Scaricare la presentazione
1
BATTERI E VIRUS

2
I BATTERI LE PIÙ ANTICHE FORME DI VITA, COMPARSE SULLA TERRA PIÙ DI 3 MILIARDI DI ANNI FA, SONO CARATTERIZZATE DA UN’ORGANIZZAZIONE CELLULARE PROCARIOTE. TRA ESSE CI SONO I BATTERI

3
LA CELLULA PROCARIOTE I PROCARIOTI HANNO PICCOLE DIMENSIONI (DA 1 A 10 µM) NON PRESENTANO ORGANULI CIRCONDATI DA MEMBRANA POSSIEDONO UNA PARETE CELLULARE CHE LI PROTEGGE DALLA LISI OSMOTICA DISPONGONO DI MATERIALE GENETICO POSTO NEL NUCLEOIDE E COSTITUITO DA UN CROMOSOMA CHE CONTIENE DNA OLTRE AL DNA CROMOSOMICO, LA MAGGIOR PARTE DEI PROCARIOTI CONTIENE ALTRE MOLECOLE DI DNA CHIAMATE PLASMIDI

4
CLASSIFICAZIONE DEI BATTERI
I BATTERI SONO CLASSIFICATI IN DUE DOMINI DERIVATI DA UNA CELLULA ANTENATA COMUNE: QUELLO DEGLI ARCHEA E QUELLO DEGLI EUBACTERIA (*) Cianobatteri, batteri verdi e alcuni batteri purpurei sono fotosintetici
 Cianobatteri, batteri verdi e alcuni batteri purpurei sono fotosintetici.")
5
CARATTERISTICHE DEGLI ARCHEOBATTERI
GLI ARCHEOBATTERI SONO I PROCARIOTI PIÙ ANTICHI E HANNO UNA STRUTTURA PECULIARE CHE LI DISTINGUE DAGLI EUBATTERI: POSSIEDONO ENZIMI CARATTERISTICI HANNO UNA PARETE CELLULARE COMPOSTA DA PROTEINE, POLISACCARIDI O MOLECOLE DI GLICOPEPTIDI L’IMPALCATURA DELLE LORO MEMBRANE CELLULARI È COSTITUITA DA LIPIDI CONTENENTI GLICEROLO E ISOPRENOIDI POSSIEDONO UN PIGMENTO SENSIBILE ALLA LUCE ROSSA, LA ALORODOPSINA (MOLTO SIMILE ALLA RODOPSINA, PRESENTE ANCHE NELLA RETINA DEI VERTEBRATI) POSSONO VIVERE NEGLI AMBIENTI PIÙ INOSPITALI, COME LE SALINE O LE SORGENTI TERMALI
 POSSONO VIVERE NEGLI AMBIENTI PIÙ INOSPITALI, COME LE SALINE O LE SORGENTI TERMALI.")
6
CARATTERISTICHE DEGLI ARCHEOBATTERI
NELLE MEMBRANE CELLULARI DEGLI EUBATTERI E DEGLI ORGANISMI EUCARIOTI SONO PRESENTI FOSFOLIPIDI COSTITUITI DA GLICEROLO UNITO CON LEGAMI ESTERE A DUE ACIDI GRASSI E A UN ACIDO FOSFORICO NEGLI ARCHEOBATTERI IL GLICEROLO È UNITO CON LEGAME ETERE A DUE ISOPRENOIDI. GLI ISOPRENOIDI SONO LIPIDI COSTITUITI DA UNITÀ RIPETUTE DI 5 ATOMI DI CARBONIO (ISOPRENE), CHE FORMANO LUNGHE CATENE SATURE
, CHE FORMANO LUNGHE CATENE SATURE.")
7
Habitat degli archeobatteri
Grotte Lago salato Geyser Saline

8
EUBATTERI: MORFOLOGIA
GLI EUBATTERI HANNO COLONIZZATO UN’ENORME QUANTITÀ DI HABITAT: ACQUE DOLCI E SALATE, REGIONI FREDDE E REGIONI CALDE, TERRENI FANGOSI E SEDIMENTI MARINI. SI TROVANO ANCHE ALL’INTERNO DI ALTRI ORGANISMI COME INSETTI, MOLLUSCHI E MAMMIFERI IN BASE ALLA LORO FORMA SI POSSONO CATALOGARE IN: BACILLI, A BASTONCINO COCCHI, SFERICI SPIRILLI, A ELICA BACILLOCOCCHI, OVALI VIBRIONI, A VIRGOLA

9
EUBATTERI: PARETE LA PARETE CELLULARE DEGLI EUBATTERI CONTIENE UNO SPECIALE COMPOSTO CHIAMATO PEPTIDOGLICANO, UN GLICOPEPTIDE FORMATO DA ACIDO N-ACETIL MURAMICO (NAM) E N- ACETILGLUCOSAMMINA (NAG) UNITI TRA LORO DA PONTI PEPTIDICI A SECONDA DEL TIPO DI PARETE I BATTERI SI DISTINGUONO IN: GRAM POSITIVI E GRAM NEGATIVI
 E N- ACETILGLUCOSAMMINA (NAG) UNITI TRA LORO DA PONTI PEPTIDICI. A SECONDA DEL TIPO DI PARETE I BATTERI SI DISTINGUONO IN: GRAM POSITIVI E GRAM NEGATIVI.")
10
GRAM+ LA PARETE DEI BATTERI GRAM POSITIVI È COSTITUITA PREVALENTEMENTE DA PEPTIDOGLICANO, OLTRE CHE DA ACIDI E PROTEINE

