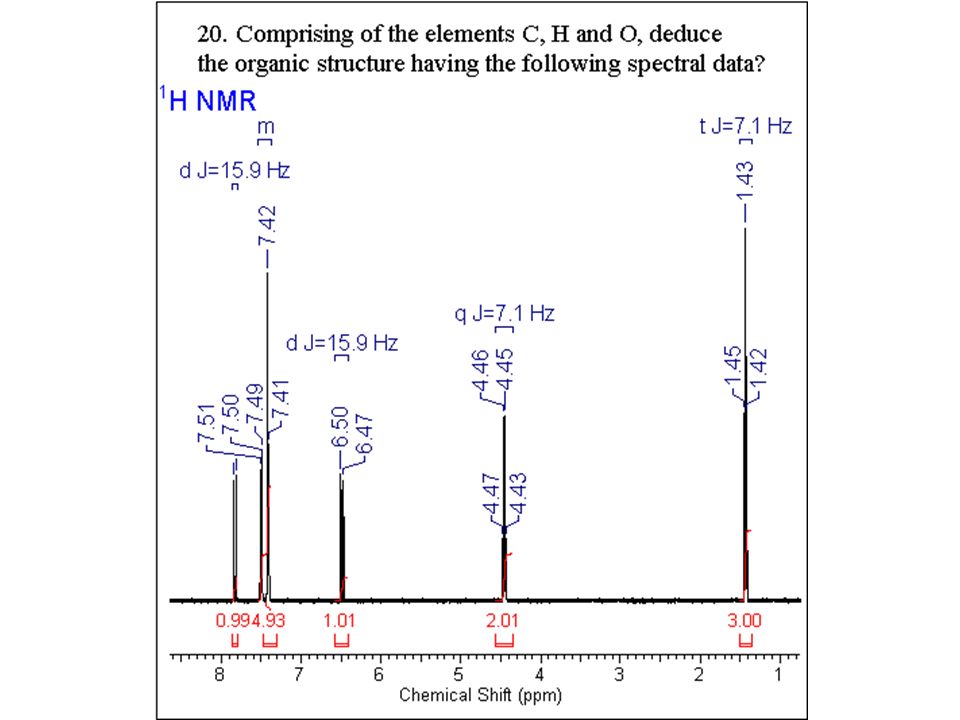
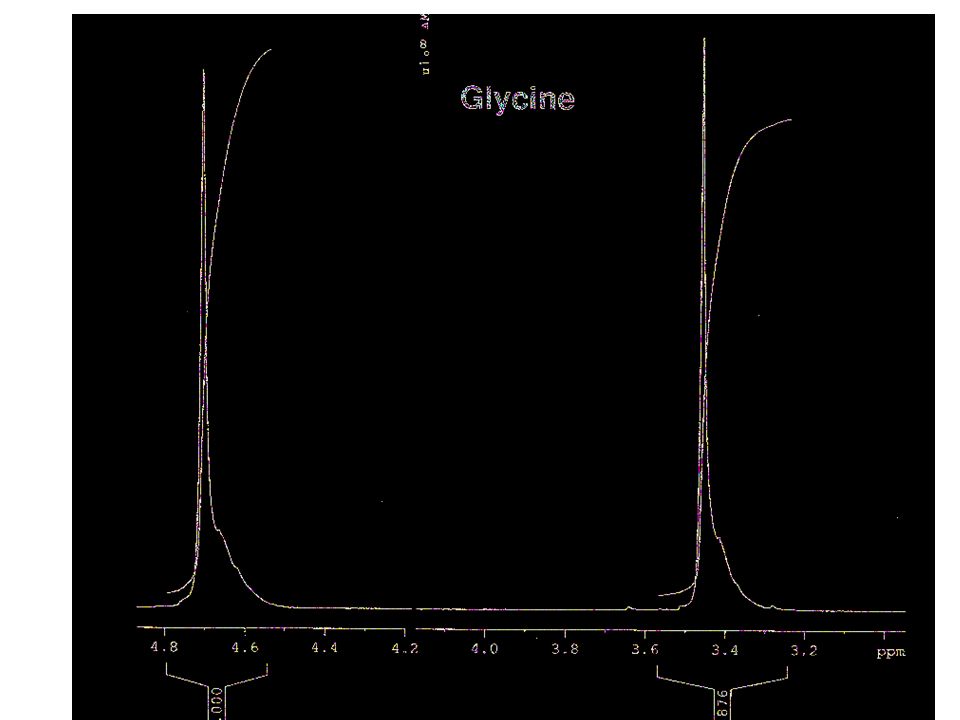
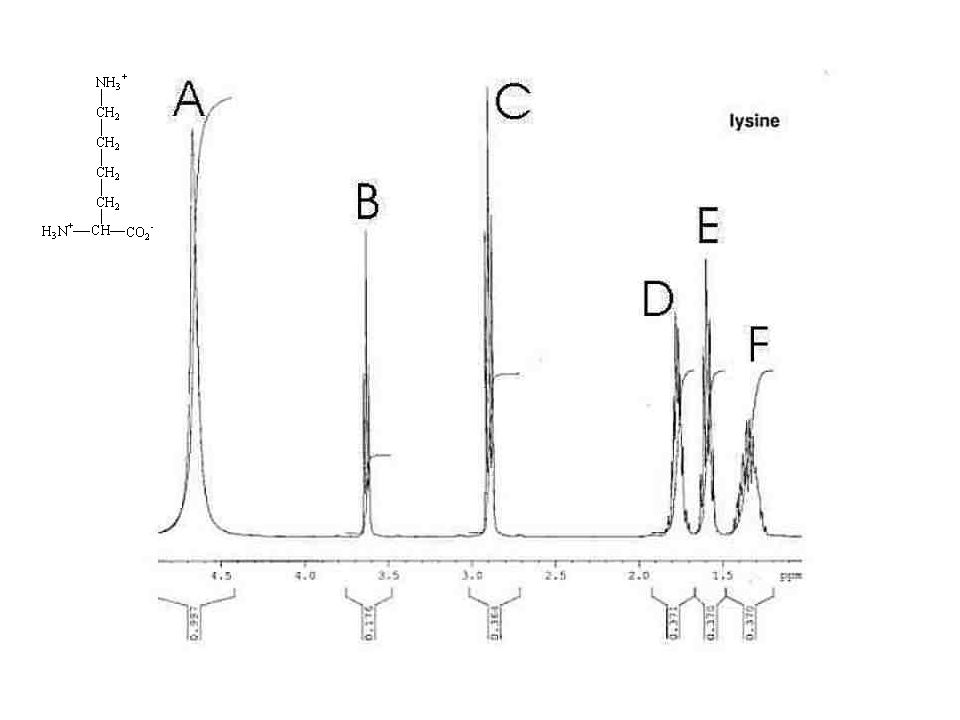
Some paradigmatic examples

