Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
amminoacidi

2
L’alanina è un a -amminoacido
acido 2(S)-ammino propanoico L’alanina è un a -amminoacido
-ammino propanoico. L’alanina è un a -amminoacido.")
3
La b-alanina è un b -amminoacido

4
Il GABA è un g -amminoacido
acido 4-ammino butanoico Il GABA è un g -amminoacido

5
Gli amminoacidi che si ottengono dall'idrolisi delle proteine sono tutti a-amminoacidi
H N C H 2 R

6
Gli a-amminoacidi naturali appartengono tutti alla serie sterica L.
(ad eccezione della glicina, che non ha atomi di carbonio asimmetrici)
")
7
H O O C N C H H H R - + Allo stato cristallino gli amminoacidi sono
nella forma zwitterionica - H O O C + N C H H H R

8
C O O H H N C H R 2 Gli amminoacidi naturali differiscono
Gruppo funzionale H N C H 2 R Gli amminoacidi naturali differiscono per la struttura della catena laterale

9
COO- G A V L I P Gly Ala Val Leu Ile Pro + H3N C H R R apolare

10
H COO- C H3N + Glicina Gly G

11
CH3 COO- C H H3N + Alanina Ala A

12
CH2 COO- C H H3N + CH CH3 H3C Leucina Leu L

13
COO- + H3N C H H C CH3 CH2 CH3 Isoleucina Ile I

14
COO- C H H3N + CH CH3 H3C Valina Val V

15
H N CH COO- CH2 H2C + Prolina Pro P

16
R Ser Thr Cys Met Asn Gln S T C M N Q COO- H3N C H R polare non carica
+ H3N C H R R polare non carica

17
COO- + H3N C H CH2OH Serina Ser S

18
C CH3 H OH COO- H3N + Treonina Thr T

19
COO- + H3N C H CH2 SH Cisteina Cys C

20
CH2 S CH3 COO- C H H3N + Metionina Met M

21
COO- C H H3N + CH2 O H2N Asparagina Asn N

22
CH2 COO- C H H3N + O H2N Glutammina Gln Q

23
COO- + H3N C H F Y W Phe Tyr Trp R R aromatica

24
COO- C H H3N + CH2 Fenilalanina Phe F

25
OH COO- C H H3N + CH2 Tirosina Tyr Y

26
COO- C H H3N + N CH2 Triptofano Trp W

27
COO- + H3N C H D E Asp Glu R R con carica negativa

28
CH2 COO- C H H3N + Ac. aspartico Asp D

29
CH2 COO- C H H3N + Ac. glutammico Glu E

30
R carica positivamente
COO- + H3N C H K H R Lys His Arg R R carica positivamente

31
CH2 NH3 + COO- C H H3N Lisina Lys K

32
COO- C H H3N + N CH2 Istidina His H

33
CH2 NH C NH2 + COO- H H3N Arginina Arg R

34
Alcuni amminoacidi sono essenziali
“o indispensabili”

35
indispensabili H N CH COO- CH2 H2C + glicina [G] (Gly) H COO- C H3N +
alanina [A] (Ala) CH3 COO- C H H3N + prolina [P] (Pro) indispensabili leucina [L] (Leu) CH2 COO- C H H3N + CH CH3 H3C isoleucina [I] (Ile) C CH3 CH2 H COO- H3N + valina [V] (Val) COO- C H H3N + CH CH3 H3C
![indispensabili H N CH COO- CH2 H2C + glicina [G] (Gly) H COO- C H3N +](http://slideplayer.it/slide/546700/1/images/35/indispensabili+H+N+CH+COO-+CH2+H2C+%2B+glicina+%5BG%5D+%28Gly%29+H+COO-+C+H3N+%2B.jpg "alanina [A] (Ala) CH3. COO- C. H. H3N. + prolina [P] (Pro) indispensabili. leucina [L] (Leu) CH2. COO- C. H. H3N. + CH. CH3. H3C. isoleucina [I] (Ile) C. CH3. CH2. H. COO- H3N. + valina [V] (Val) COO- C. H. H3N. + CH. CH3. H3C.")