11
GRAM- LA PARETE DEI GRAM NEGATIVI È FORMATA DA DUE STRATI: UN SOTTILE STRATO DI PETIDOGLICANO E UNO STRATO LIPOPROTEICO SIMILE ALLA MEMBRANA PLASMATICA MOLECOLE CARATTERISTICHE DEI GRAM– SONO I LIPOPOLISACCARIDI, DOTATI DI POTERE TOSSICO E PIROGENO

12
COME SI DISTINGUONO I GRAM+ DAI GRAM-
NEL 1884 IL MEDICO DANESE HANS JOACHIM CHRISTIAN GRAM MISE A PUNTO UN METODO DI COLORAZIONE CHE PERMETTE DI DISTINGUERE I BATTERI IN BASE ALLA QUANTITÀ DI PEPTIDOGLICANO CONTENUTO NELLA LORO PARETE CELLULARE GRAM + GRAM -

13
COLORAZIONE DI GRAM I BATTERI CHE RESTANO COLORATI DI VIOLA SONO I GRAM POSITIVI, MENTRE QUELLI CHE NON TRATTENGONO IL COLORANTE E DIVENTANO ROSSI SONO I GRAM NEGATIVI

14
COLORAZIONE DI GRAM

15
COLORAZIONE DI GRAM ECCEZIONE

16
COLORAZIONE DI GRAM ECCEZIONE: MYCOBACTERIUM SPP. PARETE CELLULARE

17
COLORAZIONE DI ZIEHL NEELSEN MYCOBACTERIUM SPP.

18
SPORE QUANDO LE CONDIZIONI AMBIENTALI DIVENTANO SFAVOREVOLI, MOLTI BATTERI GRAM+ SONO IN GRADO DI RICOPRIRE LA PROPRIA ZONA NUCLEARE CON UNA STRUTTURA PARTICOLARMENTE RESISTENTE, FORMANDO UNA SPORA LE SPORE POSSONO RESISTERE PER CENTINAIA DI ANNI IN UNO STATO METABOLICAMENTE INATTIVO. QUANDO LE CONDIZIONI RITORNANO OTTIMALI LA SPORA SI DIFFERENZIA IN CELLULA BATTERICA E IL CICLO VITALE RICOMINCIA

19
I BATTERI SONO MOBILI

20
I BATTERI POSSONO ESSERE:
chemieterotrofi → decompositori → eterotrofi → → patogeni fotoeterotrofi → utili chemioautotofri → autotrofi autotrofi → fotoautotrofi

