Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Canali Voltaggio-dipendenti per il Ca2+

2
La struttura molecolare dei canali del Ca2+ è nel suo insieme del tutto simile a quella dei NaV, sebbene (piccole) differenze nelle sequenze cambino profondamente la selettività, il gating e la sensibilità ai farmaci. Le subunità alfa sono costantemente associate a subunità accessorie, in numero maggiore di quelle del Na+: α2, β, γ e δ. I canali del Ca VD sono tutti bloccati (pur con sensibilità diverse) dal Cadmio (Cd2+) e dai “metalli pesanti” (Ni2+, Co2+ e La3+).
 dal Cadmio (Cd2+) e dai metalli pesanti (Ni2+, Co2+ e La3+).")
3
Organizzazione strutturale dei canali del Ca VD
Subunità a1: 4 domini omologhi (I-IV), con 6 segmenti transmembrana ciascuno; Subunità b: intracellulare, costituita da più a eliche; Subunità a2d: in giallo le regioni idrofobiche; I siti di interazione tra le subunità sono indicati da frecce bipolari.
, con 6 segmenti transmembrana ciascuno; Subunità b: intracellulare, costituita da più a eliche; Subunità a2d: in giallo le regioni idrofobiche; I siti di interazione tra le subunità sono indicati da frecce bipolari.")
4
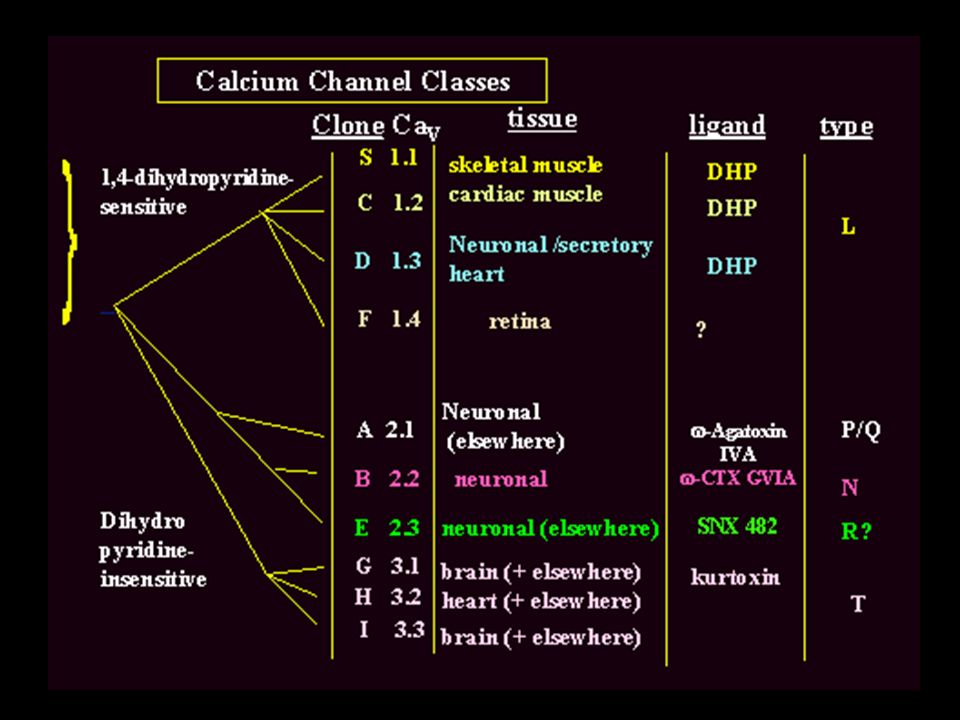
Classificazione in base alle proprietà elettrofisiologiche:
a) Canali LVA (low-voltage-activated (-70mV); avendo una spiccata inattivazione voltaggio-dipendente, sono stati chiamati canali T (“transient”). Sono sensibili all’amiloride. b) Canali HVA (high-voltage-activated (-30m/-20V); hanno un’inattivazione molto più lenta. 0 mV 400 pA 30 ms Correnti di Ca HVA -40 mV 100 pA 30 ms Correnti di Ca LVA Poi s’è visto che si tratta di una famiglia comprendente molti membri, che sono stati indicati da lettere (P, Q, R ecc.)
 Canali LVA (low-voltage-activated (-70mV); avendo una spiccata inattivazione voltaggio-dipendente, sono stati chiamati canali T ( transient ). Sono sensibili all’amiloride. b) Canali HVA (high-voltage-activated (-30m/-20V); hanno un’inattivazione molto più lenta. 0 mV. 400 pA. 30 ms. Correnti di Ca HVA. -40 mV. 100 pA. 30 ms. Correnti di Ca LVA. Poi s’è visto che si tratta di una famiglia comprendente molti membri, che sono stati indicati da lettere (P, Q, R ecc.)")
