Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
CONTROLLO DEL METABOLISMO

2
≠ Completamente ossidato Muscolo scheletrico
Periodi di bassa richiesta di E Periodi di alta richiesta di Energia (dal glicogeno) ≠ Muscolo cardiaco Continua necessità di E per una contrazione regolare: glucosio completamente ossidato Completamente ossidato
 ≠ Muscolo cardiaco. Continua necessità di E per una contrazione regolare: glucosio completamente ossidato. Completamente ossidato.")
3
G1P + UTP UDPGlucosio (UDPG)
+ 2 NAD+ sintesi del glicogeno UDP-Glucuronato UDP-Glucuronato + R-OH R-O-Glucuronato + UDP Composti apolari coniugati con accettori non polari per formare composti più polari - composti endogeni: bilirubina, ormoni steroidei, T3 - composti esogeni: farmaci

4
LIVELLI DI CONTROLLO DEL METABOLISMO
IMMEDIATO non richiede energia - flusso del substrato (controllato da Km) - regolazione allosterica prodotto (inibizione a feed back) metaboliti H+ ; Ca+2 A BREVE TERMINE (MINUTI) - RICHIEDE ENERGIA modificazione covalente (fosforilazione - defosforilazione di proteine) A LUNGO TERMINE (ORE) - RICHIEDE ENERGIA Modificazione dei livelli proteici tramite - biosintesi proteica - degradazione proteica
 - regolazione allosterica. prodotto (inibizione a feed back) metaboliti. H+ ; Ca+2. A BREVE TERMINE (MINUTI) - RICHIEDE ENERGIA. modificazione covalente (fosforilazione - defosforilazione di proteine) A LUNGO TERMINE (ORE) - RICHIEDE ENERGIA. Modificazione dei livelli proteici tramite. - biosintesi proteica. - degradazione proteica.")
5
Controllo della glicolisi
A BREVE TERMINE - controllo allosterico - ciclo dei substrati A LUNGO TERMINE - modificazione covalente - modificazione dei livelli enzimatici

6
2 ADP + miochinasi ATP + AMP

7
Controllo allosterico
Fosfofruttochinasi ATP AMP Ca2+ CITRATO H F2,6bisP (insulina) F1,6bisP fosfatasi AMP F2,6BP Glicogeno fosforilasi ATP AMP Ca2+ G6P Glicogeno sintasi ATP G6P INIBIZIONE DA H* FUNZIONA DA SISTEMA DI PROTEZIONE
 F1,6bisP fosfatasi AMP F2,6BP. Glicogeno fosforilasi ATP AMP Ca2+ G6P. Glicogeno sintasi ATP. G6P. INIBIZIONE DA H* FUNZIONA DA SISTEMA DI PROTEZIONE.")
8
Controllo allosterico e Ciclo dei substrati Muscolo ATP/AMP 50
ATP/ADP 10 ATP 5 mM 10% 4,5 mM AMP 0,1 mM 600% ,6 mM L’aumento dell’AMP comporta un aumento di 10 volte dell’attività della PFK contemporaneamente calo di 10 volte della attività della fosfatasi RISULTATO: aumento 100 volte del flusso glicolitico

9
Meccanismo d’azione degli ormoni

10
SEGNALI CHIMICI EXTRACELLULARI
hanno un meccanismo generale comune CONTROLLO ORMONALE NEUROTRASMISSIONE OLFATTO GUSTO VISTA CRESCITA DIFFERENZIAMENTO