21
SIMBIOSI BATTERICA MUTUALISMO COMMENSALISMO PARASSITIVISMO

22
BATTERI SIMBIONTI DELL’UOMO
NELL’INTESTINO UMANO VIVONO CIRCA DIFFERENTI SPECIE DI BATTERI (LA “FLORA BATTERICA INTESTINALE”) SIA ANAEROBI (COME BIFIDOBACTERIUM) SIA AEROBI (COME L’ESCHERICHIA E L’ENTEROBACTER) LA FLORA BATTERICA INTESTINALE SVOLGE IMPORTANTI FUNZIONI: PROTEGGE L’ORGANISMO DALL’ATTACCO DEI MICRORGANISMI NOCIVI SCINDE LE SOSTANZE ALIMENTARI SFUGGITE ALLA DIGESTIONE E ALL’ASSORBIMENTO NELL’INTESTINO TENUE, RENDENDO DISPONIBILI PER IL NOSTRO ORGANISMO VITAMINE, SALI MINERALI E ALTRI MICRONUTRIENTI PRODUCE LA VITAMINA K, IMPORTANTE PER LA CORRETTA COAGULAZIONE DEL SANGUE, IL BUON FUNZIONAMENTO DEL FEGATO E LA CALCIFICAZIONE DELLE OSSA PRODUCE LA VITAMINA B12, IMPORTANTE PER LA RIPRODUZIONE DELLE CELLULE E PER LA SINTESI DELL’EMOGLOBINA
 SIA ANAEROBI (COME BIFIDOBACTERIUM) SIA AEROBI (COME L’ESCHERICHIA E L’ENTEROBACTER) LA FLORA BATTERICA INTESTINALE SVOLGE IMPORTANTI FUNZIONI: PROTEGGE L’ORGANISMO DALL’ATTACCO DEI MICRORGANISMI NOCIVI. SCINDE LE SOSTANZE ALIMENTARI SFUGGITE ALLA DIGESTIONE E ALL’ASSORBIMENTO NELL’INTESTINO TENUE, RENDENDO DISPONIBILI PER IL NOSTRO ORGANISMO VITAMINE, SALI MINERALI E ALTRI MICRONUTRIENTI. PRODUCE LA VITAMINA K, IMPORTANTE PER LA CORRETTA COAGULAZIONE DEL SANGUE, IL BUON FUNZIONAMENTO DEL FEGATO E LA CALCIFICAZIONE DELLE OSSA. PRODUCE LA VITAMINA B12, IMPORTANTE PER LA RIPRODUZIONE DELLE CELLULE E PER LA SINTESI DELL’EMOGLOBINA.")
23
ANTIBIOTICI NEL 1929 ALEXANDER FLEMING, UN MEDICO SCOZZESE RICERCATORE PRESSO IL ST. MARY'S HOSPITAL DI LONDRA, SCOPRÌ LA PENICILLINA: LA PRIMA SOSTANZA CONOSCIUTA E STUDIATA CAPACE DI COMBATTERE I BATTERI E DI SCONFIGGERE MALATTIE PRECEDENTEMENTE MORTALI. LA PENICILLINA FU IL CAPOSTIPITE DI UNA FAMIGLIA DI FARMACI AD AZIONE ANTIBIOTICA Attualmente con il termine antibiotico si indica un farmaco, di origine naturale o di sintesi, in grado di rallentare o fermare la proliferazione dei batteri Gli antibiotici si distinguono in: Le penicilline sono antibiotici beta-lattamici isolati da prodotti del metabolismo di alcune specie di Penicillium, in particolare Penicillium notatum oggi noto come Penicillium chrysogenum. Le penicilline sono attive contro la maggior parte dei batteri gram positivi come gli stafilococchi e glistreptococchi, contro le spirochete (Treponema pallidum e Leptospira), contro gonococchi e meningococchi. Nonostante le penicilline siano antibiotici battericidi, a causa dell'intenso uso che se ne è fatto e alla produzione di particolari enzimi (penicillinasi o β-lattamasi) da parte dei batteri, spesso si incorre in fenomeni diresistenza batterica. Le penicilline naturali sono inattive nei confronti del batterio della tubercolosi e dei batteri Gram negativi. Le penicilline semi-sintetiche, prodotte da colture biologiche di Penicillium, hanno in comune un sistema beta-lattamico-diazolico e hanno un ampio spettro di azione, anche contro alcuni batteri gram negativi. Le penicilline sono disponibili sotto diverse preparazioni farmaceutiche: preparazioni somministrabili per via orale (sotto forma di capsule, compresse, sciroppi, polvere solubile, gocce), per via parenterale (sospensioni sterili liofilizzate in flaconi da ricostituire e somministrare poi per via sottocutanea, intramuscolare, endovenosa) ma anche supposte e ovuli, ecc., preparazioni somministrabili per via topica, come creme, pomate, unguenti, colliri, pomate oftalmiche. Alexander Fleming, nel 1928, era rimasto assente dal suo laboratorio per una breve vacanza e stava lavorando su alcuni ceppi di batteri, coltivati in una capsula di coltura. Al ritorno dalla vacanza, Fleming notò che in una capsula c'era un alone chiaro inusuale: in quella zona i batteri non erano cresciuti. Al centro dell'area più chiara, c'era una muffa che aveva contaminato le colture. Fleming pensò che la muffa fosse la causa della morte dei batteri. Le prime sperimentazioni della penicillina sull'uomo non ebbero grandi risultati; bisognava renderla più efficace. Con lo scoppio della seconda guerra mondiale cresceva infatti la richiesta di un antibiotico per le truppe e i civili colpiti dalle infezioni. Si pensò di far arrivare campioni di muffe Penicillum, partendo dal presupposto che alcune producevano penicillina e altre meno, per trovare quella capace di produrre la maggior quantità di antibiotico. La muffa migliore fu scoperta da Mary Hunt su un melone comprato in un supermercato. Tale muffa, battezzata "muffa-Mary", aumentò di oltre dieci volte le capacità produttive e lanciò definitivamente la produzione su larga scala della penicillina. Le penicillina è un estratto dei funghi Penicillium notatum e Penicillium chrysogenum. Sono estratti beta-lattamici che contengono cioè un anello beta-lattamico (deriva dalla condensazione di una funzione carbossilica con l'ammina sul Cβ). La base è l'acido 6-amino penicillanico. L'anello beta-lattamico è condensato con un anello tiazolidinico (è a 5 termini con S e N) e prende nome di nucleo penam. batteriostatici (che bloccano la riproduzione del batterio, impedendone la scissione) battericidi (che uccidono il microrganismo) La penicillina impedisce l’azione dell’enzima che catalizza la formazione dei legami trasversali durante la sintesi del peptidoglicano costituente la parete batterica: senza parete i batteri vanno incontro alla lisi osmotica e muoiono
, contro gonococchi e meningococchi. Nonostante le penicilline siano antibiotici battericidi, a causa dell intenso uso che se ne è fatto e alla produzione di particolari enzimi (penicillinasi o β-lattamasi) da parte dei batteri, spesso si incorre in fenomeni diresistenza batterica. Le penicilline naturali sono inattive nei confronti del batterio della tubercolosi e dei batteri Gram negativi. Le penicilline semi-sintetiche, prodotte da colture biologiche di Penicillium, hanno in comune un sistema beta-lattamico-diazolico e hanno un ampio spettro di azione, anche contro alcuni batteri gram negativi. Le penicilline sono disponibili sotto diverse preparazioni farmaceutiche: preparazioni somministrabili per via orale (sotto forma di capsule, compresse, sciroppi, polvere solubile, gocce), per via parenterale (sospensioni sterili liofilizzate in flaconi da ricostituire e somministrare poi per via sottocutanea, intramuscolare, endovenosa) ma anche supposte e ovuli, ecc., preparazioni somministrabili per via topica, come creme, pomate, unguenti, colliri, pomate oftalmiche. Alexander Fleming, nel 1928, era rimasto assente dal suo laboratorio per una breve vacanza e stava lavorando su alcuni ceppi di batteri, coltivati in una capsula di coltura. Al ritorno dalla vacanza, Fleming notò che in una capsula c era un alone chiaro inusuale: in quella zona i batteri non erano cresciuti. Al centro dell area più chiara, c era una muffa che aveva contaminato le colture. Fleming pensò che la muffa fosse la causa della morte dei batteri. Le prime sperimentazioni della penicillina sull uomo non ebbero grandi risultati; bisognava renderla più efficace. Con lo scoppio della seconda guerra mondiale cresceva infatti la richiesta di un antibiotico per le truppe e i civili colpiti dalle infezioni. Si pensò di far arrivare campioni di muffe Penicillum, partendo dal presupposto che alcune producevano penicillina e altre meno, per trovare quella capace di produrre la maggior quantità di antibiotico. La muffa migliore fu scoperta da Mary Hunt su un melone comprato in un supermercato. Tale muffa, battezzata muffa-Mary , aumentò di oltre dieci volte le capacità produttive e lanciò definitivamente la produzione su larga scala della penicillina. Le penicillina è un estratto dei funghi Penicillium notatum e Penicillium chrysogenum. Sono estratti beta-lattamici che contengono cioè un anello beta-lattamico (deriva dalla condensazione di una funzione carbossilica con l ammina sul Cβ). La base è l acido 6-amino penicillanico. L anello beta-lattamico è condensato con un anello tiazolidinico (è a 5 termini con S e N) e prende nome di nucleo penam. batteriostatici (che bloccano la riproduzione del batterio, impedendone la scissione) battericidi (che uccidono il microrganismo) La penicillina impedisce l’azione dell’enzima che catalizza la formazione dei legami trasversali durante la sintesi del peptidoglicano costituente la parete batterica: senza parete i batteri vanno incontro alla lisi osmotica e muoiono.")
24
RIPRODUZIONE BATTERICA
ASSESSUATA SESSUATA Scissione binaria Gemmazione Frammentazione (actinomiceti) Coniugazione Trasformazione Trasduzione
 Coniugazione. Trasformazione. Trasduzione.")
25
RIPRODUZIONE BATTERICA ASESSUATA SCISSIONE BINARIA
LA DIVISIONE DI UN PROCARIOTE COMPORTA LA DUPLICAZIONE DEL CROMOSOMA BATTERICO E LA MIGRAZIONE DELLE DUE MOLECOLE AI LATI OPPOSTI DELLA MEMBRANA: QUANDO LA CELLULA SI ALLUNGA, I CROMOSOMI SI TROVANO ANCORATI ALLA MEMBRANA DA PARTI OPPOSTE AL TERMINE DEL PROCESSO DI ALLUNGAMENTO LA MEMBRANA E LA PARETE SI RIPIEGANO AL CENTRO FORMANDO DUE CELLULE FIGLIE PERFETTAMENTE IDENTICHE ALLA CELLULA MADRE Manca il fuso mitotico, ma si forma un apparato mitotico primordiale nel quale risulta centrale la funzione della membrana citoplasmatica (mesosomi settali).
.")
26
RIPRODUZIONE BATTERICA ASESSUATA GEMMAZIONE
Nella gemmazione, si ha una normale divisione del nucleo della cellula madre per mitosi, che a causa di una distribuzione ineguale di citoplasma durante la citocinesi, da origine ad una cellula figlia più piccola. La cellula più piccola può subito separarsi dalla cellula madre o rimanere per qualche tempo legata ad essa ed andare incontro ad ulteriori divisioni