3
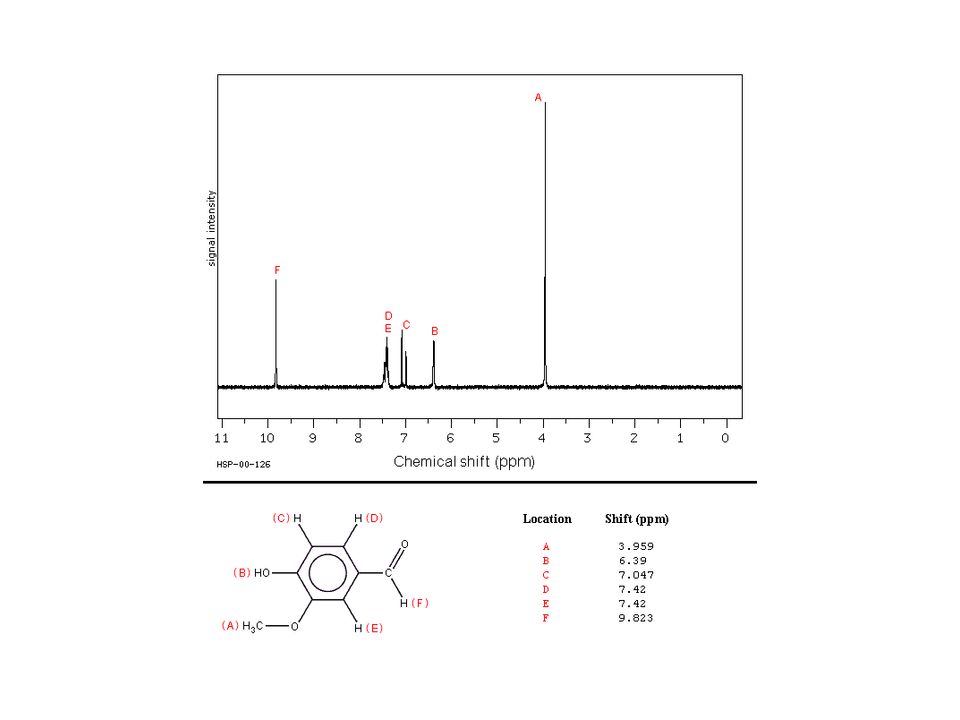
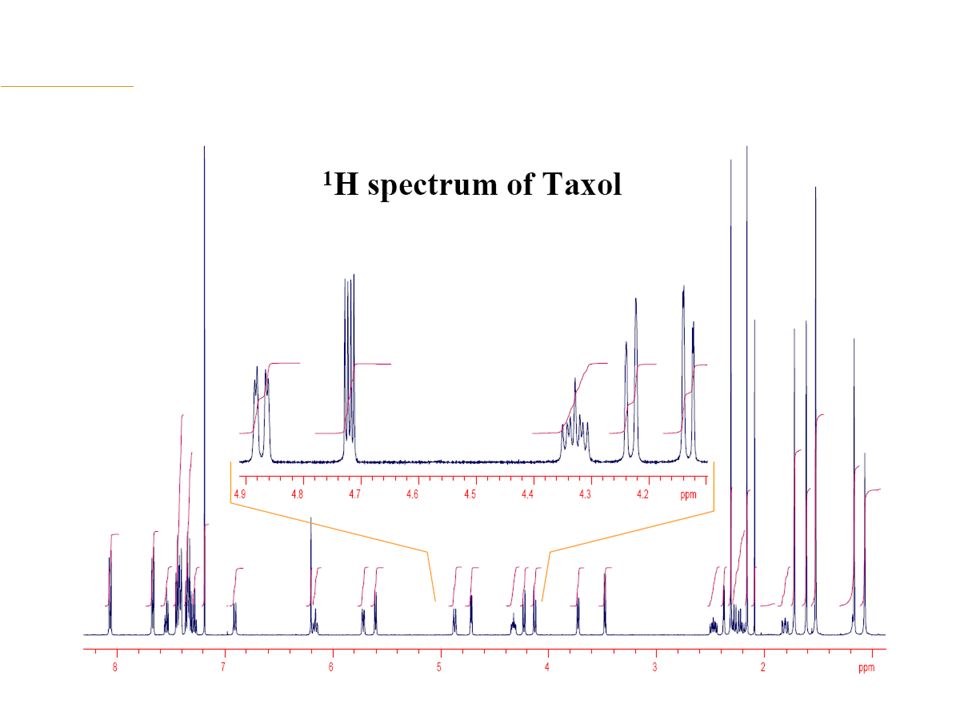
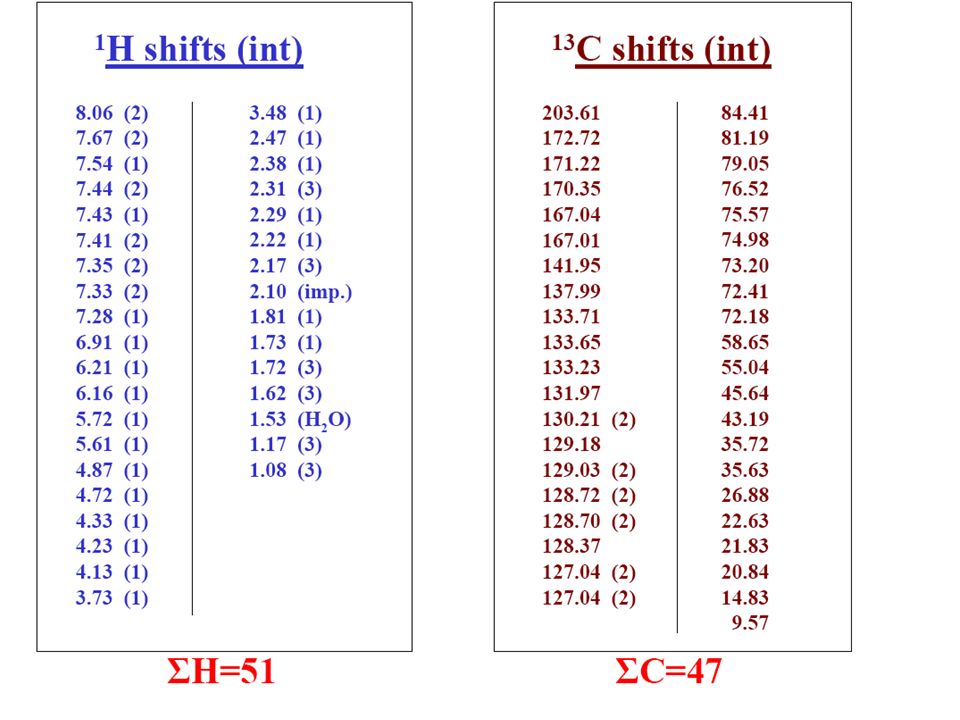