36
indispensabili asparagiNa [N] (Asn) COO- C H H3N + CH2 O H2N CH2
(Q)lutammina [Q] (Gln) COO- C H H3N + O H2N cisteina [C] (Cys) SH CH2 COO- C H H3N + serina [S] (Ser) CH2OH COO- C H H3N + COO- C H H3N + COO- C H H3N + CH2 H C OH CH2 indispensabili CH3 Treonina [T ] (Thr) S CH3 Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli Metionina [M] (Met)
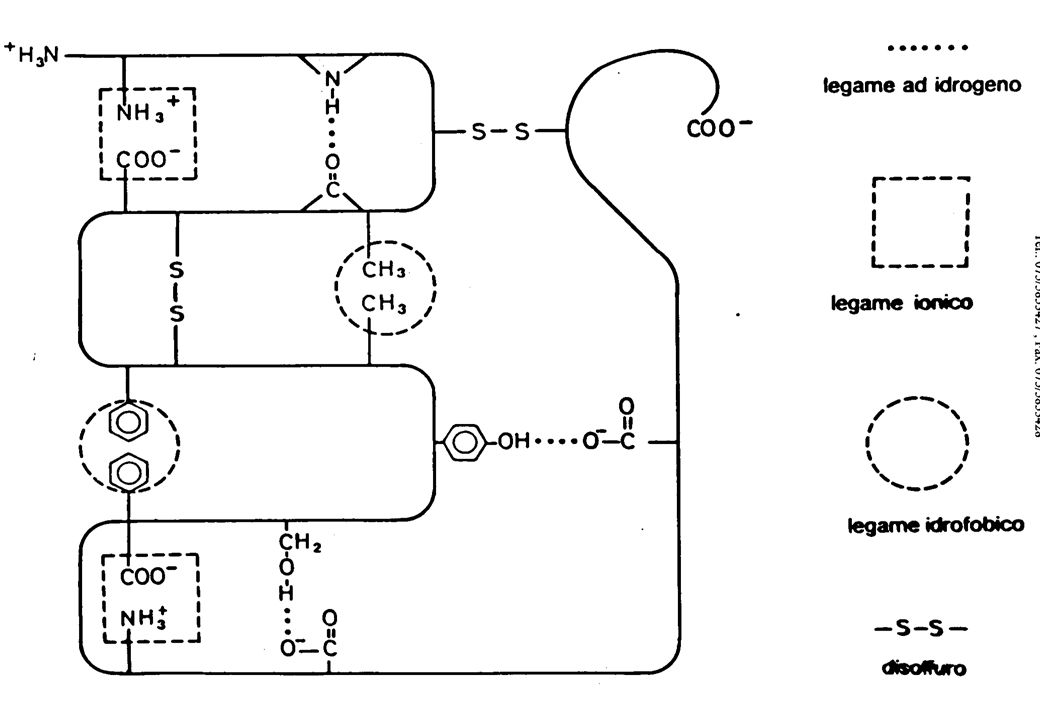
![indispensabili asparagiNa [N] (Asn) COO- C H H3N + CH2 O H2N CH2](http://slideplayer.it/slide/546700/1/images/36/indispensabili+asparagiNa+%5BN%5D+%28Asn%29+COO-+C+H+H3N+%2B+CH2+O+H2N+CH2.jpg "(Q)lutammina [Q] (Gln) COO- C. H. H3N. + O. H2N. cisteina [C] (Cys) SH. CH2. COO- C. H. H3N. + serina [S] (Ser) CH2OH. COO- C. H. H3N. + COO- C. H. H3N. + COO- C. H. H3N. + CH2. H. C. OH. CH2. indispensabili. CH3. Treonina [T ] (Thr) S. CH3. Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli. Metionina [M] (Met)")
37
indispensabili COO- C H H3N + N CH2 COO- C H H3N + CH2 OH COO- C H H3N
Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli indispensabili COO- C H H3N + N CH2 COO- C H H3N + CH2 OH COO- C H H3N + CH2 Tripto(W)fano [W] (Trp) Fenilalanina [F] (Phe) tYrosina [Y] (Tyr)
fano [W] (Trp) Fenilalanina [F] (Phe) tYrosina [Y] (Tyr)")
38
acido glu(E)tammico [E]
COO- C H H3N + CH2 COO- C H H3N + CH2 COO- acido aspar(D)ico [D] (Asp) acido glu(E)tammico [E] (Glu) Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli
![acido glu(E)tammico [E]](http://slideplayer.it/slide/546700/1/images/38/acido+glu%28E%29tammico+%5BE%5D.jpg "COO- C. H. H3N. + CH2. COO- C. H. H3N. + CH2. COO- acido aspar(D)ico [D] (Asp) acido glu(E)tammico [E] (Glu) Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli.")
39
Tutti indispensabili CH2 NH3 + COO- C H H3N aRginina [R] (Arg) CH2 NH
Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli Tutti indispensabili CH2 NH3 + COO- C H H3N aRginina [R] (Arg) CH2 NH C NH2 + COO- H H3N COO- C H H3N + N CH2 (K)lisina [K] (Lys) (H)istidina [H] (His)
![Tutti indispensabili CH2 NH3 + COO- C H H3N aRginina [R] (Arg) CH2 NH](http://slideplayer.it/slide/546700/1/images/39/Tutti+indispensabili+CH2+NH3+%2B+COO-+C+H+H3N+aRginina+%5BR%5D+%28Arg%29+CH2+NH.jpg "Attenzione!!!: la corruzione del nome di alcuni AA è solo un trucco mnemonico utile a ricordarne i simboli. Tutti indispensabili. CH2. NH3. + COO- C. H. H3N. aRginina [R] (Arg) CH2. NH. C. NH2. + COO- H. H3N. COO- C. H. H3N. + N. CH2. (K)lisina [K] (Lys) (H)istidina [H] (His)")
40
Caratteristiche chimiche
degli amminoacidi