27
RIPRODUZIONE BATTERICA ASESSUATA FRAMMENTAZIONE
Propria degli ACTINOMICETI Le pareti cellulari si accrescono all’interno della cellula che viene scissa in numerose cellule di nuova costituzione Sono sporigeni e la loro riproduzione è singolare, avviene per frammentazione dei filamenti in elementi coccoidi o bacillari, a ognuno dei quali prende origine un nuovo micelio, in alcuni casi si ha la produzione di vere e proprie artrospore o spore simili ai conidi di miceti. Hanno questo nome per la loro caratteristica disposizione nelle lesioni tissutali a raggiera. Nonostante le numerose somiglianze con i miceti, sono batteri a tutti gli effetti, per la struttura procariote, il peptidoglicano della parete cellulare, la sensibilità ad alcuni antibiotici, ricombinazione genetica (merozigoti). Si dividono in: Actinomyces: anaerobi obbligati o microaerofili, producono un micelio ramificato e si riproducono per frammentazione, hanno aspetto bacillare simile ai corinebatteri. Nocardia: aerobi, producono un esteso micelio vegetativo, sono acido-resistenti e si riproducono per frammentazione. Streptomyces: aerobi, producono un esteso micelio vegetativo, produce spore simili a conidi, sono normalmente saprofiti del suolo e hanno la capacità di produrre antibiotici. - See more at:
. Si dividono in: Actinomyces: anaerobi obbligati o microaerofili, producono un micelio ramificato e si riproducono per frammentazione, hanno aspetto bacillare simile ai corinebatteri. Nocardia: aerobi, producono un esteso micelio vegetativo, sono acido-resistenti e si riproducono per frammentazione. Streptomyces: aerobi, producono un esteso micelio vegetativo, produce spore simili a conidi, sono normalmente saprofiti del suolo e hanno la capacità di produrre antibiotici. - See more at:")
28
RIPRODUZIONE BATTERICA SESSUATA
LA CONIUGAZIONE, OVVERO IL TRASFERIMENTO DIRETTO DI DNA DA UN BATTERIO ALL’ALTRO MEDIANTE PLASMIDI LA TRASFORMAZIONE, MEDIANTE CUI UNA CELLULA CATTURA FRAMMENTI DI DNA DALL’AMBIENTE CIRCOSTANTE LA TRASDUZIONE, CIOÈ IL PASSAGGIO TRA CELLULE DI MATERIALE GENETICO BATTERICO MEDIANTE VIRUS

