Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Elementi trasponibili
Sequenze che hanno la capacità di muoversi da un sito all’altro del cromosoma sito accettore sito donatore Diversamente dagli altri processi coinvolti nella ristrutturazione genomica, per la trasposizione non c’è nessuna omologia tra le sequenze del sito donatore e accettore

2
GATCA CTAGT sito bersaglio GATCA CTAGT GATCA CTAGT ripetizioni dirette del sito bersaglio

3
{ IS1 IS2, IS3, IS4, IS5 Gli elementi IS InsA InsB 768 bp
Le IS sono unità autonome e codificano solo per le proteine necessarie alla propria trasposizione. Sono costituenti comuni di cromosomi e plasmidi { IR 210 bp 273 bp IR E.coli Shigella Serratia IS1 InsA InsB 768 bp IR IR Trasposasi IS2, IS3, IS4, IS5 lunghezza media sequenze IS: bp lunghezza media IR: bp

4
Trasposoni composti altri geni IR IR IR IR Tn3 4957 Amp Tn501 8200 Hg
sequenza IS sequenza IS trasposone lunghezza modulo marcatori genetici (bp) terminale Tn Amp Tn Hg Tn Utilizzazione lattosio Tn IS50 Kc Tn IS1 Cm Tn IS10 Tet Tn IS1 Enterotossina stabile al calore
 terminale. Tn Amp. Tn Hg. Tn Utilizzazione lattosio. Tn IS50 Kc. Tn IS1 Cm. Tn IS10 Tet. Tn IS1 Enterotossina stabile. al calore.")
5
Origine dei trasposoni composti
trasposizione in un sito adiacente IS IS 1 o più geni batterici

6
Formazione di sequenze ripetute dirette nel sito bersaglio di un trasposone
GGGTTTCCAAA CCCAAAGGTTT La trasposasi induce tagli sfalsati nel sito bersaglio GGGTTTCCAA A C CCAAAGGTTT

7
Formazione di sequenze ripetute dirette nel sito bersaglio di un trasposone
inserzione del trasposone GGGTTTCCAA A C CCAAAGGTTT riempimento delle interruzioni G GGTTTCCAA GGTTTCCAA A C CCAAAGGTT CCAAAGGTT T

8
Cambiamenti genetici prodotti dai trasposoni
Inattivazione del gene bersaglio promotore DNA mRNA proteina

9
Cambiamenti genetici prodotti dai trasposoni
Inattivazione del gene bersaglio promotore DNA mRNA proteina assente o tronca

10
Nel caso di trasposizione in un operone si osserva un effetto “polare”
gene 1 gene 2 gene 3 mRNA policistronico P Anche i geni 2 e 3 non vengono espressi in quanto la trascrizione è bloccata dall’ elemento trasponibile

11
Cambiamenti genetici prodotti dai trasposoni
Attivazione di geni silenti promotore debole DNA gene non trascritto o scarsamente trascritto promotore forte gene altamente trascritto Elementi trasponibili hanno spesso promotori direzionati verso l’esterno

12
L’odissea evolutiva del gene ampC di Klebsiella
I trasposoni svolgono un ruolo importante nel movimento dei geni per la resistenza agli antibiotici e altri geni da un genoma a un plasmide e viceversa Il gene ampC di K. pneumoniae codifica per una b-lattamasi promotore debole ampC Il ceppo iniziale di Klebsiella era poco resistente ai beta-lattamici poichè il gene ampC era scarsamente trascritto

13
L’odissea evolutiva del gene ampC di Klebsiella
Sotto la selezione degli antibiotici il gene ampC ha acquisito due sequenze IS con il risultato di essere maggiormente trascritto conferendo un livello di resistenza clinicamente significativo promotore forte ampC IS IS Il risultante trasposone è stato capace di muovere ampC in un plasmide coniugativo, oggi diffuso a molti ceppi di Klebsiella Quello che un tempo era un gene di resistenza clinicamente irrilevante è diventato il terrore degli ospedali – tutto grazie alle IS S 111

14
Riarrangiamenti prodotti dai trasposoni
X delezione

15
Riarrangiamenti prodotti dai trasposoni
b c d b a X d c inversione a c b d

16
Riarrangiamenti prodotti dai trasposoni
fusione di repliconi formazione del cointegrato

17
il trasposone Tn3 sito di risoluzione del cointegrato tnpA tnpR bla
trasposasi resolvasi b-lattamasi sequenza ripetuta invertita di 38 bp