6
I diversi sottotipi di canali del Ca inattivano in maniera differente
a1C (L) a1E (R) a1G (T) 50 ms
 a1E (R) a1G (T) 50 ms.")
7
Possibile modello di inattivazione VD dei canali del Ca
Il linker intracellulare tra i domini I e II potrebbe agire da particella inattivante che fisicamente occluderebbe il poro del canale interagendo, almeno in parte, con le regioni S6 dei domini II e III del canale. La subunità b del canale del Ca si associa con il linker tra i domini I-II modificando le proprietà dell’inattivazione; essa potrebbe interagire sia con la subunità a1 che con la membrana.

8
Possibile modello di inattivazione Ca-dipendente dei canali del Ca
Un’elevata concentrazione di Ca intracellulare innesca l’inattivazione dei canali di tipo L. Il sensore per il Ca sarebbe una molecola di calmodulina (CaM). CaM è costitutivamente legata al complesso del canale. L’inattivazione avverrebbe tramite un’interazione Ca-dipendente della CaM legata con un motivo IQ-simile sulla coda carbossilica di a1C. È possibile che un meccanismo analogo intervenga anche a livello dei canali N, P/Q ed R (Peterson, Neuron, 1999). CHIUSO APERTO INATTIVATO Ca2+ Voltaggio Nello stato chiuso (iperpolarizzato) l’accesso della CaM alla regione IQ-simile è ridotto. Nello stato aperto (depolarizzato) la CaM legata ha un rapido accesso al dominio IQ-simile. L’accumulo di Ca2+ sulla bocca interna del canale porta all’attivazione Ca-dip. della CaM. L’interazione di CaM-Ca2+ con la regione IQ-simile induce l’inattivazione del canale.
. CaM è costitutivamente legata al complesso del canale. L’inattivazione avverrebbe tramite un’interazione Ca-dipendente della CaM legata con un motivo IQ-simile sulla coda carbossilica di a1C. È possibile che un meccanismo analogo intervenga anche a livello dei canali N, P/Q ed R (Peterson, Neuron, 1999). CHIUSO. APERTO. INATTIVATO. Ca2+ Voltaggio. Nello stato chiuso (iperpolarizzato) l’accesso della CaM alla regione IQ-simile è ridotto. Nello stato aperto (depolarizzato) la CaM legata ha un rapido accesso al dominio IQ-simile. L’accumulo di Ca2+ sulla bocca interna del canale porta all’attivazione Ca-dip. della CaM. L’interazione di CaM-Ca2+ con la regione IQ-simile induce l’inattivazione del canale.")
9
Canali VD per il Ca2+ di tipo L nel muscolo liscio
Nel muscolo liscio, il reticolo sarcoplasmatico è praticamente assente. Qui l’attivazione dell’apparato contrattile (non organizzato in sarcomeri) è totalmente dipendente dal Ca2+ che entra attraverso i canali L della membrana plasmatica. Il fenomeno dell’accoppiamento eccitazione contrazione nei tessuti muscolari illustra bene la doppia importanza dei canali del Ca2+: fanno entrare nella cellula ioni con carica positiva (similmente ai NaV) fanno anche comparire nella cellula un vero e proprio secondo messaggero (gli ioni Ca2+) che ha uno straordinario potere regolatore di molti processi intracellulari
 è totalmente dipendente dal Ca2+ che entra attraverso i canali L della membrana plasmatica. Il fenomeno dell’accoppiamento eccitazione contrazione nei tessuti muscolari illustra bene la doppia importanza dei canali del Ca2+: fanno entrare nella cellula ioni con carica positiva (similmente ai NaV) fanno anche comparire nella cellula un vero e proprio secondo messaggero (gli ioni Ca2+) che ha uno straordinario potere regolatore di molti processi intracellulari.")
10
Canali voltaggio-dipendenti per il Ca2+ di tipo L
I canali “L” o recettori per le di-idro-piridine (DHP, come la Nifedipina o la nimodipina) hanno un’enorme importanza in Fisiologia. Sono tipici del muscolo scheletrico, ma sono presenti anche nel cuore (*) e nel sistema nervoso. Hanno un’inattivazione più o meno lenta, che è anche Ca-dipendente. (*) il bloccante Verapamil viene impiegato nella cura della tachicardia parossitica
 hanno un’enorme importanza in Fisiologia. Sono tipici del muscolo scheletrico, ma sono presenti anche nel cuore (*) e nel sistema nervoso. Hanno un’inattivazione più o meno lenta, che è anche Ca-dipendente. (*) il bloccante Verapamil viene impiegato nella cura della tachicardia parossitica.")