29
CONIUGAZIONE LA CONIUGAZIONE È IL PROCESSO IN CUI IL DNA DEL PLASMIDE È TRASFERITO DA UN BATTERIO DONATORE A UN RICEVENTE ATTRAVERSO UN PONTE CITOPLASMATICO, FORMATO DA APPENDICI DETTE PILI. LE CELLULE F- (RICEVENTI) SONO PRIVE DI PLASMIDE, QUELLE F+ (DONATRICI) LO CONTENGONO
 SONO PRIVE DI PLASMIDE, QUELLE F+ (DONATRICI) LO CONTENGONO.")
30
TRASFORMAZIONE LA TRASFORMAZIONE SI VERIFICA QUANDO UN BATTERIO CATTURA DALL’AMBIENTE EXTRACELLULARE UN FILAMENTO DI DNA APPARTENUTO A UN ALTRO BATTERIO MORTO IL DNA ESTRANEO SI INSERISCE NEL CROMOSOMA BATTERICO MODIFICANDONE IL PATRIMONIO GENETICO POSSONO COMPIERE LA TRASFORMAZIONE SOLO QUEI BATTERI CHE POSSEGGONO SPECIALI PROTEINE, DETTE FATTORI DI COMPETENZA, CHE FACILITANO LA CATTURA, L’INGRESSO E L’INSERIMENTO DEL DNA ESTRANEO

31
TRASDUZIONE LA TRASDUZIONE AVVIENE QUANDO UN VIRUS, CHE HA INFETTATO UN BATTERIO E HA ACQUISITO PARTE DEL SUO GENOMA, PENETRA IN UN SECONDO BATTERIO INSERENDOVI IL MATERIALE GENETICO DERIVANTE DAL PRIMO BATTERIO INFETTATO

32
I VIRUS Virus del Mosaico del tabacco DMITRI IVANOWSKY 1892
MARTINUS BEIJERINCK 1898 Adenovirus (virus dell’influenza)
")
33
ALTRI ESEMPI DI VIRUS Batteriofago T4

34
IDENTIFICAZIONE

35
RIVESTIMENTO PROTEICO (FINO AL 90% DEL VIRIONE)
CAPSIDE RIVESTIMENTO PROTEICO (FINO AL 90% DEL VIRIONE) FORNISCE PROTEZIONE ALL'ACIDO NUCLEICO È IMPORTANTE PER L’INFETTIVITÀ (STRUTTURE ANTI-RECETTORIALI) DAL MOMENTO CHE IL GENOMA VIRALE HA UN NUMERO LIMITATO DI GENI, IL CAPSIDE È COSTITUITO DALLA ASSOCIAZIONE DI SUBUNITÀ PROTEICHE IN PROTOMERI RIPETUTI (CAPSOMERI) COLLEGATI DA LEGAMI TRA GRUPPI CHIMICI PRESENTI SULLA SUPERFICIE E AUTO ASSEMBLANTI
 FORNISCE PROTEZIONE ALL ACIDO NUCLEICO. È IMPORTANTE PER L’INFETTIVITÀ (STRUTTURE ANTI-RECETTORIALI) DAL MOMENTO CHE IL GENOMA VIRALE HA UN NUMERO LIMITATO DI GENI, IL CAPSIDE È COSTITUITO DALLA ASSOCIAZIONE DI SUBUNITÀ PROTEICHE IN PROTOMERI RIPETUTI (CAPSOMERI) COLLEGATI DA LEGAMI TRA GRUPPI CHIMICI PRESENTI SULLA SUPERFICIE E AUTO ASSEMBLANTI.")
36
CAPSIDE IL CAPSIDE PUÒ:
ESSERE COSTITUITO DA UN’UNICA PROTEINA CHE SI RIPETE PIÙ VOLTE O DA PROTEINE DIVERSE AVERE SULLA SUPERFICIE GLICOPROTEINE CHE SERVONO A RICONOSCERE LE CELLULE BERSAGLIO LE FORME PIÙ COMUNI SONO: ELICOIDALE, COME QUELLA DEL MOSAICO DEL TABACCO ICOSAEDRICA, COME QUELLA DEL VIRUS DEL RAFFREDDORE