18
Un virus trasponibile: il batteriofago Mu
E’ un virus temperato In seguito ad infezione, il suo DNA si integra in siti distribuiti casualmente sul cromosoma Tra i batteri lisogeni si osservano mutanti dovuti all’inattivazione per integrazione del profago Si replica mediante trasposizione (una nuova copia del DNA virale è inserita in un punto diverso del cromosoma batterico) Come i trasposoni è in grado di promuovere riarrangiamenti del materiale genetico (delezioni, inversioni, fusione di repliconi ecc.)
 Come i trasposoni è in grado di promuovere riarrangiamenti del materiale genetico (delezioni, inversioni, fusione di repliconi ecc.)")
19
La mappa genetica del fago Mu
immunità geni della testa e della coda c A B C integrazione replicazione regione G invertibile

20
Il plasmide di resistenza R1

21
Gli elementi trasponibili sono responsabili dell’integrazione del fattore F
gd IS2 inc, rep oriT A L E B C F H G S D I J K 94 Kb

22
L’adattamento dei batteri
Strategie di adattamento mutazione regolazione dell’espressione genica regolazione della trascrizione trasferimento genico orizzontale regolazione della traduzione regolazione post-traduzionale

24
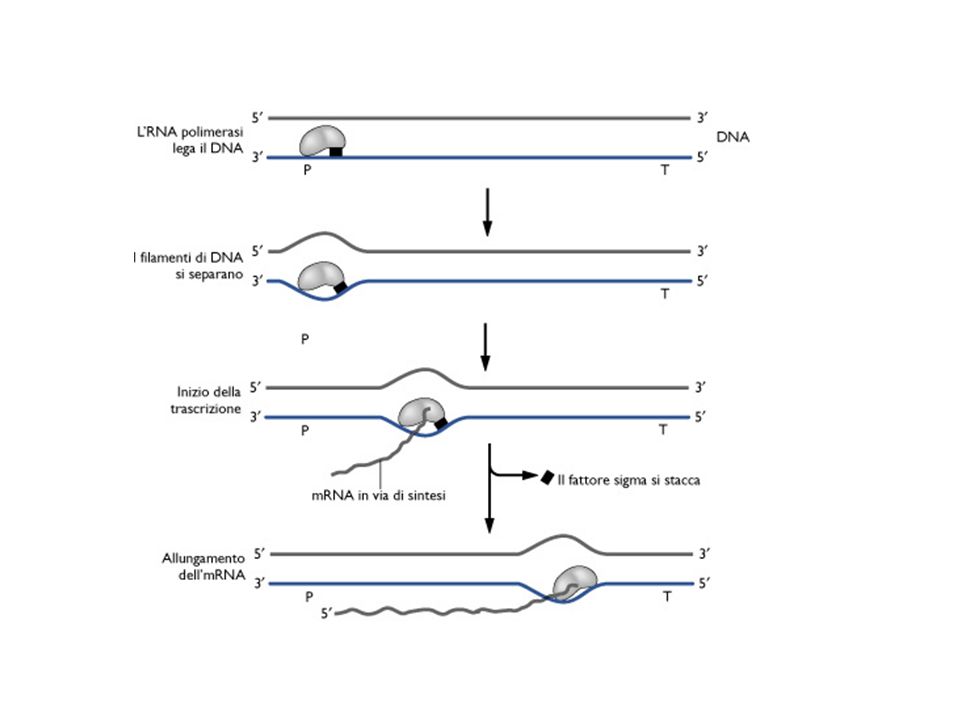
RNA polimerasi e inizio della trascrizione
regione -35 regione -10 T82T84G78A65C54A45 T80A95T45A60A50T96

25
s a2bb’s a2bb’ RNA polimerasi
Complesso proteico di D costituito da 5 subunità: Nucleo enzimatico a2bb’ Fattore sigma s Il nucleo enzimatico ha la capacità di sintetizzare RNA su uno stampo di DNA, ma non è in grado di iniziare la trascrizione al sito corretto. Il fattore sigma è responsabile del corretto riconoscimento del promotore. Viene rilasciato appena inizia la trascrizione.

26
Fattori sigma alternativi e regolazione globale dell’espressione genica
Subunità sigma Geni trascritti Segnale in E. coli s 70 maggior parte dei geni s 32 geni heat-shock temperatura elevata s 55 geni regolati dall’azoto carenza di ammonio s 38 geni da stress ossidativi agente ossidante Ogni fattore sigma riconosce una classe di promotori con specifiche sequenze consensus -35 e -10.