11
Canali VD per il Ca2+ di tipo L nel muscolo scheletrico
Nel muscolo scheletrico, i canali L sono addensati nei tubuli trasversi del reticolo sarcoplasmatico, dove funzionano da “sensori del voltaggio”. Sono accoppiati (con l’ansa di connessione II-III) ad altri canali del Ca2+ (Ca-dipendenti) presenti nei tubuli longitudinali del ret.sarc., detti “recettori della Ryanodina”. Si attiva così un il rilascio di Ca che attiva con grande rapidità (per feedback positivo) l’apparato contrattile.
 ad altri canali del Ca2+ (Ca-dipendenti) presenti nei tubuli longitudinali del ret.sarc., detti recettori della Ryanodina . Si attiva così un il rilascio di Ca che attiva con grande rapidità (per feedback positivo) l’apparato contrattile.")
12
Modello per il rilascio voltaggio-dipendente del Ca2+
Sensore del volt. Canale di rilascio TT RS A riposo Ca 2+ Vm + - Depolarizzata Ca 2+ Ca 2+ Ca 2+ Ca 2+ Ca 2+ Ca 2+

13
Canali VD per il Ca2+ di tipo L nel muscolo cardiaco
Accoppiamento EC Cardiaco (Ca2+-induced-Ca2+-release) SR + TT SR TT Vm + + Ca2+ Canale del Ca2+ Canale di rilascio
 SR. + TT. SR. TT. Vm. + + Ca2+ Canale del Ca2+ Canale di rilascio.")
14
Ruolo dei canali voltaggio-dipendenti per il Ca nel PdA cardiaco (tessuto di lavoro)
Solitamente l’ingresso di Ca2+ attraverso canali VD accompagna (e prolunga) i pda al Na+. E’ il caso delle fibrocellule muscolari scheletriche, ma soprattutto delle fibrocellule muscolari cardiache. Qui l’ingresso di Ca2+ non solo potenzia l’effetto del Na+ nella fase iniziale del pda (depolarizzazione ed overshoot), ma perdura anche quando i canali del Na+ si sono inattivati (l’inattivazione dei canali del Ca2+ c’è, ma è incompleta), dando luogo ad un lungo “plateau”. Domanda: ma se durante il plateau entrano cariche positive sotto forma di ioni Ca2+, perché il pdm si mantiene costante <attorno allo zero> ? Risposta: perché durante il “plateau” la corrente entrante di Ca2+ è esattamente controbilanciata da una corrente uscente di K+ <attraverso canali al K+
 i pda al Na+. E’ il caso delle fibrocellule muscolari scheletriche, ma soprattutto delle fibrocellule muscolari cardiache. Qui l’ingresso di Ca2+ non solo potenzia l’effetto del Na+ nella fase iniziale del pda (depolarizzazione ed overshoot), ma perdura anche quando i canali del Na+ si sono inattivati (l’inattivazione dei canali del Ca2+ c’è, ma è incompleta), dando luogo ad un lungo plateau . Domanda: ma se durante il plateau entrano cariche positive sotto forma di ioni Ca2+, perché il pdm si mantiene costante <attorno allo zero> Risposta: perché durante il plateau la corrente entrante di Ca2+ è esattamente controbilanciata da una corrente uscente di K+ <attraverso canali al K+")
15
Questo lunghissimo “plateau” del pda cardiaco è funzionalmente importante per due buone ragioni:
a) il Ca2+ che entra attraverso i canali (L) della membrana plasmatica attiverà il “Ca-activated Ca-release” del reticolo sarcoplasmatico, quindi la contrazione del cuore; b) mantenendo depolarizzata la membrana, la rende ineccitabile per tutta la sua durata (perché mantiene inattivati i canali del Na+). In altre parole: durante il plateau, il cuore si trova in uno stato di refrattarietà assoluta, quindi per tutta la durata della contrazione (della sistole) non può essere nuovamente eccitato. La situazione è molto diversa da quella che si ha nel muscolo scheletrico, nel quale la refrattarietà assoluta termina ancor prima che inizi la contrazione.
 il Ca2+ che entra attraverso i canali (L) della membrana plasmatica attiverà il Ca-activated Ca-release del reticolo sarcoplasmatico, quindi la contrazione del cuore; b) mantenendo depolarizzata la membrana, la rende ineccitabile per tutta la sua durata (perché mantiene inattivati i canali del Na+). In altre parole: durante il plateau, il cuore si trova in uno stato di refrattarietà assoluta, quindi per tutta la durata della contrazione (della sistole) non può essere nuovamente eccitato. La situazione è molto diversa da quella che si ha nel muscolo scheletrico, nel quale la refrattarietà assoluta termina ancor prima che inizi la contrazione.")