37
PERICAPSIDE IL CAPSIDE PUÒ ESSERE AVVOLTO DA UN RIVESTIMENTO LIPOPROTEICO, DETTO PERICAPSIDE (O ENVELOPE), CHE DERIVA DALLA MEMBRANA DELLA CELLULA OSPITE ED È COMPOSTO DA UN DOPPIO STRATO DI FOSFOLIPIDI INTERVALLATI DA NUMEROSE GLICOPROTEINE
, CHE DERIVA DALLA MEMBRANA DELLA CELLULA OSPITE ED È COMPOSTO DA UN DOPPIO STRATO DI FOSFOLIPIDI INTERVALLATI DA NUMEROSE GLICOPROTEINE.")
38
GENOMA VIRALE IL GENOMA VIRALE CODIFICA LE PROTEINE DEL CAPSIDE E UNO O PIÙ ENZIMI CHE CATALIZZANO LA SUA DUPLICAZIONE; IL GENOMA VIRALE PUÒ ESSERE COSTITUITO DA DNA OPPURE DA RNA A DOPPIO FILAMENTO O A FILAMENTO SINGOLO CON DISPOSIZIONE LINEARE O CIRCOLARE

39
CLASSIFICAZIONE DI BALTIMORE
La Classificazione di Baltimore è una classificazione dei virus, proposta dal biologo David Baltimore premio Nobel per la medicina nel1975, basato sulla natura e sulla polarità dei genomi virali. I vari gruppi di virus vengono suddivisi in famiglie a seconda della natura del loro genoma (sia esso DNA, RNA, a singolo a doppio filamento) e dal loro tipo di replicazione.
 e dal loro tipo di replicazione.")
40
LA FUNZIONE DEI RIVESTIMENTI ESTERNI
1) PROTEZIONE DEL FRAGILE GENOMA DA DANNI FISICI, CHIMICI, ENZIMATICI. I CAPSIDI SONO FORMATI DA UN CERTO NUMERO DI UNITÀ PROTEICHE UGUALI. IL DANNO AD UNA RENDE QUELL’UNA NON FUNZIONALE, MA DIFFICILMENTE DANNEGGIA L’INTERA PARTICELLA. QUESTO FA DEL CAPSIDE UN’EFFICACE BARRIERA. 2)CONFERIMENTO DELLA CAPACITÀDI RICONOSCIMENTO CON IL RECETTORE CELLULARE.
 PROTEZIONE DEL FRAGILE GENOMA DA DANNI FISICI, CHIMICI, ENZIMATICI. I CAPSIDI SONO FORMATI DA UN CERTO NUMERO DI UNITÀ PROTEICHE UGUALI. IL DANNO AD UNA RENDE QUELL’UNA NON FUNZIONALE, MA DIFFICILMENTE DANNEGGIA L’INTERA PARTICELLA. QUESTO FA DEL CAPSIDE UN’EFFICACE BARRIERA. 2)CONFERIMENTO DELLA CAPACITÀDI RICONOSCIMENTO CON IL RECETTORE CELLULARE.")
41
LA FUNZIONE DEI RIVESTIMENTI ESTERNI
3)PENETRAZIONE DEL GENOMA VIRALE NELLA CELLULA IN UNA CONFORMAZIONE CON LA QUALE PUÒ INTERAGIRE CON LE STRUTTURE CELLULARI ED INIZIARE IL PROCESSO INFETTIVO - IN ALCUNI CASI QUESTO È UN PROCESSO FACILE CHE CONSISTE SEMPLICEMENTE NEL FAR ENTRARE IL GENOMA NEL CITOPLASMA -IN ALTRI CASI INVECE, QUESTO MOMENTO È PIÙ COMPLESSO. PER ES. I RETROVIRUS
PENETRAZIONE DEL GENOMA VIRALE NELLA CELLULA IN UNA CONFORMAZIONE CON LA QUALE PUÒ INTERAGIRE CON LE STRUTTURE CELLULARI ED INIZIARE IL PROCESSO INFETTIVO. - IN ALCUNI CASI QUESTO È UN PROCESSO FACILE CHE CONSISTE SEMPLICEMENTE NEL FAR ENTRARE IL GENOMA NEL CITOPLASMA. -IN ALTRI CASI INVECE, QUESTO MOMENTO È PIÙ COMPLESSO. PER ES. I RETROVIRUS.")
42
CLASSIFICAZIONE DEI VIRUS
I VIRUS SI POSSONO DISTINGUERE IN BASE ALL’ORGANISMO NEL QUALE SI RIPRODUCONO IN: BATTERIOFAGI (O FAGI), CHE INFETTANO I BATTERI VIRUS VEGETALI, CHE INFETTANO LE CELLULE DELLE PIANTE (COME IL VIRUS DEL MOSAICO DEL TABACCO) VIRUS ANIMALI, CHE INFETTANO LE CELLULE ANIMALI (COME HERPES VIRUS CHE PROVOCA LA VARICELLA)
, CHE INFETTANO I BATTERI. VIRUS VEGETALI, CHE INFETTANO LE CELLULE DELLE PIANTE (COME IL VIRUS DEL MOSAICO DEL TABACCO) VIRUS ANIMALI, CHE INFETTANO LE CELLULE ANIMALI (COME HERPES VIRUS CHE PROVOCA LA VARICELLA)")
43
Fusione della membrana
INGRESSO DEI VIRUS Fusione della membrana