27
La sporulazione in Bacillus subtilis e i fattori sigma alternativi
La sporulazione rappresenta un drastico cambiamento dello stile di vita del batterio La transizione da una fase all’altra si realizza attraverso cambiamenti globali dell’espressione genica mediati da fattori sigma alternativi Subunità sigma Geni trascritti s 43 fase vegetativa s 32 inizio sporulazione s 29 inizio sporulazione (fase II) gp28 sporulazione intermedia gp fasi finali della sporulazione
 gp28 sporulazione intermedia. gp33-34 fasi finali della sporulazione.")
28
Terminazione Rho-indipendente
5’-A-A-A-G-C-C-G-C-C-G-N-N-N-N-N-C-C-G-G-C-G-G-C-U-U-U-U-U-U-U-3’ regione ricca in GC serie di U

29
la polimerasi continua la trascrizione
terminatore la polimerasi termina la trascrizione

30
Terminazione Rho-dipendente

33
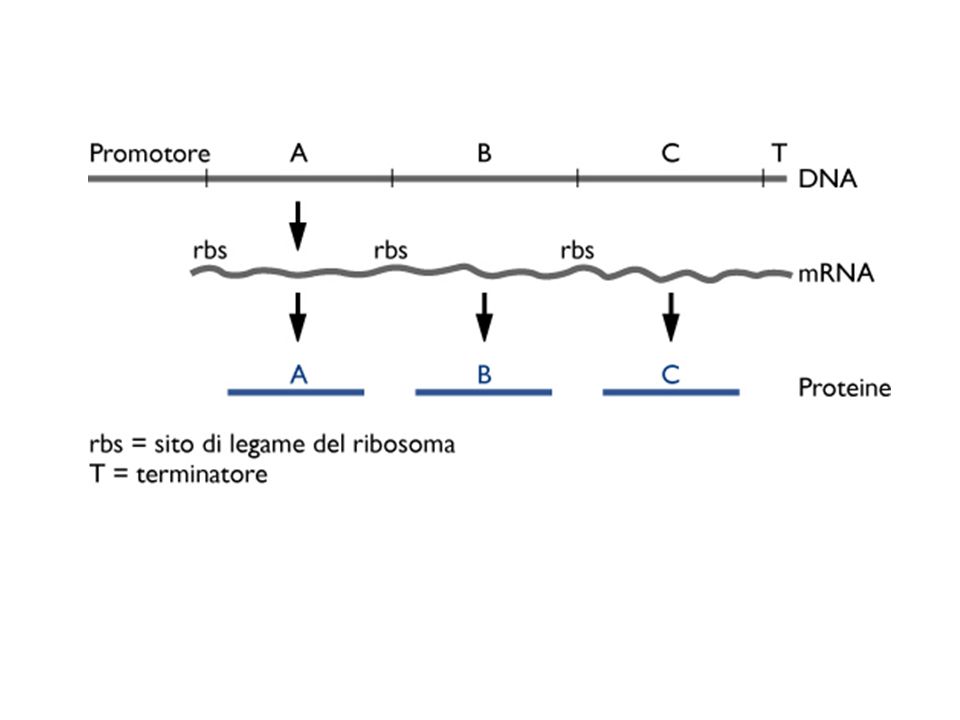
L’operone lac I P O Z Y A Geni strutturali lacZ: b-galattosidasi
DNA mRNA proteine lacZ: b-galattosidasi lacY: b-galattoside permeasi lacA: b-galattoside transacetilasi Geni strutturali mRNA policistronico regolazione coordinata: tutti i geni si esprimono all’unisono Il gene regolatore lacI codifica per la proteina regolatrice (repressore)
")
34
Induzione: sintesi di enzimi in risposta alla comparsa di un substrato specifico