11
NATURA CHIMICA degli ORMONI
POLIPEPTIDICA insulina, glucagone, ormoni ipofisari paratormone AMMINOACIDICA (dalla tirosina) adrenalina, ormoni tiroidei (in blu) composti lipofili, STEROIDEA richiedono trasportatori ematici ormoni sessuali si legano a recettori intracellulari corticosurrenalici 1,25-diidrossi colecalciferolo o 1,25 (OH)2 D3
 adrenalina, ormoni tiroidei. (in blu) composti lipofili, STEROIDEA - richiedono trasportatori ematici. ormoni sessuali - si legano a recettori intracellulari. corticosurrenalici. 1,25-diidrossi colecalciferolo o 1,25 (OH)2 D3.")
12
I recettori per gli ormoni steroideI formano eterodimeri con
RXR recettore per l’acido retinoico (derivato Vit A) Extrac. citoplasma RXR Complesso coattivatore Trascrizione basale nucleo DNA
 Extrac. citoplasma. RXR. Complesso. coattivatore. Trascrizione. basale. nucleo. DNA.")
13
MECCANISMI DI TRASDUZIONE DEL SEGNALE DI ADRENALINA E DI ORMONI POLIPEPTIDICI
(Glucagone)
")
14
RECETTORE (membrana) legame 1:1 R-ormone II. TRASDUZIONE (membrana)
I. SEGNALE: ORMONE RECETTORE (membrana) legame 1:1 R-ormone II. TRASDUZIONE (membrana) amplificazione del segnale tramite enzimi proteine G, adenilato ciclasi, fosfolipasi C III. SECONDI MESSAGGERI (citoplasma, membrana) AMPc, Ca2+ , inositolo 1,4,5,trifosfato, diacilglicerolo IV. PROTEIN CHINASI; FOSFOPROTEIN FOSFATASI V. RISPOSTA CELLULARE attivazione enzimi, fattori di trascrizione, canali di membrana
 legame 1:1 R-ormone. II. TRASDUZIONE (membrana) amplificazione del segnale tramite enzimi. proteine G, adenilato ciclasi, fosfolipasi C. III. SECONDI MESSAGGERI (citoplasma, membrana) AMPc, Ca2+ , inositolo 1,4,5,trifosfato, diacilglicerolo. IV. PROTEIN CHINASI; FOSFOPROTEIN FOSFATASI. V. RISPOSTA CELLULARE. attivazione enzimi, fattori di trascrizione, canali di membrana.")
15
Recettore -adrenergico (R)
R + ormone R..ormone conseguente cambio conformazionale del recettore DISATTIVAZIONE (se permane il legame R..ormone) 1. la “chinasi del recettore -adrenergico” riconosce la forma attiva 2. il recettore viene fosforilato (R-P) 3. la proteina -arrestina lega il R-P 4. si interrompe l’interazione con le proteine G
 1. la chinasi del recettore -adrenergico riconosce la forma attiva. 2. il recettore viene fosforilato (R-P) 3. la proteina -arrestina lega il R-P. 4. si interrompe l’interazione con le proteine G.")
16
Subunità : lenta attività GTPasica (sec)
L’idrolisi del GTP funge da orologio incorporato che spontanemante riporta allo stato inattivo adenilato ciclasi inattiva adenilato ciclasi attiva La tossina colerica blocca nella forma attiva La tossina della pertosse inattiva il sistema

17
- PROTEIN CHINASI Ser/Thr, Tyr
Premio Nobel 1992 Dal genoma si calcola differenti protein chinasi - PROTEIN FOSFATASI