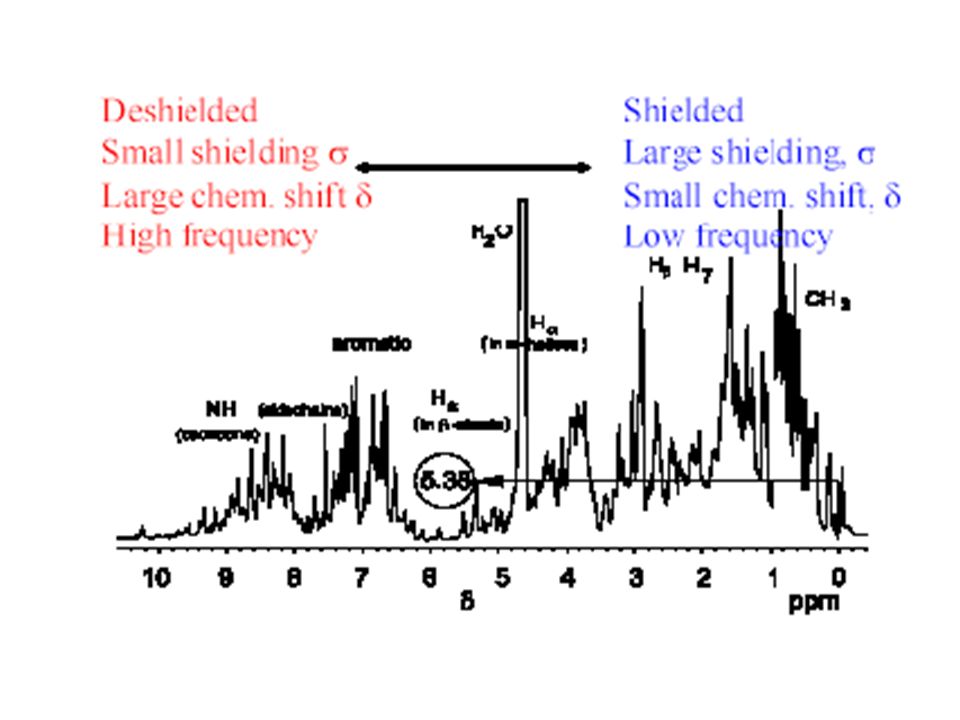
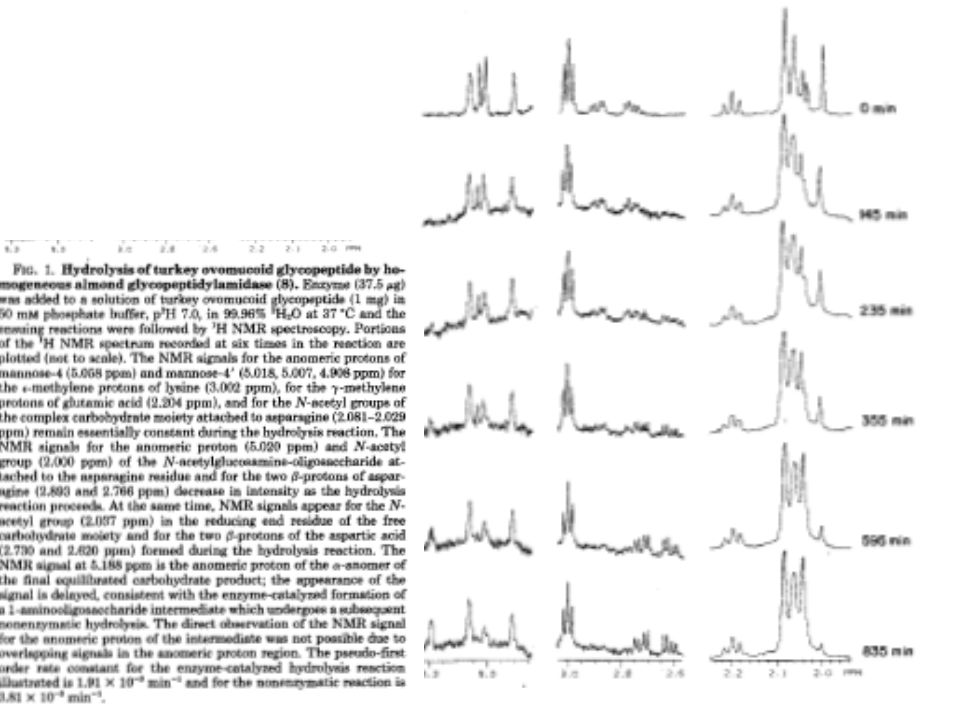
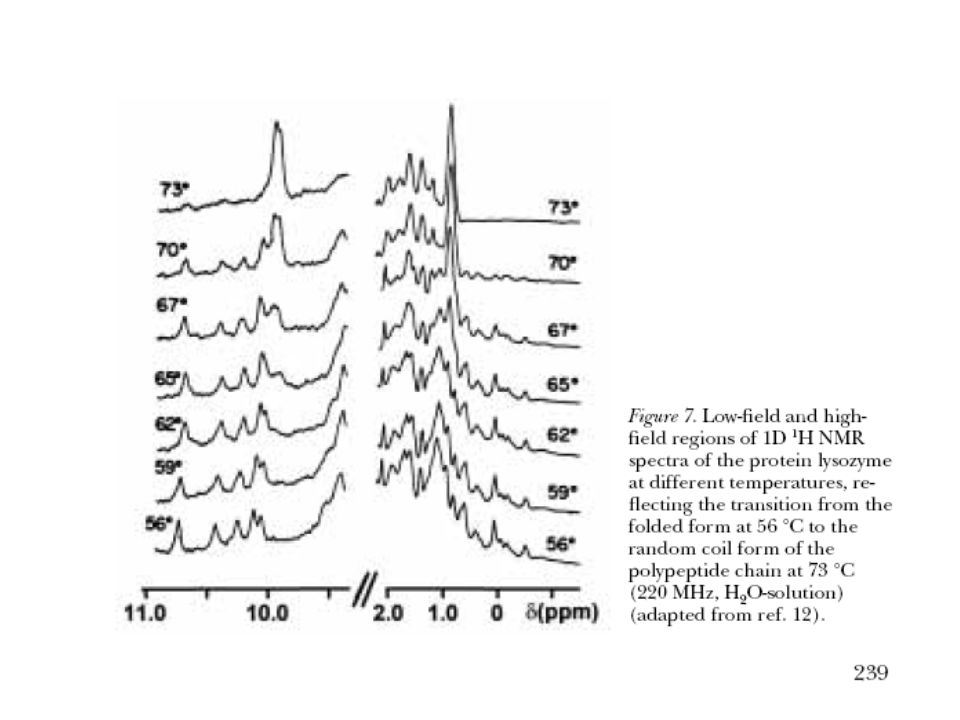
Typical 1H NMR Spectrum Absorbance