16
Il muscolo scheletrico infatti (per fortuna
Il muscolo scheletrico infatti (per fortuna !) è tetanizzabile, cioè le singole “scosse” possono sommarsi tra loro fino alla completa fusione, producendo un aumento della forza contrattile. Di fatto, la maggior parte delle nostre contrazioni sono dei “tetani”. Il cuore invece (per fortuna !) non è “tetanizzabile”: qualunque stimolo “ectopico” è inefficace per tutta la durata della sistole (I), e può produrre una seconda contrazione (“extrasistole”) solo mentre il cuore si sta rilasciando..
 è tetanizzabile, cioè le singole scosse possono sommarsi tra loro fino alla completa fusione, producendo un aumento della forza contrattile. Di fatto, la maggior parte delle nostre contrazioni sono dei tetani . Il cuore invece (per fortuna !) non è tetanizzabile : qualunque stimolo ectopico è inefficace per tutta la durata della sistole (I), e può produrre una seconda contrazione ( extrasistole ) solo mentre il cuore si sta rilasciando..")
17
Potenziali d’azione al Ca2+ nelle cellule pacemaker cardiache
Non mancano esempi di pda sostenuti solo dall’ingresso autorigenerativo di Ca2+, come il gigantesco pda delle cellule del Purkinje del cervelletto, evocato dalla stimolazione delle fibre rampicanti (attivazione di canali P). Un altro esempio funzionalmente molto rilevante è quello del miocardio di conduzione (tessuto “pacemaker” del cuore). Questi pda al Ca2+ si generano spontaneamente (o meglio, per effetto dell’automatismo tipico del pacemaker), e poi si propagano a tutte le fibrocellule del miocardio di lavoro (che di per sé resterebbero in riposo). Queste, eccitate, genereranno i pda provvisti di “plateau” che abbiamo visto prima. Curiosamente, nel miocardio di conduzione il contributo del Na+ non manca perché i canali del Na+ siano assent:. questi ci sono, ma non entrano in gioco perché sono sempre inattivati (nel tessuto pacemaker, il potenziale di membrana non diviene mai sufficientemente negativo da rimuoverne l’inattivazione).
. Un altro esempio funzionalmente molto rilevante è quello del miocardio di conduzione (tessuto pacemaker del cuore). Questi pda al Ca2+ si generano spontaneamente (o meglio, per effetto dell’automatismo tipico del pacemaker), e poi si propagano a tutte le fibrocellule del miocardio di lavoro (che di per sé resterebbero in riposo). Queste, eccitate, genereranno i pda provvisti di plateau che abbiamo visto prima. Curiosamente, nel miocardio di conduzione il contributo del Na+ non manca perché i canali del Na+ siano assent:. questi ci sono, ma non entrano in gioco perché sono sempre inattivati (nel tessuto pacemaker, il potenziale di membrana non diviene mai sufficientemente negativo da rimuoverne l’inattivazione).")
18
Interazione tra canali “T” e canali “f”
L’esempio del pda (al Ca2+) del pacemaker cardiaco ci offre l’opportunità di parlare di un altro tipo di canali voltaggio-dipendenti: i canali “f” (“funny”), espressi anche in molti neuroni autoritmici del SNC (dove vengono chiamati canali “h”) I canali del Ca2+ di tipo T (e in minor misura L) sostengono il pda, ma per essere attivati necessitano di una depolarizzazione della membrana. Questa, nelle cellule autoritmiche, avviene “spontaneamente” e si chiama prepotenziale. Nel tessuto pacemaker, il prepotenziale (e con esso l’automatismo cardiaco) è generato dall’apertura dei canali-f .
 del pacemaker cardiaco ci offre l’opportunità di parlare di un altro tipo di canali voltaggio-dipendenti: i canali f ( funny ), espressi anche in molti neuroni autoritmici del SNC (dove vengono chiamati canali h ) I canali del Ca2+ di tipo T (e in minor misura L) sostengono il pda, ma per essere attivati necessitano di una depolarizzazione della membrana. Questa, nelle cellule autoritmiche, avviene spontaneamente e si chiama prepotenziale. Nel tessuto pacemaker, il prepotenziale (e con esso l’automatismo cardiaco) è generato dall’apertura dei canali-f .")
19
La struttura dei canali HCN
Appartengono alla classe dei canali HCN: “canali cationici attivati dall’iperpolarizzazione”. Vengono attivati alla fine di ogni pda, quando la membrana si iperpolarizza. La loro apertura genera una corrente entrante che, depolarizzando la membrana, produce il prepotenziale, quindi il pda successivo. Ciascuna delle 4 subunità presenta: 6-DTM come i canali Kv S4 riconosciuto come sensore del voltaggio per l'elevata presenza di residui carichi positivamente motivo GYG nel linker S5-S6, tipico del filtro di selettività dei canali Kv nel C-terminale è presente un dominio CNB come nei canali CNG si pensa che i canali siano formati da TETRAMERI come i canali Kv indietro

20
Modulazione dei Canali “f”
Di grande importanza è la “modulazione” (variazione della sensibilità al voltaggio) dei canali f operata dall’orto- e dal para-simpatico tramite i rispettivi neurotrasmettitori <noradrenalina(+adrenalina) ed acetilcolina>. Modulando i canali f, l’orto- ed il para-simpatico regolano la frequenza cardiaca (!!), come se fossero l’uno l’acceleratore e l’altro il freno di un’automobile. Questi neurotrasmettitori agiscono su recettori accoppiati a proteine-G e fanno rispettivamente aumentare e diminuire il livello intracellulatre di AMPc. I canali f sono voltaggio-dipendenti, ma anche chemio-dipendenti (dal versante intracellulare), una condizione tutt’altro che infrequente.