41
PLP -amminoacido aldimmina HC CH2OPO3H2 HO H3C - H2O + H2O C O H N
COOH C H R NH2 aldimmina HC COOH C H R N + CH2OPO3H2 HO H3C - H2O + H2O + PLP C O H N CH2OPO3H2 HO H3C

42
chetimmina aldimmina H HC N CH2OPO3H2 HO H3C HC HO CH2OPO3H2 H3C N H
+ N CH2OPO3H2 HO H3C COOH C R COOH R C H N HC HO CH2OPO3H2 + H3C N H aldimmina

43
-chetoacido chetimmina + piridossammina-fosfato HC HO CH2OPO3H2 CH2
COOH C R O + H CH2 N CH2OPO3H2 HO H3C NH2 piridossammina-fosfato COOH R C N + H2O HC H - H2O HO CH2OPO3H2 + H3C N H chetimmina

44
-amminoacido1 + -chetoacido2
-chetoacido1 + -amminoacido2 Transamminasi PLP-dipendente

45
pigmento violaceo COOH C H R H2N ninidrina amminoacido ninidrina CO2 R
3 H2O

46
cisteina cistina ossidazione (-2H) riduzione (+2H) SH CH2 COO- C H H3N
 riduzione (+2H) SH CH2 COO- C H H3N")
47
H O O C N C H H H R - + Allo stato cristallino gli amminoacidi sono
nella forma zwitterionica (anfionica) - H O O C + N C H H H R
 - H. O. O. C. + N. C. H. H. H. R.")
48
gli amminoacidi sono anfoliti
forma anionica forma anfionica forma cationica

49
possono essere considerati acidi diprotici
Gli amminoacidi* possono essere considerati acidi diprotici Ciascuno dei due equilibri comporta la possibilità che l’amminoacido funzioni da tampone *monocarbossilici e monoamminici