11
Valore medio: 2 MUSD/anno

12
500 MHz 1H NMR K1 T2 L3 T4 L5 E6 A7 A8 L9 R10 N11 A12 W13 L14 R15 E16 V17 G18 L19 K20

13
Ubiquitin 76 amino acids, 8,5 kDa

14
Protein 1H NMR spectrum: a “real spectrum”
The NMR signal in the time domain Free Induction Decay A short pulse will excite all spins All spins will relax (all together) during time AQ The FT of FID gives the NMR spectrum Fourier Transformation
 during time AQ. The FT of FID gives the NMR spectrum. Fourier Transformation.")
18
1D experiment ..Too crowded.. Chemical shifts relaxation rates
Could be nice but... Not enough to get a structure What do we learn?

19
STRUTTURE IN SOLUZIONE VIA NMR

20
The need for multidimensional NMR

22
Cosa è un esperimento bidimensionale ?
Dopo un impulso a 90° il segnale è pronto per essere acquisito Facciamo l’acquisizione ma NON terminiamo l’esperimento ed applichiamo ancora uno o piu’ impulsi in modo da perturbare ulterioremente il sistema Attraverso una combinazione di impulsi e delays noi facciamo in modo che ci sia uno scambio di magnetizzazione tra spin accoppiati SUCCESSIVAMENTE, acquisiamo il segnale una seconda volta, Registrando il segnale NMR che rimane sul piano xy dopo la perturbazione

23
Eccito (impulso a 90°)-Acquisisco (t1)- Perturbo (trasferisco)- Acquisisco (t2)
Se la perturbazione non ha effetto e se non c’è trasferimento di alcun tipo, Ottengo lo stesso spettro in ciascuna delle 2 dimensioni tempo (t1 e t2) Dopo la trasformate di Fourier io otterro’ uno spettro dove i segnali appaiono su una diagonale di una matrice quadrata
 Dopo la trasformate di Fourier io otterro’ uno spettro dove i segnali appaiono su una diagonale di una matrice quadrata.")
24
Acquisisco (t1)- Perturbo (trasferisco)- Acquisisco (t2)
Se durante la perturbazione una parte della magnetizzazione si traferisce da un nucleo ad un altro, per esempio per effetto di accoppiamento scalare, allora lo spettro della dimensione t2 sarà diverso da quello della dimensione t1. Il risultato è che avro’ dei segnali fuori dalla diagonale. Ciascun segnale fuori dalla diagonale darà l’informazione sugli accoppiamenti scalari attivi nel sistema M (wI t1) (wSt2)
 (wSt2)")
25
EXAMPLE O H H N C C We make a 1H experiment and we acquire. I observe Hn Then all signals transfer the information because of scalar coupling H H N C Spectrum before The J coupling Then I observe Hc I consider the first and the second acquisition as two indpendent dimensions Spectrum after The J coupling

26
EXAMPLE O H H N C C Signal! 9 ppm 4 ppm H H N C Spectrum before
This indicates that there is a scalar coupling between Hn and Hc H H N C Spectrum before The J coupling 9 ppm 4 ppm Spectrum after The J coupling

27
EXAMPLE O H H N C C Signal! 9 ppm 4 ppm Hn Hn Hc J-coupling H H N C
This indicates that there is a scalar coupling between Hn and Hc H H N C Spectrum before The J coupling 9 ppm 4 ppm Hn Hn Hc J-coupling Spectrum after The J coupling

28
EXAMPLE O H H N C C If you begin from Hc , the situation is the same !
Spectrum before The J coupling Hc Hc Hn J-coupling Spectrum after The J coupling

29
EXAMPLE O H H N C C Therefore, if I consider only this system Hc Hc Hn
Spectrum before The J coupling Hn J-coupling Hn Hn Hc J-coupling Spectrum after The J coupling

30
The second dimension = t2
The first dimension = t1 The second dimension = t2 the series of pulses that I have to apply to my system = PULSE SEQUENCE example t1 dimension Or F1 t1 t2 t2 dimension Or F2

31
Usually t1 is also defined as indirect dimension
t2 is also defined as direct dimension the series of pulses that I have to apply to my system = PULSE SEQUENCE example t1 dimension Or F1 t1 t2 t2 dimension Or F2

32
CROSS PEAK= Yes, There is a COUPLING between the two frequencies
Definitions Diagonal peak The same frequency is observed in both dimensions F1 Cross peak Two different frequencies are observed in the two dimensions F2 CROSS PEAK= Yes, There is a COUPLING between the two frequencies

33
Accoppiamento scalare
L’accoppiamento scalare puo’ comunque essere osservato attraverso esperimenti NMR bidimensionali, quali il COSY

34
COrrelation SpectroscopY
Example: COSY Through-bond connectivities H4’-H5’ H4-H5 COSY= COrrelation SpectroscopY

35
COrrelation SpectroscopY
Example: COSY Through-bond connectivities H4’-H5’ 2 4 3 H4-H5 5 6 COSY= COrrelation SpectroscopY 1

36
Beyond COSY COSY is not the only 2D experiment
It is possible to transfer the information from spin A to spin B via several possible mechanisms The most important routes, which is COMPLEMENTARY TO J-coupling Is THROUGH SPACE COUPLING

37
Accoppiamento dipolare
L’accoppiamento dipolare si ha tra due spin che sono vicini nello spazio Si tratta della interazione tra due dipoli magnetici, tra i quali, quando essi sono vicini nello spazio, si ha uno scambio di energia L’entità dell’effetto dipende dal campo magnetico e dalle dimensioni della molecola. Nel caso di spin 1H, l’accoppiamento dipolare si trasferisce per spin che si trovano a distanze inferiori ai 5 A. NON si osservano doppietti L’accoppiamento dipolare da luogo ad un trasferimento di magnetizzazione da uno spin all’altro. Questo effetto va sotto il nome di effetto NOE Nuclear Overhauser Effect Aumenta la intensità di B Perturbo A

38
Accoppiamento dipolare
L’accoppiamento dipolare è “indipendente dall’accoppiamento scalare 2 spin possono essere accoppiati : -Scalarmente E dipolarmente se sono vicini nello spazio e legati da legami chimici -scalarmente ma non dipolarmente se sono legati da legami chimici ma non vicini nello spazio -dipolarmente ma non scalarmente se sono spazialmente vicini ma non legati da legamei chimici L’effetto NOE è osservabile in un esperimento NMR bidimensionale , detto NOESY (in realtà si puo’ anche osservare in esperimenti monodimensionle (1D NOE) di cui pero’ non parleremo Pensate a degli esempi, per favore
 di cui pero’ non parleremo. Pensate a degli esempi, per favore.")
39
Through space AND throuhg bonds
Through bond

40
Nuclear Overhauser Effect SpectroscopY
Example: Nuclear Overhauser Effect SpectroscopY NOESY NOE Effect: If two spins that are close in space are excited out of equilibrium, they will mutually transfer their magnetization AA AB

41
Example: The real case:
Some 1500 peaks are observed for a protein of 75 aminoacids NOESY experiment Cross peaks: A and B are close Diagonal peak AA AB

42
2D NOESY Spectrum

43
Distance constraints mJ B0 mI q r
NOESY volumes are proportional to the sixth power of the interproton distance and to the correlation time for the dipolar coupling mJ B0 mI q r

44
The “old times” approach
NOESY Identify through space connectivities HN(i)-Ha(i) and HN(i)Ha(i-1) Identify through bond connectivities HN(i)-Ha(i) COSY NOESY conn. COSY conn
-Ha(i) and HN(i)Ha(i-1) Identify through bond connectivities. HN(i)-Ha(i) COSY. NOESY conn. COSY conn.")
47
1J couplings for backbone resonances

48
1J couplings for backbone resonances

49
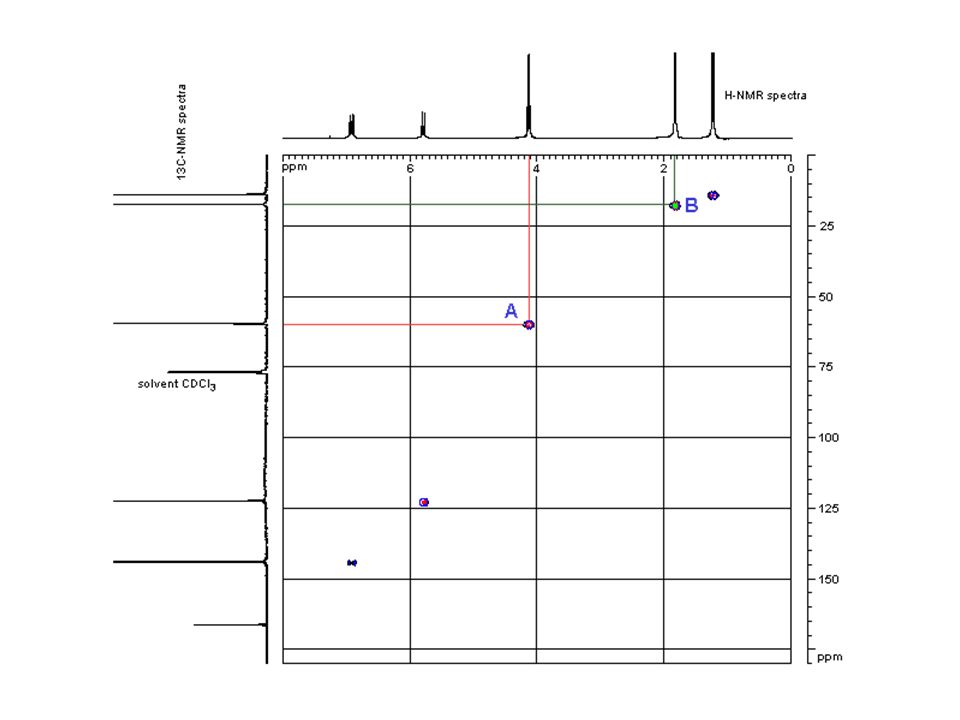
The 2D Hetcor experiment
Two dimensional Heteronuclear correlation Experiment