44
INGRESSO DEI VIRUS ENDOCITOSI

45
CICLO LITICO E LISOGENO
I PRINCIPALI GRUPPI VIRALI UTILIZZANO MECCANISMI DI RIPRODUZIONE SPECIFICI CHE SI POSSONO RICONDURRE A DUE MODALITÀ PRINCIPALI: IL CICLO LITICO IL CICLO LISOGENO

46
CICLO LITICO NEL CICLO LITICO LA CELLULA OSPITE DIVENTA UNA “FABBRICA” DI VIRUS: DOPO L’INGRESSO NELLA CELLULA OSPITE, L’ACIDO NUCLEICO E LE PROTEINE VIRALI SONO PRODOTTI RAPIDAMENTE SFRUTTANDO LE STRUTTURE E L’ENERGIA DELLA CELLULA LA CELLULA OSPITE SI ROMPE E LE PARTICELLE VIRALI SONO LIBERATE ALL’ESTERNO

47
CICLO LISOGENO NEL CICLO LISOGENO IL GENOMA VIRALE SI INTEGRA NEL CROMOSOMA OSPITE IL GENOMA VIRALE SI DUPLICA DURANTE IL PROCESSO DI DUPLICAZIONE DELLA CELLULA OSPITE. QUANDO QUESTA SI DIVIDE, QUINDI, TRASMETTE ALLE CELLULE FIGLIE ANCHE IL DNA VIRALE (CHE PERÒ NON È ESPRESSO) DOPO MIGLIAIA DI DIVISIONI CELLULARI, IL DNA VIRALE PUÒ ATTIVARSI, STACCANDOSI DAL CROMOSOMA E DANDO INIZIO A UN CICLO LITICO Il genoma virale integrato è chiamato profago, se la cellula ospite è un batterio, o provirus, se la cellula ospite è eucariote
 DOPO MIGLIAIA DI DIVISIONI CELLULARI, IL DNA VIRALE PUÒ ATTIVARSI, STACCANDOSI DAL CROMOSOMA E DANDO INIZIO A UN CICLO LITICO. Il genoma virale integrato è chiamato profago, se la cellula ospite è un batterio, o provirus, se la cellula ospite è eucariote.")
48
FASI DEL CICLO VIRALE ENTRAMBI I CICLI RIPRODUTTIVI SONO CARATTERIZZATI DA CINQUE FASI: ATTACCO PENETRAZIONE REPLICAZIONE MATURAZIONE LIBERAZIONE

49
CICLO RIPRODUTTIVO DEI VIRUS: ATTACCO
L’ATTACCO (O ADSORBIMENTO) CONSISTE NELLA FORMAZIONE DI UN LEGAME SPECIFICO TRA MOLECOLE (IN GENERE GLICOPROTEINE) PRESENTI SULLA SUPERFICIE VIRALE E MOLECOLE PROTEICHE RECETTRICI PRESENTI SULLA SUPERFICIE DELLA CELLULA OSPITE IN TAL MODO SOLO ALCUNI TIPI DI CELLULE VENGONO INFETTATE DA UN PARTICOLARE VIRUS: IL VIRUS DEL RAFFREDDORE, PER ESEMPIO, ATTACCA SOLO LE CELLULE DELLA MUCOSA NASALE
 CONSISTE NELLA FORMAZIONE DI UN LEGAME SPECIFICO TRA MOLECOLE (IN GENERE GLICOPROTEINE) PRESENTI SULLA SUPERFICIE VIRALE E MOLECOLE PROTEICHE RECETTRICI PRESENTI SULLA SUPERFICIE DELLA CELLULA OSPITE. IN TAL MODO SOLO ALCUNI TIPI DI CELLULE VENGONO INFETTATE DA UN PARTICOLARE VIRUS: IL VIRUS DEL RAFFREDDORE, PER ESEMPIO, ATTACCA SOLO LE CELLULE DELLA MUCOSA NASALE.")
50
CICLO RIPRODUTTIVO DEI VIRUS: PENETRAZIONE
QUASI TUTTI I VIRUS UMANI ENTRANO NELLA CELLULA CON IL CAPSIDE INTATTO IN PRESENZA DEL PERICAPSIDE, QUESTO SI FONDE CON LA MEMBRANA CELLULARE; ALTRIMENTI IL VIRUS PUÒ ENTRARE PER ENDOCITOSI