50
Il pKa1 degli amminoacidi è influenzato dal gruppo amminico
glicina (ac. amminoacetico) pKa1 = 2.34 acido acetico pKa = 4.75
 pKa1 = acido acetico. pKa =")
51
pH OH- (equivalenti) Ka1 COO- C H H3N + Ka2 COO- C H H2N COOH C H H3N
7 12 pH + NaOH pK2 = 9.60 + NaOH Punto isoelettrico pKa1 = 2.34 0.5 1 1.5 2 OH- (equivalenti)
")
52
Al punto isoelettrico [AH2 ] = [A-]
+ AH2+ A- AH =
![Al punto isoelettrico [AH2 ] = [A-]](http://slideplayer.it/slide/546700/1/images/52/Al+punto+isoelettrico+%5BAH2+%5D+%3D+%5BA-%5D.jpg "+ AH2+ A- AH. =")
53
. . AH2+ AH + H + Ka1 Ka1 = [AH] [H+] [AH2+] A- AH + H + Ka2 Ka2 =
Per calcolare a quale valore di pH si abbia il punto isoelettrico dell’amminoacido si può ricorrere ad un “artificio matematico”
![. . AH2+ AH + H + Ka1 Ka1 = [AH] [H+] [AH2+] A- AH + H + Ka2 Ka2 =](http://slideplayer.it/slide/546700/1/images/53/.+.+AH2%2B+AH+%2B+H+%2B+Ka1+Ka1+%3D+%5BAH%5D+%5BH%2B%5D+%5BAH2%2B%5D+A-+AH+%2B+H+%2B+Ka2+Ka2+%3D.jpg "Per calcolare a quale valore di pH si abbia il punto isoelettrico. dell’amminoacido si può ricorrere ad un artificio matematico")
54
. . . . [AH] [H+] [AH2+] [A- ] [H+] [AH] K1·K2 = · K1 = [AH] [H+]
Dal prodotto delle due equazioni si ottiene la seguente : [AH] [H+] [AH2+] . [A- ] [H+] [AH] . K1·K2 = Elidendo i termini che compaiono sia al numeratore che al denominatore ... K1 = [AH] [H+] [AH2+] . K2 = [A- ] [H+] [AH] .