 dei canali f. operata dall’orto- e dal para-simpatico tramite i rispettivi neurotrasmettitori. <noradrenalina(+adrenalina) ed acetilcolina>. Modulando i canali f, l’orto- ed il para-simpatico regolano la frequenza cardiaca (!!), come se fossero l’uno l’acceleratore e l’altro il freno di un’automobile. Questi neurotrasmettitori agiscono su recettori accoppiati a proteine-G e fanno rispettivamente aumentare e diminuire il livello intracellulatre di AMPc. I canali f sono voltaggio-dipendenti, ma anche chemio-dipendenti (dal versante intracellulare), una condizione tutt’altro che infrequente.")
21
Canali voltaggio-dipendenti per il Ca2+ di tipo N e P/Q nell’esocitosi
Un altro esempio di enorme importanza funzionale è l’ “esocitosi regolata” di vescicole (v. rilascio di insulina: qui sono implicati canali del Ca2+ di tipo L ) in tutte le cellule secernenti. Un caso particolarissimo di “esocitosi regolata” è la liberazione di vescicole contenenti neurotrasmettitore nella trasmissione sinaptica. Qui operano canali “N” (per neuronal, bloccati dall’ ω-Conotossina), che sono esclusivi del SNC, ma anche canali “P/Q” (bloccati dall’ ω-Agatossina) ed R, per i quali non è noto alcun bloccante specifico.
 in tutte le cellule secernenti. Un caso particolarissimo di esocitosi regolata è la liberazione di vescicole contenenti neurotrasmettitore nella trasmissione sinaptica. Qui operano canali N (per neuronal, bloccati dall’ ω-Conotossina), che sono esclusivi del SNC, ma. anche canali P/Q (bloccati dall’ ω-Agatossina) ed R, per i quali non è noto alcun bloccante specifico.")
22
La trasmissione sinaptica
La combinazione del neurotrasmettitore con i propri recettori presenti nella membrana postsinaptica consentirà la trasmissione sinaptica (una depolarizzazione [EPSP] nelle sinapsi eccitatorie o un’ iperpolarizzazione [IPSP] nelle sinapsi inibitorie…)
")
23
Eventi pre-simnaptici
Questi canali sono addensati nelle terminazioni presinaptiche, dove sono essenziali per il rilascio di neurotrasmettitore. In particolare sono presenti al centro delle zone attive alle quali sono ancorate le vescicole sinaptiche pronte al rilascio (“pool” di rilascio). Le zone attive sono formazioni del citoscheletro altamente ordinate, visibili dal versante citoplasmatico della membrana presinaptica). In corrispondenza delle zone attive, i lipidi della membrana sono organizzati a formare dei “rafts” (zattere). L’arrivo del pda e la conseguente depolarizzazione della membrana presinaptica apre i canali del Ca2+ …. …. ed il Ca2+ fa scattare un complesso sistema di controllo. La fusione delle vescicole promossa dal Ca2+ è preceduta da fasi preliminari:
. Le zone attive sono formazioni del citoscheletro altamente ordinate, visibili dal versante citoplasmatico della membrana presinaptica). In corrispondenza delle zone attive, i lipidi della membrana sono organizzati a formare dei rafts (zattere). L’arrivo del pda e la conseguente depolarizzazione della membrana presinaptica apre i canali del Ca2+ …. …. ed il Ca2+ fa scattare un complesso sistema di controllo. La fusione delle vescicole promossa dal Ca2+ è preceduta da fasi preliminari:")
24
“Docking” (attracco) …
La T. botulinica è il veleno più potente che si conosca Anche la T.tetanica agisce sul sistema di controllo della liberazione di vescicole sinaptiche B) “Priming” (preparazione) … La latratossina, il principio attivo del veleno della Vedova Nera, provoca la liberazione spontanea ed incontrollata delle vescicole C) all’arrivo del pda si ha la fusione Ca-dipendente, (preceduta dalla formazione di un poro attraverso il quale passerà il neurotrasmettitore), che consente l’esocitosi delle vescicole ancorate alle zone attive (“pool di rilascio”).