50
The 2D Hetcor experiment
Two dimensional Heteronuclear correlation Experiment

51
E’ possibile, in uno stesso esperimento mandare impulsi su nuclidi diversi (Es: 1H, 13C)
’ possibile, combinare questa possibilità con ciò che sappiamo a proposito degli accoppiamenti scalari e quindi UTILIZZARE gli accoppiamenti scalari per trasferire la magnetizzazione dauno spin 1H ad uno spin 13C ad esso scalarmente accoppiato

52
E’ possibile, in uno stesso esperimento mandare impulsi su nuclidi diversi (Es: 1H, 13C)
Inoltre possiamo combinare tutto cio’ con quello che sappiamo sugli esperimenti bidimensionali

53
2D HETCOR Expriment Eccito (impulso a 90°) 1H Acquisisco (t1) 1H –
Perturbo (Trasferisco la magnetizzazione da 1H a 13C utilizzando l’accoppiamento scalare 1JHC Acquisisco (t2) 13C
 13C.")
54
2D HETCOR Expriment

55
2D HETCOR Expriment Prima dimensione

56
2D HETCOR Expriment Prima dimensione

57
2D HETCOR Expriment Prima dimensione Seconda dimensione

58
2D HETCOR Expriment Prima dimensione Seconda dimensione

60
Esempio COSY

61
Esempio COSY N.B. In questo caso non si osserva solo l’accoppiamento 3J ma si osserva una “propagazione” dell’informazione attraverso gli accoppiamenti scalari

62
Esempio HETCOR 4 3 1 5 2

63
2D HSQC Experiment Heteronuclear Single Quantum coherence

64
2D HSQC Expriment Heteronuclear Single Quantum coherence

65
2D HSQC Experiment Heteronuclear Single Quantum coherence
Seconda dimensione Prima dimensione

66
2D HSQC Expriment Heteronuclear Single Quantum coherence
Prima dimensione

67
2D HSQC Experiment Heteronuclear Single Quantum coherence
Seconda dimensione Prima dimensione

68
2D HSQC Experiment Heteronuclear Single Quantum coherence
E’ possibile progettare esperimenti per trasferire la magnetizzazione da un nucleo all’altro anche indipendentemente dall’acquisizione Seconda dimensione Prima dimensione In questo esperimento il primo spin che viene eccitato è 1H, la magnetizzazione viene trasferita da 1H a 13C PRIMA della acquisizione della prima dimensione, che quindi è 13C. SOLO i 13C che sono accoppiati ad 1H possono essere osservati! Successivamente la magnetizzazione e di nuovo trasferita 1H utilizzando sempre l’accoppiamento scalare ed alla fine osservo 1H

69
2D HSQC Experiment Eccito (impulso a 90°) 1H Trasferisco la magnetizzazione da 1H a 13C utilizzando l’accoppiamento scalare 1JHC Acquisisco (t1) 13C – Perturbo -Trasferisco la magnetizzazione da 13C a 1H utilizzando l’accoppiamento scalare 1JHC Acquisisco (t2) 1H
 13C – Perturbo -Trasferisco la magnetizzazione da 13C a 1H utilizzando l’accoppiamento scalare 1JHC. Acquisisco (t2) 1H.")
70
2D HSQC Experiment Questo tipo di esperimento si chiama anche
Out and back Significa che parto da 1H, trasferisco da 1H a 13C (out), acquisisco 13C nella prima dimensione e poi torno (back) sullo stesso nucleo da cui sono partito Il doppio trasferimento fa si che l’esperimento sia molto piu’ selettivo Osservo solo 1H e 13C che sono accoppiati tra di se per effetto di 1J
, acquisisco 13C nella prima dimensione e poi torno (back) sullo stesso nucleo da cui sono partito. Il doppio trasferimento fa si che l’esperimento sia molto piu’ selettivo. Osservo solo 1H e 13C che sono accoppiati tra di se per effetto di 1J.")
71
The HSQC experiment

72
Caratteristiche dell’esperimento HSQC
Non esiste la diagonale La magnetizzazione viene trasferita da 1H al 13C ad esso accoppiato Successivamente si acquisisce, nella dimensione indiretta, 13C Infine si ri-trasferisce su 1H e si osserva 1H Tutti gli 1H che non sono accoppiati a 13C NON si osservano

73
Heteronuclear NMR Transfer the information to all 1H coupled
OBSERVE 13C during t1 Transfer the information to all 1H coupled OBSERVE 1H during t2 13C No more diagonal Each peak indicate A different H-C pair 1H

74
Heteronuclear NMR OBSERVE 13C during t1
Transfer the information to all 1H coupled Observe 1H during t2 No more diagonal CH2 Two protons are bound to the same carbon 13C 1H

75
The HSQC experiment

76
Heteronuclear cases The scheme of 1J scalar couplings

77
The 1H- 15N HSQC experiment
Heteronuclear Single Quantum Coherence

78
The HSQC experiment Detect H-N couplings
Each amide NH group gives rise to one peak Same sensitivity of a 1H experiment (although you are observing 15N) but much larger resolution In 5 minutes you may know…. if your protein is properly folded if all aminoacids gives rise to an observable peak if you can do the job (whatever is your job)
 but much larger resolution. In 5 minutes you may know…. if your protein is properly folded. if all aminoacids gives rise to an observable peak. if you can do the job. (whatever is your job)")
79
Heteronuclear NMR in proteins example: 15N labelled proteins

80
Heteronuclear NMR in proteins example: 15N labelled proteins

81
The HSQC experiment Detect H-N couplings
Each amide NH group gives rise to one peak Same sensitivity of a 1H experiment (although you are observing 15N) but much larger resolution In 5 minutes you may know…. if your protein is properly folded if all aminoacids gives rise to an observable peak
 but much larger resolution. In 5 minutes you may know…. if your protein is properly folded. if all aminoacids gives rise to an observable peak.")
82
The role of metal cofactor in protein unfolding
Metal triggered protein folding Apo 3.3 M GdmCl 3.3 M GdmCl Ca2+ Loss of secondary structure elements: unfolded protein Refolding

83
Apo vs holo protein, mapping the environment of the metal ion
95 77 60 20 59 HNd 28 35 15 6 26 45 96 7 90 HNe 32 75 72 94 33 37 13 12 8 3 78 65 28 34 100 88 23 41 36 46 83 64 27 80 89 48 62 99 14 84 21 17 43 24 25 74 29 86 30 79 47 51 58 16 42 87 22 4 11 98 39 10 76 69 97 53 91 81 93 56 101 40 67 57 19 38 71 52 55 68 54 85 66 82 5 HNe1 83 102 15H