18
R C ATP C R 2 + 2 membrana cellulare cAMP AMP Protein chinasi A
PKA (C2R2 ) fosforila residui di Ser Glucagone Adrenalina Paratormone ACTH, LH, FSH R C membrana cellulare ATP + 4 cAMP 2 R -cAMP + 2 C adenilato ciclasi attiva cAMP + ATP Fosfodiesterasi proteina fosfoproteina inibita da caffeina teofillina AMP fosfatasi EFFETTI FISIOLOGICI
 fosforila residui di Ser. Glucagone. Adrenalina. Paratormone. ACTH, LH, FSH. R. C. membrana. cellulare. ATP. + 4 cAMP. 2. R. -cAMP C. adenilato. ciclasi attiva. cAMP. + ATP. Fosfodiesterasi. proteina fosfoproteina. inibita da. caffeina. teofillina. AMP. fosfatasi. EFFETTI. FISIOLOGICI.")
19
REIDRATAZIONE CON SALI E GLUCOSIO
A1 + A2 A Tossina colerica 5 subunita B B B si lega alla membrana della mucosa intestinale A entra all’interno della cellula e blocca proteine G nella forma attiva catalizza la ADP ribosilazione delle proteine G Subunità -Arg-Ribosio -P-P Ribosio - Adenina (ADPribosio) AMPc 100 volte più elevato PKA apertura canali per il Cl- ed eccessiva perdita di NaCl e H2O Diarrea con perdita di 1 litro/h acqua ricca di sali REIDRATAZIONE CON SALI E GLUCOSIO
 AMPc 100 volte più elevato. PKA. apertura canali per il Cl- ed eccessiva perdita di NaCl e H2O. Diarrea con perdita di 1 litro/h acqua ricca di sali. REIDRATAZIONE CON SALI E GLUCOSIO.")
21
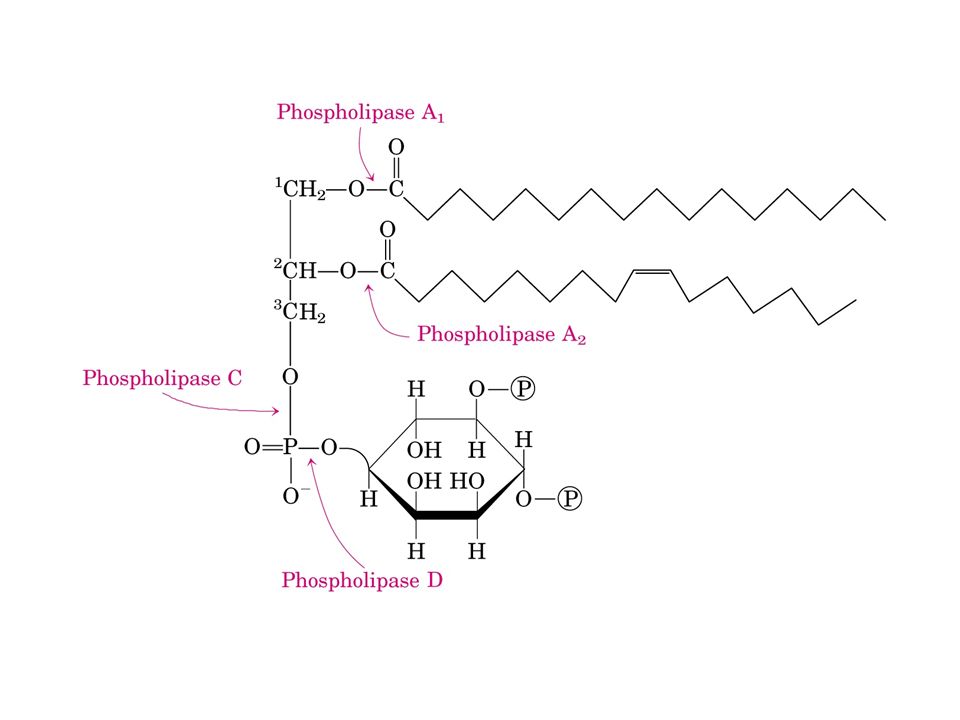
Fosfatidil inositolo 4,5 bisfosfato (PIP2)
acetilcolina, vasopressina, ossitocina, neurotrasmettitori membrana Fosfatidil inositolo 4,5 bisfosfato (PIP2) FOSFOLIPASI C diacilglicerolo (DAG) (apolare) regolatore di PKC- Ca2+ fosforila Ser/Thr inositolo 1,4,5,trisfosfato (IP3) (polare idrosolubile) Rilascio di Ca2+ dal R.E. Protein chinasi C (PKC) forma solubile PKC- Ca2+ trasloca sulla membrana secondi messaggeri sinergici
 FOSFOLIPASI C. diacilglicerolo (DAG) (apolare) regolatore di PKC- Ca2+ fosforila Ser/Thr. inositolo 1,4,5,trisfosfato (IP3) (polare idrosolubile) Rilascio di Ca2+ dal R.E. Protein chinasi C (PKC) forma solubile. PKC- Ca2+ trasloca sulla membrana. secondi messaggeri sinergici.")
22
Recettore dell’insulina
famiglia delle tirosinchinasi Tetramero 22 alfa extracellulare: sito di legame dell’insulina beta intracellulare: attività chinasica I. Legame dell’insulina attiva autofosforilazione di residui di Tyr II. La forma fosforilata ha attività chinasica verso altre proteine intrac

23
IRS-1 (substrato 1 del recettore dell’ insulina)
P p membrana IRS-1 (substrato 1 del recettore dell’ insulina) la sua fosforilazione induce effetti mitogeni, espressione genica GLUT-4 biosintesi proteine trasporto glucosio muscolo, tessuto adiposo biosintesi glicogeno biosintesi acidi grassi
 la sua fosforilazione induce. effetti mitogeni, espressione genica. GLUT-4. biosintesi. proteine. trasporto glucosio muscolo, tessuto adiposo. biosintesi. glicogeno. biosintesi. acidi grassi.")
24
Insulina stimola la fosfodiesterasi con calo livelli di AMPciclico stimola fosfoprotein fosfatasi