 Priming (preparazione) … La latratossina, il principio attivo del veleno della Vedova Nera, provoca la liberazione spontanea ed incontrollata delle vescicole. C) all’arrivo del pda si ha la fusione Ca-dipendente, (preceduta dalla formazione di un poro attraverso il quale passerà il neurotrasmettitore), che consente l’esocitosi delle vescicole ancorate alle zone attive ( pool di rilascio ).")
25
Ruolo dei canali del Ca2+ al di fuori delle zone attive della membrana pre-sinaptica
Canali VD per il Ca2+ (solitamente del tipo L) sono anche presenti nella membrana presinaptica al di fuori delle zone attive (A). La loro attivazione (B) serve a distaccare le vescicole del “pool di deposito” dai filamenti di actina del citoscheletro, quindi a rifornire il “pool di rilascio”. Lo schema C illustra il processo in maggior dettaglio. Anche in questo caso si mette bene in evidenza il ruolo del Ca2+ come secondo messaggero intracellulare !
 sono anche presenti nella membrana presinaptica al di fuori delle zone attive (A). La loro attivazione (B) serve a distaccare le vescicole del pool di deposito dai filamenti di actina del citoscheletro, quindi a rifornire il pool di rilascio . Lo schema C illustra il processo in maggior dettaglio. Anche in questo caso si mette bene in evidenza il ruolo del Ca2+ come secondo messaggero intracellulare !")
26
Correnti T neuronali e spikes al Ca2+
Vhold-90 mV

27
Vedi registrazione Attività ritmica spontanea in un neurone talamico
Attività oscillatoria -65 mV 1 s Bursts di PdA dovuti all’interazione della corrente di Ca2+ IT con la corrente pacemaker cationica entrante Ih -65 mV PdA al Na+ Spike al Ca 2+ attivazione Ih attivazione IT deattivazione Ih inattivazione IT rimozione Potenziale pacemaker Vedi registrazione Quale meccanismo innesca questa attività autoritmica?

28
Organizzazione della corteccia cerebrale
La corteccia cerebrale è costituita da sei strati. Le afferenze talamiche arrivano al IV strato. Le fibre efferenti (motrici) partono dal V e VI strato
 partono dal V e VI strato.")
29
- TALAMO CORTECCIA + NRT
GABA GLUT CORTECCIA NUCLEI di RELAY TALAMO NRT afferenze talamiche Il neurone GABAergico determina sul neurone del nucleo di relay un PPSI che genera iperpolarizzazione e fa aprire i canali “h” innescando l’attività autoritmica in tali neuroni NRT: nucleo reticolare del talamo

30
La genesi di un burst di pda in un singolo neurone del nucleo reticolare del talamo (NRT) può generare un IPSP nelle cellule talamocorticali del nucleo di relay che è sufficientemente ampio da generare uno spike al Ca2+ e un burst di pda. Ampiezza e andamento temporale dell’IPSP generato nel neurone talamocorticale del nucleo di relay dipendono fortemente dal tipo di scarica generata a livello del NRT, e viceversa.

31
In realtà, i neuroni talamocorticali nei nuclei di relay del talamo hanno due modalità di scarica
(vedi registrazioni)
")
32
In realtà, i neuroni talamocorticali dei nuclei di relè presentano anche una seconda modalità di scarica (vedi registrazioni) L’applicazione di ACh o NE causa una simile depolarizzazione dei neuroni talamocorticali attraverso la riduzione di una corrente di K+ di "leakage" attiva a riposo. La depolarizzazione di neuroni talamocorticali in seguito ad una iniezione intracellulare di corrente provoca l’abolizione della scarica ritmica a burst e la sua sostituzione con un’attività tonica. L’abolizione della scarica ritmica a burst è causata dall’inattivazione della corrente di Ca2+ T e dalla mancata attivazione della corrente “Ih”.

33
Gli stadi del sonno sono caratterizzati da cambiamenti nelle registrazioni EEG
La sostituzione della scarica ritmica con quella tonica è simile alla modificazione dell’attività elettrica che si può registrare in corteccia nella transizione dal sonno ad onda lenta alla veglia (o al sonno REM). Veglia – occhi aperti Veglia – occhi chiusi Sonno non REM – stadio I Sonno non REM – stadi II e III Sonno non REM – stadio IV Sonno REM Sonno non-REM, Stadio IV (SWS; Slow-Wave-Sleep ‘sonno profondo’): onde ampie a bassa frequenza (~1 Hz, ‘onde delta’) sono attivi pochi neuroni ma altamente sincronizzati
. Veglia – occhi aperti. Veglia – occhi chiusi. Sonno non REM – stadio I. Sonno non REM – stadi II e III. Sonno non REM – stadio IV. Sonno REM. Sonno non-REM, Stadio IV. (SWS; Slow-Wave-Sleep ‘sonno profondo’): onde ampie a bassa frequenza (~1 Hz, ‘onde delta’) sono attivi pochi neuroni ma altamente sincronizzati.")