84
The need for multidimensional NMR

85
Troppi segnali 1H ?

86
Isotope labeling For biomolecules, tipically, 15N or 13C and 15N, or 13C, 15N, 2H 15N Only A more effective fingerprint -characterization -folding -dynamics protein size >10000 Homonuclear 2D experiments do not have enough resolution HSQC or HMQC HSQC-NOESY or HSQC TOCSY

87
Isotope labeling For biomolecules, tipically, 15N or 13C and 15N, or 13C, 15N, 2H protein size >20000 15N and 13C Scalar couplings through 13C atoms -triple resonance -assignment -structure

88
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Expression systems are based on the insertion of a gene into a host cell for its translation and expression into protein . Many recombinant proteins can be expressed to high levels in E. coli systems. most common choice for expressing labeled proteins for NMR Yeast (Pichia pastoris, Saccaromyces cerevisiae) is an alternative choice for NMR protein samples issues with glycosolyation of protein, which is not a problem with E. coli. choice between E. coli and yeast generally depend on personal experience. Insect cells (Baculovirus) and mammalian cell lines (CHO) are very popular expression systems that are currently not amenable for NMR samples no mechanism to incorporate isotope labeling or the process is cost prohibitive 15N labeling in CHO cells can cost $ K!
 is an. alternative choice for NMR protein samples. issues with glycosolyation of protein, which is not. a problem with E. coli. choice between E. coli and yeast generally depend. on personal experience. Insect cells (Baculovirus) and mammalian cell lines. (CHO) are very popular expression systems that are. currently not amenable for NMR samples. no mechanism to incorporate isotope labeling or. the process is cost prohibitive. 15N labeling in CHO cells can cost $ K!")
89
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression First step of the process involves the insertion of the DNA coding region of the protein of interest into a plasmid. plasmid - small, circular pieces of DNA that are found in E. coli and many other bacteria generally remain separate from the bacterial chromosome carry genes that can be expressed in the bacterium plasmids generally replicate and are passed on to daughter cells along with the chromosome Plasmids are highly infective, so many of the bacteria will take up the particles from simple exposure. Treating with calcium salts make membranes permeable and increase uptake of plasmids Plasmids used for cloning and expressing proteins are modified natural vectors more compact and efficient unnecessary elements removed Some Common plasmids pBR322 pUC19 pBAD large collections of plasmids with unique features and functions see:

90
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Basic Features of a Plasmid Defined region with restriction sites for inserting the DNA Gene that provides antibiotic resistance (ampicillin resistance in this case) replication is initiated
 replication is initiated.")
91
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Restriction Enzymes Recognizes and cuts DNA only at particular sequence of nucleotides blunt end – cleaves both ends sticky ends – cleaves only one strand Complimentary strand from DNA insert will “match” sticky end and insert in plasmid followed by ligation of the strands (T4 DNA Ligase)
")
92
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Restriction Enzymes Very large collection of restriction enzymes that target different DNA sequences

93
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Restriction Enzymes Restriction Map of plasmid showing the location where all restriction enzymes will cleave. allows determination of where & how to insert a particular DNA sequence want a clean insertion point, don’t want to cleave plasmid multiple times

94
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Next step of the process involves getting E. coli to express the protein from the plasmid. this occurs by the position of a promoter next to the inserted gene two common promoters are lac complex promoter T7 promoter lac complex promoter: Transcription is simply switched on by the addition of IPTG (isopropyl β-D-thiogalactoside) to remove LacI repressor protein. IPTG binds LacI which no longer binds the promoter region allowing transcription to occur
 to remove LacI repressor protein. IPTG binds LacI which no longer binds the promoter region allowing transcription to occur.")
95
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression T7 promoter: Again, transcription is switched on by the addition of IPTG to remove LacI repressor protein. IPTG binds LacI which no longer binds the promoter region allowing transcription/production of T7 RNA polymerase to occur. T7 RNA polymerase binds the T7 promoter in the plasmid to initiate expression of the protein two-step process leads to an amplification of the amount of gene product - produce very high quantities of protein.

96
Cell growth in a Shake flask
Introduction to Isotope Labeling of Proteins For NMR Overview of Protein Expression Next step of the process involves growing the E. coli cells Shake Flask cells are place in a “growth media” that provides the required nutrients to the cell amino acids, vitamins, growth factors, etc shake the flask at a constant temperature of 37O keeps homogenous mixture increases oxygen uptake grow cells to proper density (OD ~ 0.7 at 600nm) LB Broth Recipe (Luria-Bertani) 10 g tryptone 5 g of yeast extract 10 g of NaCl Cell growth in a Shake flask
 LB Broth Recipe (Luria-Bertani) 10 g tryptone 5 g of yeast extract 10 g of NaCl. Cell growth in a Shake flask.")
97
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Next step of the process involves growing the E. coli cells Bioreactors more efficient higher production volumes can be 100s of liters in size Can grow cells to a higher density better control of pH better control of oxygen levels better control of temperature better control of mixing sterile conditions 14 liter bioreactor

98
Biotechnology Letters (1999) 12,1131
Introduction to Isotope Labeling of Proteins For NMR Overview of Protein Expression Next step is to harvest and lysis the cells and purify the protein Now that E. coli is producing the desired protein, need to extract the protein from the cell and purify it. the amount of protein that can be obtained from an expression system is highly variable and can range from mg to mg to even g quantities. it depends on the behavior of the protein, expression level, method of fermentation and the amount of cells grown over-expressed protein Biotechnology Letters (1999) 12,1131
 12,1131.")
99
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Cell Lysis A number of ways to lysis or “break” open a cell Gentle Methods Osmotic – suspend cells in high salt Freeze-thaw – rapidly freeze cells in liquid nitrogen and thaw Detergent – detergents (DSD) solubilize cellular membranes Enzymatic – enzymatic removal of the cell wall with lysozyme Vigorous Methods Sonication – sonicator lyse cells through shear forces French press – cells are lysed by shear forces resulting from forcing cell suspension through a small orifice under high pressure. Grinding – hand grinding with a mortar and pestal Mechanical homogenization - Blenders or other motorized devices to grind cells Glass bead homogenization - abrasive actions of the vortexed beads break cell walls French Press
 solubilize cellular membranes. Enzymatic – enzymatic removal of the cell wall with lysozyme. Vigorous Methods. Sonication – sonicator lyse cells through shear forces. French press – cells are lysed by shear forces resulting from forcing cell. suspension through a small orifice under high pressure. Grinding – hand grinding with a mortar and pestal. Mechanical homogenization - Blenders or other motorized devices to grind cells. Glass bead homogenization - abrasive actions of the vortexed beads break. cell walls. French Press.")
100
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Protein Purification - A large number of ways to purify a protein protocols are dependent on the protein chromatography is a common component of the purification protocol where typically multiple columns are used: a) size-exclusion b) ion exchange c) Ni column d) heparin e) reverse-phase f) affinity column dialysis for buffer exchange and removal of low-molecular weigh impurities To increase the ease of purifying a protein generally include a unique tag sequence at the N- or C-terminus HIS tag – add 6 histidines to the N- or C- terminus - preferentially binds Ni column FLAG tag – DYKDDDDK added to terminus - preferentially binds M1 monoclonal antibody affinity column glutathione S-transferase (GST) tags – fusion protein - readily purified with glutathione-coupled column
 size-exclusion b) ion exchange. c) Ni column d) heparin. e) reverse-phase f) affinity column. dialysis for buffer exchange and removal of low-molecular weigh. impurities. To increase the ease of purifying a protein generally include a unique. tag sequence at the N- or C-terminus. HIS tag – add 6 histidines to the N- or C- terminus. - preferentially binds Ni column. FLAG tag – DYKDDDDK added to terminus. - preferentially binds M1 monoclonal antibody affinity. column. glutathione S-transferase (GST) tags – fusion protein. - readily purified with glutathione-coupled column.")
101
Introduction to Isotope Labeling of Proteins For NMR
Overview of Protein Expression Some Common Problems protein is not soluble and included in inclusion bodies insoluble aggregates of mis-folded proteins inclusion bodies are easily purified and can be solubilized using denaturing conditions How to re-fold the Protein? Finding a re-folding protocol may take significant effort (months-years?) and involve numerous steps something to be avoided if possible protein is toxic to cell find a different expression vector or use a similar protein from a different organisim proper protein fold proper disulphide bond formation – may need to re-fold the protein presence of tag may inhibit proper folding – may need to remove the tag low expression levels try different plasmid constructs try different protein sequences
 and. involve numerous steps. something to be avoided if possible. protein is toxic to cell. find a different expression vector or use a similar protein from a different. organisim. proper protein fold. proper disulphide bond formation – may need to re-fold the protein. presence of tag may inhibit proper folding – may need to remove the tag. low expression levels. try different plasmid constructs. try different protein sequences.")
102
Journal of Biomolecular NMR, 20: 71–75, 2001.
Introduction to Isotope Labeling of Proteins For NMR 13C and 15N Isotope Labeling of the protein cells need to be grown in “minimal media” use 13C glucose to achieve ~ 100% uniformed 13C labeling of protein use 15N NH4Cl to achieve ~ 100% uniformed 15N labeling of protein glucose and NH4Cl are sole source of carbon and nitrogen in “minimal media” E. coli uses glucose and NH4Cl to synthesize all amino-acids protein added prior to expressing protein of interest both 13C glucose and 15N NH4Cl can be added simultaneously Journal of Biomolecular NMR, 20: 71–75, 2001.