Cellule di E. coli, in assenza di lattosio, contengono poche molecole di b-galattosidasi (<5) In presenza di lattosio , nel giro di pochi minuti, vengono sintetizzate fino a 5000 molecole di b-galattosidasi (5-10% delle proteine totali) glucosio galattosio La molecola che causa la produzione dell’enzima capace di metabolizzarla è chiamata induttore lattosio La capacità di agire da induttore è altamente specifica. Molecole strutturalmente affini al lattosio possono agire da induttori della b-galattosidasi ma non vengono metabolizzate: una molecola con questa caratteristica è chiamata induttore gratuito (IPTG) Se l’induttore è rimosso la sintesi degli enzimi si blocca
 In presenza di lattosio , nel giro di pochi minuti, vengono sintetizzate fino a 5000 molecole di b-galattosidasi (5-10% delle proteine totali) glucosio galattosio. La molecola che causa la produzione dell’enzima capace di metabolizzarla è chiamata induttore. lattosio. La capacità di agire da induttore è altamente specifica. Molecole strutturalmente affini al lattosio possono agire da induttori della b-galattosidasi ma non vengono metabolizzate: una molecola con questa caratteristica è chiamata induttore gratuito (IPTG) Se l’induttore è rimosso la sintesi degli enzimi si blocca.")
35
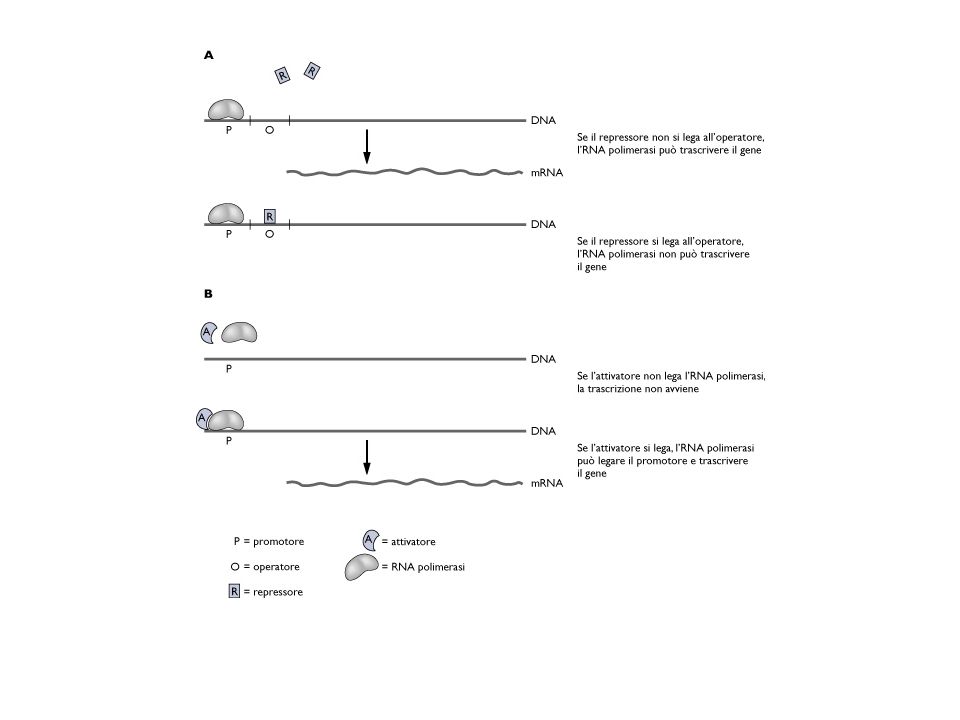
Z Y A O P I repressore RNApolimerasi Il repressore previene la trascrizione legandosi a una sequenza di DNA chiamata operatore. L’operone lac è sottoposto a regolazione negativa In presenza della proteina regolatrice è bloccata la trascrizione dei geni strutturali. La proteina regolatrice prende il nome di repressore