34
Cosa suggeriscono tali risultati?
Attività elettrica registrata contemporaneamente da neuroni corticali e talamici coinvolti nel sonno SWS (Slow-Wave-Sleep) e durante la veglia Registr. intracell dal nucleo di relay. Registraz. extracell. da neurone del NRT Registraz. extracell. da neurone corticale. Tali risultati indicano che tutte e tre le tipologie di neuroni esibiscono i due diversi stati dell’EEG: una scarica ritmica a burst durante il sonno ad onde lente, e un’attività tonica durante la veglia. Cosa suggeriscono tali risultati?
 e durante la veglia. Registr. intracell. dal nucleo di relay. Registraz. extracell. da neurone del NRT. Registraz. extracell. da neurone corticale. Tali risultati indicano che tutte e tre le tipologie di neuroni esibiscono i due diversi stati dell’EEG: una scarica ritmica a burst durante il sonno ad onde lente, e un’attività tonica durante la veglia. Cosa suggeriscono tali risultati")
35
Cellule pacemaker talamiche: cosa le attiva e cosa le spegne?
• le cellule pacemaker del nucleo di relay del talamo che scaricano ritmicamente a bursts inducono i neuroni corticali a scaricare in maniera sincrona nel pattern dell’SWS (Slow-Wave-Sleep) dell’EEG • durante la veglia: neuroni colinergici (ACh), noradrenergici (NE) e serotoninergici (5-HT) sono attivi inibiscono le cellule pacemaker talamiche • durante il sonno SWS, l’attività delle cellule che liberano ACh, NE e 5-HT diminuisce le cellule pacemaker talamiche sono attive
 dell’EEG. • durante la veglia: neuroni colinergici (ACh), noradrenergici (NE) e serotoninergici (5-HT) sono attivi inibiscono le cellule pacemaker talamiche. • durante il sonno SWS, l’attività delle cellule che liberano ACh, NE e 5-HT diminuisce le cellule pacemaker talamiche sono attive.")
36
Canali P e spikes complessi nelle cellule del Purkinje
Gli input delle fibre parallele e rampicanti hanno effetti molto diversi sulle cellule del Purkinje Molte fibre parallele (~200,000) prendono contatto con una singola cellula del Purkinje (convergenza). La scarica generata è di tipo tonico (spikes semplici) e si sovrappone ad una attività spontanea basale. Un’unica fibra rampicante contatta ciascuna cellula del Purkinje formando molte sinapsi. Ciascun singolo input genera una rapida scarica di potenziali d’azione di latenza e durata brevi (spikes complessi: azione fasica rapida). Si ritiene che l’attivazione delle fibre muscoidi possa rendere silente per alcune centinaia di ms la scarica evocata nelle cellule del Purkinje dalle fibre rampicanti
 prendono contatto con una singola cellula del Purkinje (convergenza). La scarica generata è di tipo tonico (spikes semplici) e si sovrappone ad una attività spontanea basale. Un’unica fibra rampicante contatta ciascuna cellula del Purkinje formando molte sinapsi. Ciascun singolo input genera una rapida scarica di potenziali d’azione di latenza e durata brevi (spikes complessi: azione fasica rapida). Si ritiene che l’attivazione delle fibre muscoidi possa rendere silente per alcune centinaia di ms la scarica evocata nelle cellule del Purkinje dalle fibre rampicanti.")
37
Spikes semplici e complessi generati dalle cellule del Purkinje
Le cellule di Purkinje generano potenziali d’azione spontanei anche in assenza di inputs sinaptici. La frequenza media durante la veglia è di circa 40 Hz (40 spikes per sec). La frequenza può aumentare per stimolazione delle fibre parallele. I treni in figura mostrano una mistura di cosiddetti spikes semplici e complessi. Spikes semplici S. complessi Gli spikes complessi sono spesso seguiti da una pausa di alcune centinaia di msec durante la quale l’attività degli spikes semplici è soppressa. 50 ms Apertura di canali del Ca2+ P-type Apertura di canali del Na+ Uno spike complesso è una sequenza stereotipata di potenziali d’azione con intervalli molto brevi tra uno spike e l’altro e ampiezze decrescenti. Gli spikes complessi, (che si susseguono ad una velocità basale di circa 1 Hz, massimo 10 Hz) sono costantemente associati all’attivazione delle fibre rampicanti. Gli spikes semplici sono prodotti da una combinazione di attività basale e di inputs dalle fibre parallele.