103
1H-15N HSQC spectra of 13C,15N labeled protein
Introduction to Isotope Labeling of Proteins For NMR 13C and 15N Isotope Labeling of the protein Usually isotope labeling does not negatively impact protein expression Some Common Problems with Isotope Labeling Problems “minimal media” stresses cells slower growth typically lower expression levels isotope labeling of All proteins minimal isotope affect may affect enzyme activities isotope labeling of expressed protein may affect protein’ properties solubility? proper folding? 1H-15N HSQC spectra of 13C,15N labeled protein

104
Introduction to Isotope Labeling of Proteins For NMR
13C and 15N Isotope Labeling of the protein Can introduce specific amino acid labels A variety of 13C and 15N labeled amino acids are commercial available Add saturating amounts of 19 of 20 amino acids to minimal growth media Add 13C and 15N labeled amino acid prior to protein expression media is actually very rich and the cells grow very well cells exclusively use the supplied amino-acids to synthesize proteins all of the occurrences of the amino-acid are labeled in the protein may be some additional labeled residues if the labeled amino acid is a precursor in the synthesis of other amino acids. 1H-15N-HSQC of His, Tyr & Gly labeled SH2-Domain no mechanism to label one specific amino acid i.e Gly-87

105
Introduction to Isotope Labeling of Proteins For NMR
13C and 15N Isotope Labeling of the protein Can label specific segment in protein use peptide splicing element intein (Protozyme) inteins are insertion sequences which are cleaved off after translations preceding and succeeding fragments are ligated extein Protein of Interest 15N-labeled J. Am. Chem. Soc. 1998, 120,
 inteins are insertion sequences which are cleaved off after translations. preceding and succeeding fragments are ligated extein. Protein of Interest. 15N-labeled. J. Am. Chem. Soc. 1998, 120,")
106
Introduction to Isotope Labeling of Proteins For NMR
13C and 15N Isotope Labeling of the protein Can also label only one component of a complex simply mix unlabeled and labeled components to form the complex greatly simplifies the NMR spectra only “see” 13C, 15N NMR resonances for labeled component of complex can see interactions (NOEs) between labeled and unlabeled compoents J. OF BIOL. CHEM. (2003) 278(27), 25191–25206
 between labeled and unlabeled compoents. J. OF BIOL. CHEM. (2003) 278(27), 25191–")
107
Introduction to Isotope Labeling of Proteins For NMR
2H Labeling of the protein simply requires growing the cells in D2O severe isotope effect for 1H2H stresses the cell E. coli needs to be acclimated to D2O pass cells into increasing percentage of D2O cell growth slows significantly in D2O (18-60 hrs) level of 2H labeling depends on the percent D2O the cells are grown in aromatic side-chains will be highly protonated if 1H-glucose is used exchange labile N2H to N1H by temperature increase or chemical denaturation of the protein
 level of 2H labeling depends on the. percent D2O the cells are grown in. aromatic side-chains will be highly. protonated if 1H-glucose is used. exchange labile N2H to N1H by. temperature increase or chemical. denaturation of the protein.")
108
Metabolic pathway for generating 1H-methyl-Ile
Introduction to Isotope Labeling of Proteins For NMR 2H Labeling of the protein As we have seen, deuterium labeling a protein removes a majority of protons necessary for protein structure calculation can introduce site specific protonation to regain some proton based distance constraints label the methyl groups of Leu, Ile, and Val by adding to the growth media. use 1H-glucose to generate 1H-aromatic side-chains [3,3-2H2]-13C 2-ketobutyrate. [2,3-2H2]-15N, 13C Val Metabolic pathway for generating 1H-methyl-Ile

109
EXPERIMENTAL

110
The NMR spectrometer Magnet Probe Coils Transmitters
Amplifiers and pre-amplifiers Receiver ADC

111
The Magnet History First magnets were built using
ferromagnetic material= permanent magnet Then Electromagnets: i.e. field was generated by wiring of conducting material Now: cyomagnets: i.e. electromagnets made of superconducting wire. A “cutted” magnet

112
Cryomagnets Superconducting wire
has a resistance approximately equal to zero when it is cooled to a temperature close to absolute zero ( o C or 0 K) by emersing it in liquid helium. Once current is caused to flow in the coil it will continue to flow for as long as the coil is kept at liquid helium temperatures. The length of superconducting wire in the magnet is typically several miles.
 by. emersing it in liquid helium. Once current is. caused to flow in the coil it will continue to. flow for as long as the coil is kept at liquid. helium temperatures. The length of superconducting wire in the. magnet is typically several miles.")
113
The NMR spectrometer

114
Det. ADC w0 (reference) NMR Signal Computer Memory 500 MHz ± 2500 Hz
 NMR Signal Computer Memory 500 MHz ± 2500 Hz")
115
The Probe The sample probe is the name given to that part of the spectrometer which accepts the sample, sends RF energy into the sample, and detects the signal emanating from the sample. It contains the RF coil, sample spinner, temperature controlling circuitry, and gradient coils. Picture an axial cross section of a cylindrical tube containing sample. In a very homogeneous Bo magnetic field this sample will yield a narrow spectrum

116
B0 homogeneity In a more inhomogeneous field the sample will yield a broader spectrum due to the presence of lines from the parts of the sample experiencing different Bo magnetic fields.