![[AH] [H+] [AH2+] [A- ] [H+] [AH] K1·K2 = · K1 = [AH] [H+]](http://slideplayer.it/slide/546700/1/images/54/%5BAH%5D+%5BH%2B%5D+%5BAH2%2B%5D+%5BA-+%5D+%5BH%2B%5D+%5BAH%5D+K1%C2%B7K2+%3D+%C2%B7+K1+%3D+%5BAH%5D+%5BH%2B%5D.jpg "Dal prodotto delle due equazioni si ottiene. la seguente : [AH] [H+] [AH2+] . [A- ] [H+] [AH] . K1·K2. = Elidendo i termini che compaiono sia al numeratore che al denominatore ... K1. = [AH] [H+] [AH2+] . K2. = [A- ] [H+] [AH] .")
55
. . pK1 + pK2 pI = 2 = [AH2+] [A-] [AH] [H+] [AH2+] [A- ] [H+] [AH]
E considerando che al punto isoelettrico = [AH2+] [A-] [AH] [H+] [AH2+] . [A- ] [H+] [AH] . K1·K2 = [H3O+]2 = K1 · K2 deriva che : pK1 + pK2 pI = 2 E quindi:
![. . pK1 + pK2 pI = 2 = [AH2+] [A-] [AH] [H+] [AH2+] [A- ] [H+] [AH]](http://slideplayer.it/slide/546700/1/images/55/.+.+pK1+%2B+pK2+pI+%3D+2+%3D+%5BAH2%2B%5D+%5BA-%5D+%5BAH%5D+%5BH%2B%5D+%5BAH2%2B%5D+%5BA-+%5D+%5BH%2B%5D+%5BAH%5D.jpg "E considerando che al punto isoelettrico. = [AH2+] [A-] [AH] [H+] [AH2+] . [A- ] [H+] [AH] . K1·K2. = [H3O+]2 = K1 · K2. deriva che : pK1 + pK2. pI = 2. E quindi:")
56
pK1 pK2 pKR R non polare R aromatico R polare non carico R negativo
R positivo glicina alanina valina leucina isoleucina prolina fenilalanina tirosina triptofano serina treonina cisteina metionina asparagina glutammina aspartato glutammato lisina arginina istidina

57
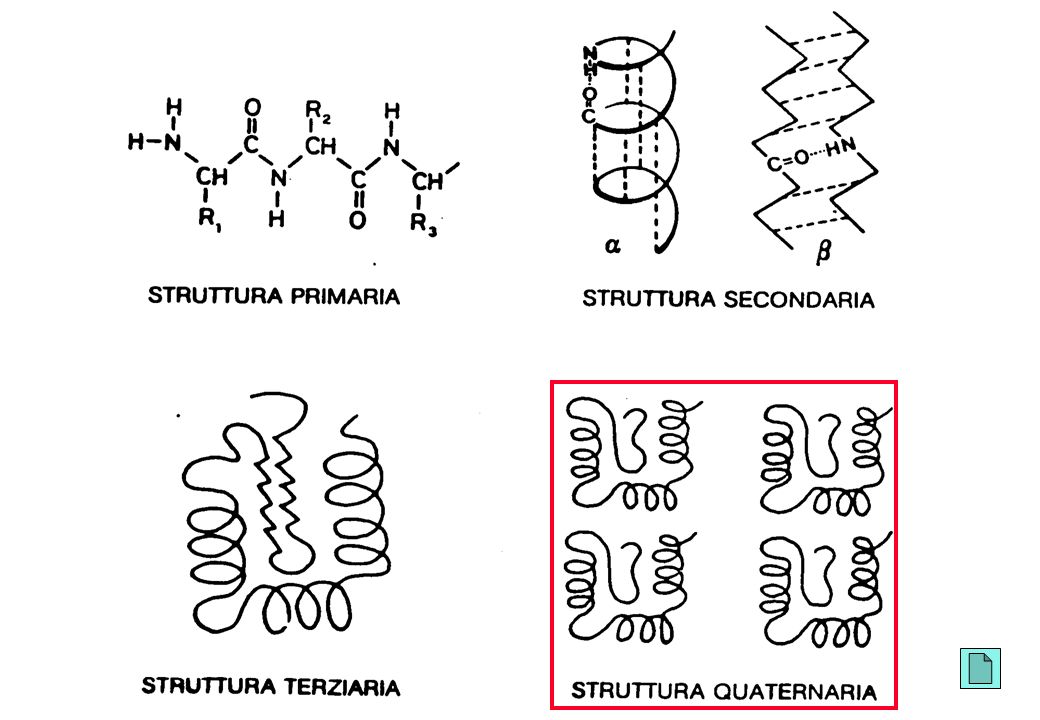
Gli amminoacidi si legano contraendo
legami peptidici