25
IPOGLICEMIA GLUCAGONE
Glicogenolisi attivata fosforilasi, inibita glicogeno sintasi Gluconeogenesi attivata fruttosio 1,6 bisfosfatasi inibita fosfofruttochinasi IPERGLICEMIA INSULINA Importo glucosio (GLUT 4) Glicogenolisi inibita fosforilasi, attivata glicogeno sintasi Glicolisi
 Glicogenolisi. inibita fosforilasi, attivata glicogeno sintasi. Glicolisi.")
26
protein chinasi A (PKA)
GLUCAGONE, ADRENALINA adenilato ciclasi cAMP protein chinasi A (PKA) GLICOGENO SINTASI-P (inattiva) fosforilasi chinasi ()4 subunità catalitica siti di fosforilazione calmodulina (lega Ca2+) FOSFORILASI b inattiva FOSFORILASI a attiva PROTEIN FOSFATASI -P (inattiva)
 GLICOGENO. SINTASI-P. (inattiva) fosforilasi chinasi ()4. subunità catalitica. siti di fosforilazione. calmodulina (lega Ca2+) FOSFORILASI b inattiva. FOSFORILASI a attiva. PROTEIN. FOSFATASI -P. (inattiva)")
27
controllo covalente ormonale controllo allosterico immediato
Fosforilasi chinasi 2 ATP 2 ADP Fosfoprotein fosfatasi Forma R attiva Forma T poco AMP ATP G6P -P P- Fosforilasi a Fosforilasi b controllo covalente ormonale non soggetto a regolazione allosterica ATP/AMP controllo allosterico immediato dipende da carica energetica regolazione allosterica scavalcata da quella ormonale se è richiesta risposta prolungata

28
Insulina induce defosforilazione
attiva - PROTEIN FOSFATASI - GLICOGENO SINTASI forma defosforilata attiva denominata: Forma I indipendente da regolazione allosterica Viceversa Glicogeno sintasi poco attiva nella forma fosforilata denominata: Forma D dipendente da regolazione allosterica

29
ATP + F6P F2,6 bisP + H2O Controllo ormonale: Gluconeogenesi epatica
- Fosfofruttochinasi-2 (PFK-2) - Fruttosio 2,6bisfosfatasi-2 (FBPasi-2) Domini diversi dello stesso enzima bifunzionale enzima defosforilato. dall’ insulina ATP + F6P F2,6 bisP + H2O fosfoenzima dal glucagone Attiva fosfofruttochinasi Inibisce fruttosio 1,6 bis fosfatasi
 - Fruttosio 2,6bisfosfatasi-2 (FBPasi-2) Domini diversi dello stesso enzima bifunzionale. enzima defosforilato. dall’ insulina. ATP + F6P F2,6 bisP + H2O. fosfoenzima dal glucagone. Attiva fosfofruttochinasi. Inibisce fruttosio 1,6 bis fosfatasi.")
30
in presenza di insulina
F6P + ATP + enzima defosforilato attivo F2,6 bisP + H2O attiva fosfofruttochinasi inattiva fruttosio 1,6 bis fosfatasi AUMENTA GLICOLISI e DIMINUISCE GLUCONEOGENESI In presenza di glucagone F6P + ATP fosfoenzima inattivo + F2,6 bisP + H2O inattiva fosfofruttochinasi attiva fruttosio 1,6 bis fosfatasi DIMINUISCE GLICOLISI e AUMENTA GLUCONEOGENESI AUMENTA GLICEMIA

31
Regolazione tramite l’aumento o la diminuizione dell’
espressione di geni che codificano per enzimi chiave. INSULINA Aumenta sintesi dell’enzima piruvato chinasi ed aumenta il flusso glicolitico (per dare precurosi per la sintesi degli acidi grassi). GLUCOCORTICOIDI Aumenta sintesi dell’enzima fosfoenolpiruvato carbossichinasi ed aumenta la gluconeogenesi.
. GLUCOCORTICOIDI. Aumenta sintesi dell’enzima fosfoenolpiruvato carbossichinasi ed aumenta la gluconeogenesi.")
Presentazioni simili


















 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")