. La frequenza può aumentare per stimolazione delle fibre parallele. I treni in figura mostrano una mistura di cosiddetti spikes semplici e complessi. Spikes semplici. S. complessi. Gli spikes complessi sono spesso seguiti da una pausa di alcune centinaia di msec durante la quale l’attività degli spikes semplici è soppressa. 50 ms. Apertura di canali del Ca2+ P-type. Apertura di canali del Na+ Uno spike complesso è una sequenza stereotipata di potenziali d’azione con intervalli molto brevi tra uno spike e l’altro e ampiezze decrescenti. Gli spikes complessi, (che si susseguono ad una velocità basale di circa 1 Hz, massimo 10 Hz) sono costantemente associati all’attivazione delle fibre rampicanti. Gli spikes semplici sono prodotti da una combinazione di attività basale e di inputs dalle fibre parallele.")
38
Canalopatie dove sono coinvolti canali del Ca2+ VD
Si è osservato che la paralisi periodica ipokaliemica, che si manifesta con debolezza muscolare che compare all’adolescenza, è associata a mutazioni del gene del canale L (cromosoma 1). I segmenti S4 dei domini II o IV hanno meno Arg (quindi i canali hanno minor sensibilità al voltaggio) e l’inattivazione è più rapida. E’ comprensibile che queste caratteristiche riducano il rilascio intracellulare di Ca2+. Si pensa che mutazioni del gene del RYR (cromosoma 19) siano la causa della ipertermia maligna: un’anomala risposta all’anestesia generale con alotano e succinilcolina (convulsioni muscolari dovute ad un aumentato rilascio di Ca2+ intracellulare, aumento della temperatura corporea, squilibrio idrosalino) che risulta fatale in 1 caso su La situazione è recentemente migliorata grazie alla diagnosi precoce ed all’uso del dantrolene, che inibisce il rilascio di Ca2+ da parte del ret.sarc.
. I segmenti S4 dei domini II o IV hanno meno Arg (quindi i canali hanno minor sensibilità al voltaggio) e l’inattivazione è più rapida. E’ comprensibile che queste caratteristiche riducano il rilascio intracellulare di Ca2+. Si pensa che mutazioni del gene del RYR (cromosoma 19) siano la causa della ipertermia maligna: un’anomala risposta all’anestesia generale con alotano e succinilcolina (convulsioni muscolari dovute ad un aumentato rilascio di Ca2+ intracellulare, aumento della temperatura corporea, squilibrio idrosalino) che risulta fatale in 1 caso su La situazione è recentemente migliorata grazie alla diagnosi precoce ed all’uso del dantrolene, che inibisce il rilascio di Ca2+ da parte del ret.sarc.")
39
Ogni ione può essere trasportato da numerosi tipi di canali ionici, ognuno con proprietà di attivazione distinte. Questo conferisce una grande flessibilità alla codificazione dei pda da parte dei neuroni ed è un elemento chiave per conferire al cervello una notevole capacità computazionale. ripolarizzazione aumentata firing ritardato diminuzione della risposta alla ripolarizzazione velocità di firing rallentata firing a scoppio (burst) mancanza di burst depolarizzazione (inattiva IT) + KCa1 + KM + KCa2 Rallentamento della scarica; aumento dell’AHP La KCa determina adattamento e AHP di lunga durata La KM determina adattamento È possibile regolare la frequenza, la durata, la regolarità, il ritardo delle scariche di pda, generando le basi per costruire il codice neuronale.
 mancanza di burst. depolarizzazione. (inattiva IT) + KCa1. + KM. + KCa2. Rallentamento della scarica; aumento dell’AHP. La KCa determina adattamento e AHP di lunga durata. La KM determina adattamento. È possibile regolare la frequenza, la durata, la regolarità, il ritardo delle scariche di pda, generando le basi per costruire il codice neuronale.")
40
FINE

41
IP3 RECEPTOR + TETRAMER (3 ISOFORMS) Ca 2+ CaMKINASE II cAMPPK BINDING
CALMODULIN (TYPES I&II) NH2 COOH IP3 BINDING DOMAIN COUPLING TRANS- MEMBRANE ATP P Ca2+ IP3 RECEPTOR PLC PIP2 DAG PKC + cAMPPK Ca 2+ CaMKINASE II TETRAMER (3 ISOFORMS)
 NH2. COOH. IP3. BINDING. DOMAIN. COUPLING. TRANS- MEMBRANE. ATP. P. Ca2+ IP3 RECEPTOR. PLC. PIP2. DAG. PKC. + cAMPPK. Ca 2+ CaMKINASE II. TETRAMER (3 ISOFORMS)")
42
INap può contribuire a generare dei PdA con plateau
Le cellule del Purkinje del cervelletto possono generare nel soma PdA con plateau in seguito all’attivazione di conduttanze del Na di tipo persistente su cui si innestano PdA ad alta frequenza

Presentazioni simili

 (Ito) (IKr, IKs) (ICa-L) (INa).>")

