59
Mesomeria del gruppo funzionale peptidico Planarità del gruppo funzionale peptidico a a

60
seril-glicil-aspartil-alanil-leucina
SGDAL S G D A L amminoacido N-terminale amminoacido C-terminale

61
2,4-dinitro-fluorobenzene
identificazione dell'amminoacido N-terminale + HF 2,4-dinitro-fluorobenzene

62
+ gli altri AA idrolisi dinitrofenil-peptide dinitrofenil-AA

63
S G D A L SGDAL 24 combinazioni possibili degli altri 4 AA (n!)
GDAL / GDLA / GADL / GALD / GLDA / GLAD DALG / DAGL / DGAL / DGLA / DLAG / DLGA ADLG / ADGL / AGLD / AGDL / ALGD / ALDG LDAG / LDGA / LGDA / LGAD / LAGD / LADG

64
Determinazione della sequenza mediante “degradazione di Edman”
fenilisotiocianato idrolisi oligopeptide di n-1 AA (pronto per il ciclo successivo)
")
65
-SCALIQAFTYKESENQPCTGWDFSIMDFIFESGDAL
Nei polipeptidi, oltre al legame peptidico, esistono anche altre interazioni fra amminoacidi. Queste interazioni danno origine a strutture superiori

67
Ogni polipeptide allo stato nativo ha uno specifico folding, che è determinato dalla sua struttura primaria

68
Le quattro interazioni deboli implicate nel folding
di un polipeptide in ambiente acquoso attrazione repulsione Legame ionico ≈10 kcal/mole fra catene laterali 3-7 kcal/mole Legame H fra gruppi amidici 1-2 kcal/mole Interazione idrofobica < 1 kcal/mole Forze di van der Waals

69
Forze implicate nel folding della barnasi
Effetto idrofobico La ribonucleasi batterica barnasi, un piccolo polipeptide di 110 aminoacidi e 12.4 kDa di massa molecolare, è un buon modello per descrivere il folding di un polipeptide Phe7 Val10 Ala11 Leu14 Leu20 Tyr24 Ala74 Ile76 Ile88 Tyr90 Trp94 Ile96 Ile109

70
Forze implicate nel folding della barnasi Core idrofobico della barnasi
Phe56 Leu63 Trp71 Leu89 Tyr97 Tyr103 Phe106

71
Forze implicate nel folding della barnasi Legami idrogeno
Barnase – helix I Amphipathic Phe7-Asp-Gly-Val-Ala-Asp-Tyr-Leu-Gln-Thr-Tyr-His18

72
Forze implicate nel folding della barnasi Interazioni elettrostatiche
Arg69 Asp93 Rompendo questo legame ionico la proteina diventa unfolded

73
Forze implicate nel folding della barnasi Ponti disolfuro
Ser-Cys85 His-Cys102 mutante

74
Conseguenze della mesomeria del gruppo funzionale peptidico
Planarità e isomeria cis/trans Angoli diedri di torsione intorno ai legami e

75
R R Queste torsioni sono limitate dall’ingombro sterico (e da interazioni elettrostatiche). La piccola catena laterale della alanina impedisce le modificazioni conformazionali molto meno di quella, “più ingombrante”, del triptofano. La glicina gode della massima libertà conformazionale mentre la prolina, per la struttura ciclica in cui è compreso l’azoto del gruppo funzionale, ha una libertà di modificazioni conformazionali fortemente limitata.
. La piccola catena laterale della alanina impedisce le modificazioni conformazionali molto meno di quella, più ingombrante , del triptofano. La glicina gode della massima libertà conformazionale mentre la prolina, per la struttura ciclica in cui è compreso l’azoto del gruppo funzionale, ha una libertà di modificazioni conformazionali fortemente limitata.")
76
Il paradosso di Levinthal
Per un polipeptide di 101 amminoacidi : 8100 = 2x1090 conformazioni possibili. Con una frequenza di modificazione conformazionale = 1013 s-1 (diecimila milardi di volte al secondo) al polipeptide occorrerebbero poco meno di 1070 anni per assumere tutte le conformazioni possibili
 al polipeptide occorrerebbero poco meno di 1070 anni per assumere tutte le conformazioni possibili.")
77
Cooperatività The folding funnel G stato “folded”