36
In che modo l’operone lac risponde all’induttore?
Z Y A O P I induttore Il repressore ha due siti di legame: un sito per il legame all’operatore e il secondo per il legame dell’induttore Quando il repressore lega l’induttore subisce una modificazione conformazionale che porta ad una diminuita affinità per il sito operatore (esempio di controllo allosterico)
")
37
Mutazioni a carico dell’operatore
Y A O P I repressore Mutazioni nel gene O (Oc) portano all’espressione costitutiva dei geni strutturali: vengono trascritti in assenza dell’induttore Costitutivi sono gli enzimi il cui livello è costante in tutte le condizioni di crescita
 portano all’espressione costitutiva dei geni strutturali: vengono trascritti in assenza dell’induttore. Costitutivi sono gli enzimi il cui livello è costante in tutte le condizioni di crescita.")
38
Mutazioni a carico del repressore
Y A O P I repressore Mutazioni nel gene I (Ic), a carico del sito di riconoscimento dell’operatore, portano all’espressione costitutiva dei geni strutturali
, a carico del sito di riconoscimento dell’operatore, portano all’espressione costitutiva dei geni strutturali.")
39
Mutazioni a carico del repressore
Y A O P I repressore induttore Mutazioni nel gene I (IS), a carico del sito di riconoscimento dell’induttore, sono responsabili della super-repressione: i geni strutturali non vengono più indotti in presenza dell’induttore
, a carico del sito di riconoscimento dell’induttore, sono responsabili della super-repressione: i geni strutturali non vengono più indotti in presenza dell’induttore.")
40
Un altro sistema di regolazione negativa:
la tossina di Corynebacterium diphtheriae La tossina viene prodotta quando il livello di ferro nella gola è basso gene regolatore P O geni per la tossina repressore ferro repressore trascrizione e produzione della tossina La molecola effettrice (il ferro) non è un induttore bensì un co-repressore Un sistema in cui la molecola effettrice coopera con il repressore per bloccare la trascrizione è definito reprimibile S80
 non è un induttore bensì un co-repressore. Un sistema in cui la molecola effettrice coopera con il repressore per bloccare la trascrizione è definito reprimibile. S80.")
41
La regolazione positiva dell’espressione genica
Nel controllo positivo, la proteina regolatrice promuove il legame della RNA-polimerasi al promotore gene1 gene2 gene3 gene regolatore P “activator binding site” induttore induttore RNApolimerasi attivatore L’attivatore non si lega al DNA in assenza dell’induttore B151

42
ABS P trascrizione ABS P trascrizione P ABS P = promotore ABS = activator binding site

43
Il regulone Il regulone è l’insieme di più geni o operoni sotto il controllo della stessa proteina regolatrice. I geni che lo costituiscono sono implicati nello stesso pathway Il regulone del maltosio è sotto controllo positivo attivatore + maltosio gene1 gene2 gene3 gene regolatore P “activator binding site” maltosio

44
Il regulone per la biosintesi dell’arginina
Gene o operone localizzazione arg A min arg B-C-E-H min arg D min arg F min arg G min Esempio di regulone sottoposto a controllo negativo: tutti i geni per la catena biosintetica dell’arginina sono regolati da un repressore

45
Sistemi di controllo globale dell’espressione genica
Un batterio può regolare molti geni diversi simultaneamente in risposta a cambiamenti ambientali. Spesso più operoni o reguloni diversi possono essere attivati o disattivati Il regulone è l’insieme di più geni o operoni sotto il controllo della stessa proteina regolatrice. I geni che lo costituiscono sono implicati nello stesso pathway Se geni e operoni appartengono a pathway differenti si parla di moduloni ESEMPI: regolazione mediante fattori sigma alternativi risposta SOS repressione da catabolita

46
Fattori sigma alternativi e regolazione globale dell’espressione genica
Subunità sigma Geni trascritti Segnale in E. coli s 70 maggior parte dei geni s 32 geni heat-shock temperatura elevata s 55 geni regolati dall’azoto carenza di ammonio s 38 geni da stress ossidativi agente ossidante Ogni fattore sigma riconosce una classe di promotori con specifiche sequenze consensus -35 e -10.

47
Repressione da catabolita
Quando un batterio cresce in un mezzo contenente glucosio come risorsa di energia, la sintesi di enzimi catabolici non correlati viene inibita In presenza di diverse fonti di energia, il batterio sceglie per prima quella più facilmente utilizzabile Gli enzimi per il catabolismo del glucosio sono costitutivi Circa 300 geni di E. coli sono regolati mediante repressione da catabolita

48
La crescita diauxica Tempo (ore) esaurimento del glucosio induzione bgalattosidasi utilizzazione del glucosio utilizzazione del lattosio Densità batterica Crescita in un terreno contenente glucosio e lattosio il glucosio inibisce la sintesi di b-galattosidasi durante la fase di latenza viene espresso l’operone lac e sintetizzata b-galattosidasi Il tasso di crescita può variare

49
La molecola effettrice della repressione da catabolita è AMP ciclico
H OH O Adenina CH2 HO-P=O ATP Adenilato ciclasi Il glucosio inibisce l’attività dell’adenilato ciclasi e il livello di cAMP

50
L’operone lac è regolato positivamente da CAP
Z Y A O P I CAP binding site cAMP CAP CAP attiva Ognuno degli operoni che CAP controlla è anche sotto il controllo di una proteina regolatrice specifica