117
Set up an experiment. What to do?
Shimming the magnet Lock Tune 90° Pulse

118
The NMR experiment: what do we need?
1. The magnet: Requires control of field homogeneity SHIM Requires stabilization of main field LOCK SHIM: additional coils with special field distribution, e.g. Z, Z2, Z3, X, Y, X3.... We have cryo shims and room temperature shims LOCK 1.contineously determine frequency of 2H signal of the solvent (deuterated solvents) 2. add a small extra field to the main field of the magnet to keep the overall field constant 3. 2H signal also used for shimming
 2. add a small extra field to the main field of the magnet. to keep the overall field constant. 3. 2H signal also used for shimming.")
119
Problem: how to keep B0 constant throughout the NMR sample?

120
B0 homogeneity In a more inhomogeneous field the sample will yield a broader spectrum due to the presence of lines from the parts of the sample experiencing different Bo magnetic fields.

121
The effect of shim coils
of z4 The effect of z1 The effect of z3 The effect of z2

122
SHIMMING line shape distortions from on-axis shims: Z1 OK Z2 Z3 Z4

123
Effect of B0 inhomogeneity in the NMR spectrum
AFTER BEFORE

124
Problem: how to keep B0 constant during an experiment?

125
The NMR experiment: what do we need?
1. The magnet: Requires control of field homogeneity SHIM Requires stabilization of main field LOCK SHIM: additional coils with special field distribution, e.g. Z, Z2, Z3, X, Y, X3.... We have cryo shims and room temperature shims LOCK 1.contineously determine frequency of 2H signal of the solvent (deuterated solvents) 2. add a small extra field to the main field of the magnet to keep the overall field constant 3. 2H signal also used for shimming
 2. add a small extra field to the main field of the magnet. to keep the overall field constant. 3. 2H signal also used for shimming.")
126
The Lock: How does it work?
The lock channel can be understood as a ‚complete indepenant spectrometer within the spectrometer‘: The resonance condition of NMR: w=g Bo but: Bo is not stable w=g (Bo+Ho) (Bo+Ho) = const. Ho Shim system Regulator Probe amplitude, frequency Transmitter 2H Receiver 2H
 (Bo+Ho) = const. Ho. Shim system. Regulator. Probe. amplitude, frequency. Transmitter 2H. Receiver 2H.")
127
Problem: how to optimized the sensitivity of the receiving coil with respect to the observed frequency?

128
Tuning

129
The tuning circuit

130
Problem: how to give a 90° pulse in real life?

131
Precession in the laboratory frame
dM/dt=M^B dM/dt=M^(B-w0/g) If B = B 0 dM/dt=0 R.F. at freq. w0 L.F. If B = B 0+B1 dM/dt=M^(B0 +B1 -w0/g) Rotation! dM/dt=M^B1 B1 Pulse: q=w1t=gB1t Pulse: q=p/2=gB1t
 If B = B 0. dM/dt=0. R.F. at freq. w0. L.F. If B = B 0+B1. dM/dt=M^(B0 +B1 -w0/g) Rotation! dM/dt=M^B1. B1. Pulse: q=w1t=gB1t. Pulse: q=p/2=gB1t.")
132
The 90° pulse Calibration of pulse lenght

133
Performing an NMR experiment
relax. FT Preparation Detection z t2 y w0 x The practical application of the rotating frame of reference….

134
A B C Static: w0 = wB z z z t2 y y y x x x Rotating (w0 = wB): t2 x’
w 108 Hz Rotating (w0 = wB): t2 x’ y’ z w 103 Hz
: t2. x’ y’ z. w 103 Hz.")
135
Det. ADC w0 (reference) NMR Signal Computer Memory 500 MHz ± 2500 Hz
 NMR Signal Computer Memory 500 MHz ± 2500 Hz")
136
AQ = DW·TD Acquisition Time
DE P1 = 1/BW AQ = DW·TD Acquisition Time PL1 D1 RG Repetition Time

137
My y y t x x x y t Mx

138
Quadrature Phase Detection

140
Pulse! -y -y The rotation of magnetization under the effect of 90° pulses according to the convention of Ernst et al.. -y -y -y -y -y -y -y y

141
The phase of an NMR signal

142
Phase Correction

143
AQ = DW·TD Acquisition Time
DE AQ = DW·TD Acquisition Time FT

144
Digital resolution Resolution is expressed in Hertz/point

145
Quadrature Phase Detection
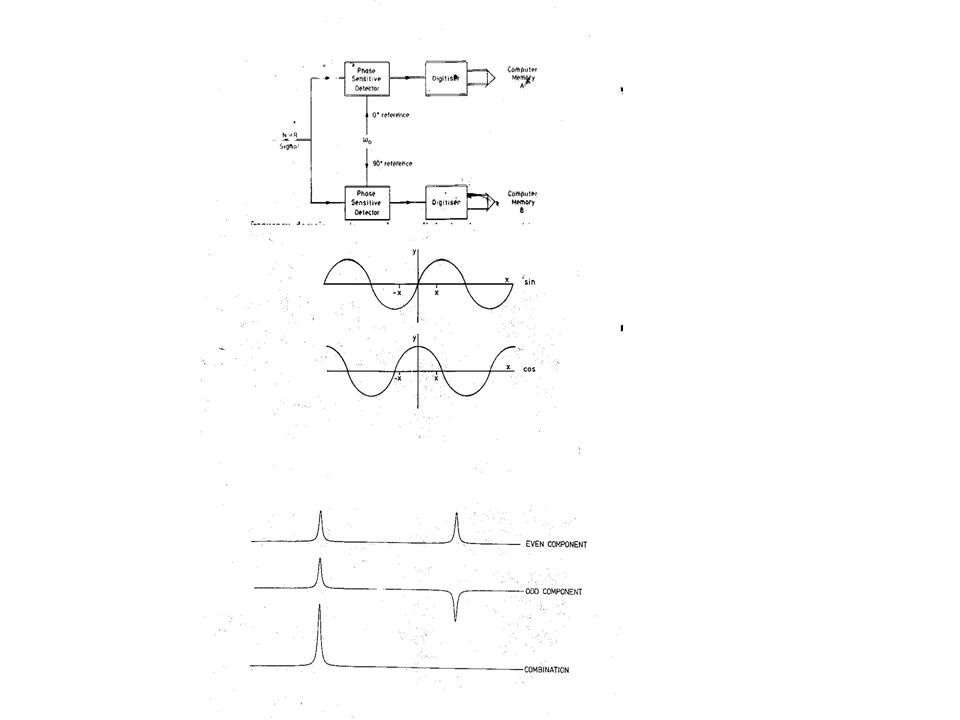
Computer Memory A PSD ADC 0° reference NMR Signal w0 90° reference Computer Memory B PSD ADC

146
Fourier Pairs

147
1D-NMR with/without removal of water

148
Free Induction Decay (FID)
Observed NMR signal in the time domain Resonance frequencies are acquired as a function of time t t t Common case of observed FIDs

149
Sensibilità dell’Esperimento NMR
S/N N g5/2 B03/2 N = Numero di spins che contribuiscono al segnale g rapporto giromagnetico del nuclide studiato B Camp magnetico utlizizzato

150
Signal to noise

151
Signal to noise Scans S/N

152
DE P1 = 1/BW AQ = DW·TD Acquisition Time PL1 D1 RG Repetition Time

153
Pulses and Phases