78
Ogni polipeptide allo stato nativo ha uno specifico folding, che è determinato dalla sua struttura primaria

79
Ogni polipeptide allo stato nativo ha uno specifico folding, che è determinato dalla sua struttura primaria

80
C O C O Ca Ca H N Ca Ca Legame H della struttura secondaria H R N H N

81
a-elica Mioglobina & GAPDH

82
Struttura secondaria a “foglietto pieghettato”
b-sheet antiparallelo b-sheet parallelo

83
Effetti della struttura secondaria sulle caratteristiche
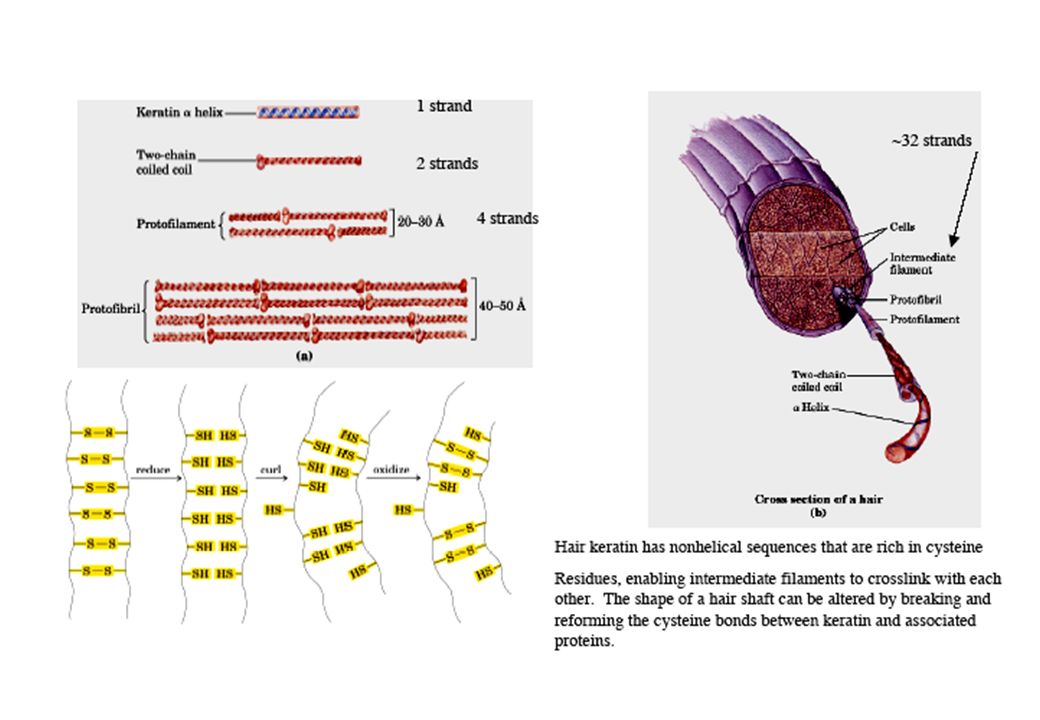
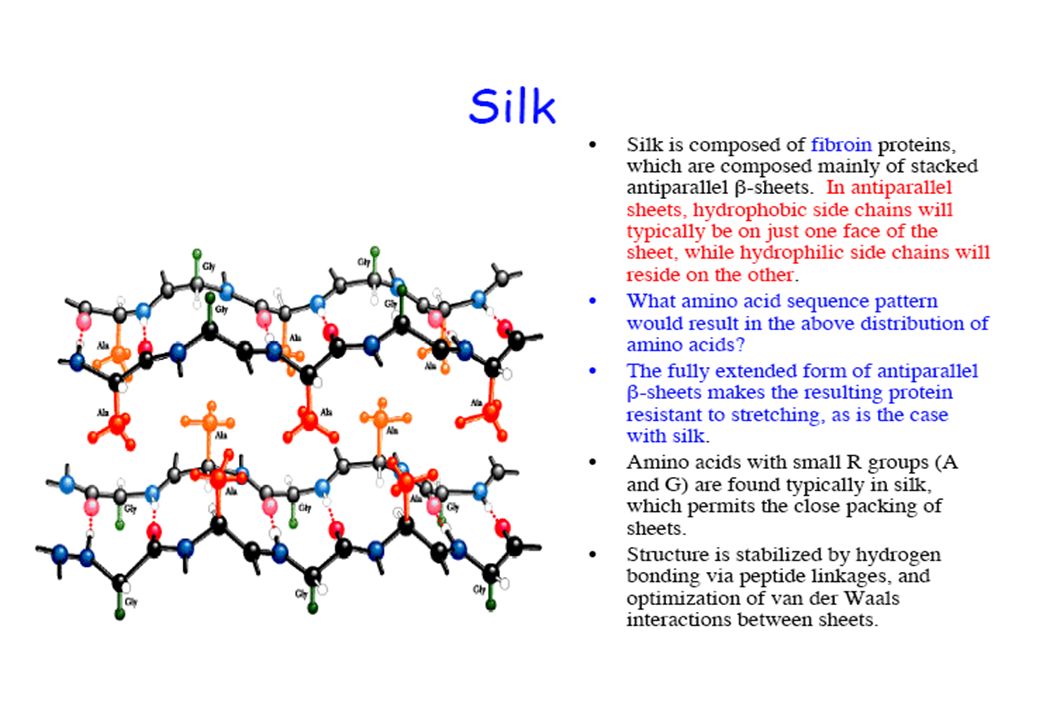
dei polipeptidi STRUTTURA CARATTERISTICHE ESEMPI a-elica con cross-linking di cistina 0,54 nm/giro dell’elica 3,6 residui AA/ giro dell’elica Catene laterali rivolte all’esterno a-cheratina (peli, unghie, piume) durezza e rigidità (variabili) b-sheet distanza fra i residui: 0,65-0,70 nm o 0,34 nm Catene laterali alternativamente sopra e sotto il piano Fibroina della seta Filamenti flessibili Tripla elica del collageno Coiled-coils determinati da legami non-covalenti (talora anche da ponti disolfuro) Collagene dei tendini, proteine della matrice ossea Alta resistenza alla tensione
 durezza e rigidità (variabili) b-sheet. distanza fra i residui: 0,65-0,70 nm o 0,34 nm. Catene laterali alternativamente sopra e sotto il piano. Fibroina della seta. Filamenti flessibili. Tripla elica del collageno. Coiled-coils determinati da legami non-covalenti (talora anche da ponti disolfuro) Collagene dei tendini, proteine della matrice ossea. Alta resistenza alla tensione.")
84
a-cheratina