51
L’attenuazione della trascrizione
L’operone triptofano trpR P O trpE trpD trpC trpB trpB repressore attivo repressore inattivo triptofano L’operone è sotto il controllo negativo del repressore codificato dal gene trpR Il triptofano agisce come corepressore, attivando il repressore e bloccando la trascrizione P262

52
L’attenuazione della trascrizione
L’operone triptofano trpR P O trpE trpD trpC trpB trpB leader 162 nt codoni trp 1 2 3 4 mRNA peptide leader (14AA) attenuatore
 attenuatore.")
53
1 2 3 4 1 2 3 4 1 2 3 4 mRNA Attenuatore (terminatore della
trascrizione) UUUUUUU 1 2 3 4 Se al segmento 1 viene impedito di appaiarsi con il segmento 2, quest’ultimo si appaia con il segmento 3. Il 4 rimane singolo e non si forma il terminatore
 UUUUUUU Se al segmento 1 viene impedito di appaiarsi con il segmento 2, quest’ultimo si appaia con il segmento 3. Il 4 rimane singolo e non si forma il terminatore.")
54
Il comportamento del ribosoma durante la traduzione del peptide leader regola l’attività della RNA polimerasi 1 2 3 4 mRNA peptide leader AUG UGA Se è presente sufficiente triptofano il ribosoma sintetizzerà il peptide leader arrivando fino al codone di stop. Il ribosoma sporge sul segmento 2 impedendogli l’appaiamento con il segmento 3 3 4 1 2 AUG UGA

55
La polimerasi continua
2 3 La polimerasi termina la trascrizione 2 3 4 2 3 4

56
Il comportamento del ribosoma durante la traduzione del peptide leader regola l’attività della RNA polimerasi 1 2 3 4 mRNA peptide leader AUG UGA Se manca il triptofano, il ribosoma si fermerà in corrispondenza dei 2 codoni trp adiacenti impedendo al segmento 1 di appaiarsi con 2. Il segmento 2 si associa con il segmento 3 2 3 4 1 AUG UGA

57
Agendo insieme, repressione e attenuazione possono coordinare la velocità di sintesi degli enzimi biosintetici per gli amminoacidi con la disponibilità degli amminoacidi e la velocità globale della sintesi proteica. Quando il triptofano è presente ad alte concentrazioni, qualsiasi RNA polimerasi non bloccata dal repressore probabilmente non oltrepasserà la sequenza dell’attenuatore. La repressione riduce la trascrizione di circa 70 volte e l’attenuazione la riduce ulteriormente di 8-10 volte: quando entrambi i meccanismi operani insieme, la trascrizione può essere ridotta di circa 600 volte. Sembra che l’attenuazione abbia un ruolo importante nella regolazione della biosintesi di molti amminoacidi Prescott pag. 262

58
Regolazione traduzionale
Operone per le tossine batteriche La proteina A entra nella cellula e provoca il danno La proteina B si lega al bersaglio (cellula umana) Due geni di un singolo operone possono essere tradotti in quantità diverse Rbs = “ribosome binding site” (Shine-Dalgarno) 5’-AGGAGG-3’
 Due geni di un singolo operone possono essere tradotti in quantità diverse. Rbs = ribosome binding site (Shine-Dalgarno) 5’-AGGAGG-3’")
59
Regolazione post-trascrizionale mediante RNA antisenso
L’RNA antisenso ha una sequenza complementare a una molecola di mRNA. Il legame dell’RNA antisenso può bloccare la traduzione dell’ mRNA.

60
Sistema host cell killing del plasmide R1
Il sistema hok/sok di alcuni plasmidi a basso numero di copie assicura il mantenimento del plasmide all’interno di una popolazione batterica hok mRNA: emivita 20 min hok SD 128 bp sok sok mRNA: emivita < 1-2 min

61
La risposta all’osmolarità in E. coli
EnvZ OmpF sintesi di OmpC micF RNA 174 b OmpC OmpR mRNA per OmpF

62
Regolazione post-traduzionale
modificazione allosterica Controllo tipico della via biosintetica degli amminoacidi Un enzima della catena può legare il prodotto finale che funziona da inibitore Se il prodotto finale viene sintetizzato in eccesso o se diventa disponibile nell’ambiente, non è più conveniente per il batterio continuare a sintetizzarlo Questo tipo di controllo è anche definito inibizione a feedback

Presentazioni simili














 viene copiata in una sequenza complementare di RNA dall’enzima.>")