87
Coiled-coils (collageno)
")
88
Si definiscono legami della struttura terziaria tutti i legami che si stabiliscono
fra catene laterali di amminoacidi costituenti la stessa catena polipeptidica

90
Gliceraldeide-3-fosfato-deidrogenasi
rasmol

92
Struttura quaternaria
dell’emoglobina

94
indice idropatico R non polare R aromatico prolina - 1.6 glicina - 0.4
R polare non carico R negativo R positivo prolina glicina alanina leucina valina isoleucina tirosina triptofano fenilalanina asparagina glutammina serina treonina metionina cisteina aspartato glutammato arginina lisina istidina - 1.6 - 0.4 1.8 3.8 4.2 4.5 - 1.3 - 0.9 2.8 - 3.5 - 0.7 - 3.2

96
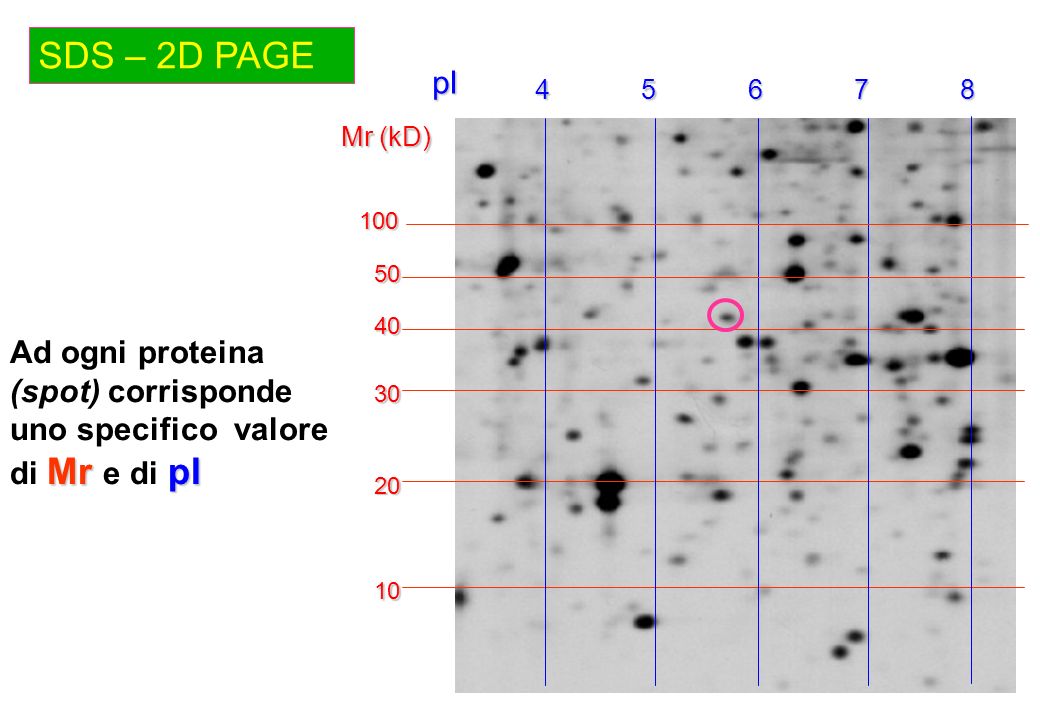
2D-PAGE

97
La soluzione proteica viene risolta nelle sue
componenti mediante focalizzazione isoelettrica su un gradiente di pH immobilizzato proteine acide proteine basiche + - pH 3 gradiente di pH immobilizzato pH 10 Ogni proteina migra nel campo elettrico fino a raggiungere la zona il cui pH (immobilizzato) corrisponde al suo punto isoelettrico
 corrisponde al suo punto isoelettrico.")
98
- Gel di agarosio Gel di poliacrilammide +

99
- +

100
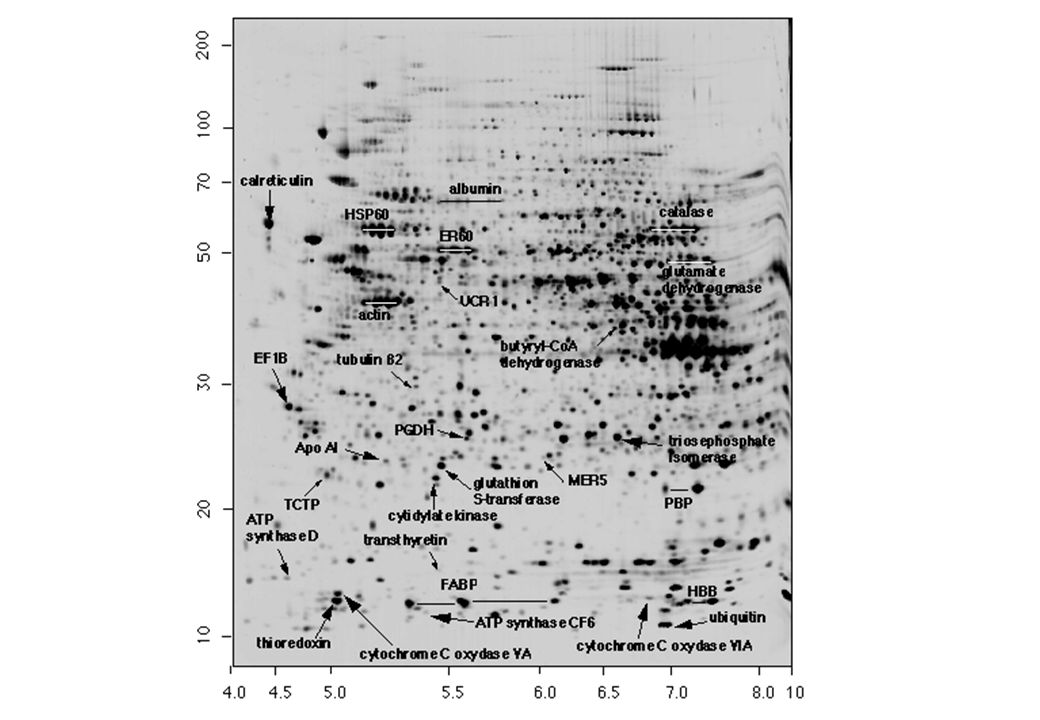
SDS – 2D PAGE 4 5 6 7 8 pI Mr (kD) 100 50 40 Ad ogni proteina (spot) corrisponde uno specifico valore di Mr e di pI 30 20 10
101
Una procedura semplice per l’analisi proteomica
“spot picking” estratto proteico coltura cellulare o tessuto elettroforesi bidimensionale digestione triptica ricerca in banca-dati identificazione Spettrometria di massa

103
eme mioglobina

Presentazioni simili


 Gruppo carbossilico Gruppo amminico>")

















>")