51
CICLO RIPRODUTTIVO DEI VIRUS: REPLICAZIONE
LA REPLICAZIONE CONSISTE NELLA DUPLICAZIONE DEL MATERIALE GENETICO VIRALE UTILIZZANDO LE RISORSE ENERGETICHE E GLI ENZIMI DELLA CELLULA OSPITE LA REPLICAZIONE DELL’ACIDO NUCLEICO PUÒ AVVENIRE CON IL MATERIALE GENETICO VIRALE SEPARATO DA QUELLO DELLA CELLULA OSPITE (COME SI VERIFICA NEL CICLO LITICO) O INSERITO NEL DNA DELLA CELLULA OSPITE (COME SI VERIFICA NEL CICLO LISOGENO)
 O INSERITO NEL DNA DELLA CELLULA OSPITE (COME SI VERIFICA NEL CICLO LISOGENO)")
52
CICLO RIPRODUTTIVO DEI VIRUS: MATURAZIONE
DURANTE LA MATURAZIONE SONO SINTETIZZATI I COMPONENTI STRUTTURALI DEL VIRUS (CAPSIDE, ENZIMI VIRALI) NEL CICLO LITICO QUESTA SINTESI AVVIENE SUBITO DOPO L’INFEZIONE, MENTRE NEL CICLO LISOGENO IL VIRUS PUÒ RIMANERE “LATENTE” (DUPLICARSI SENZA PRODURRE NUOVE COMPONENTI STRUTTURALI DEL VIRUS) ANCHE A LUNGO
 NEL CICLO LITICO QUESTA SINTESI AVVIENE SUBITO DOPO L’INFEZIONE, MENTRE NEL CICLO LISOGENO IL VIRUS PUÒ RIMANERE LATENTE (DUPLICARSI SENZA PRODURRE NUOVE COMPONENTI STRUTTURALI DEL VIRUS) ANCHE A LUNGO.")
53
CICLO RIPRODUTTIVO DEI VIRUS: LIBERAZIONE
LA LIBERAZIONE DEI VIRUS DALLA CELLULA PUÒ AVVENIRE PER GEMMAZIONE O PER LISI CELLULARE I VIRUS CARATTERIZZATI DALLA PRESENZA DEL PERICAPSIDE FUORIESCONO PER GEMMAZIONE (ES. VIRUS EBOLA), I VIRUS SENZA PERICAPSIDE PER LISI CELLULARE Il virus infatti risulta in forma attiva soltanto all’interno della cellula ospite, mentre quando si trova all’esterno è come inerte e prende il nome di virione. Il genoma si trova isolato all’interno della “testa” del virus in una struttura che prende il nome di capside. Si conoscono virus diversi per tipologia e forma: fra i più studiati e usati in laboratorio come prototipo modello dei meccanismi virali vi sono i batteriofagi, o semplicemente fagi. Questi virus sono in grado di infettare cellule batteriche in maniera quasi totalmente esclusiva. I fagi, come tutti i virus, subiscono mutazioni casuali nel proprio corredo genomico e di conseguenza possono diventare in grado di attaccare anche cellule eucariotiche. I virus si replicano con due principali meccanismi alternativi: il ciclo litico e il ciclo lisogeno. Entrambi porteranno alla lisi della cellule ospite, ovvero alla sua degradazione, ma con modalità e tempistiche diverse. Il ciclo litico consiste nell’iniezione da parte del virus del proprio corredo genetico all’interno dell’ospite (1). Ciò è possibile grazie alle “zampette” proteiche di cui il fago dispone e che gli permettono l’aggancio alla cellula. Una volta all’interno, il genoma del virus viene replicato in modo da essere un doppio filamento (sempre che non lo fosse già in partenza) e va a costituire un plasmide (2), ovvero una molecola di DNA circolare. Nel caso in cui il genoma del virus sia ad RNA, esso verrà retrotrascritto a DNA a doppio filamento, che rappresenta la forma più stabile in cui il genoma del fago litico può trovarsi nell’ospite: in caso contrario gli enzimi in grado di digerire gli acidi nucleici (nucleasi) saranno in grado di eliminare il virus più facilmente. Il plasmide virale viene quindi replicato ma soprattutto trascritto e tradotto automaticamente dall’ospite, in quanto gli apparati trascrizionale e traduzionale non sono in grado di discriminare il proprio genoma da quello virale. Il genoma virale contiene i geni che codificano per le proteine che costituiscono i virus stesso, le quali verrano sintetizzate (3) e successivamente assemblate fra loro nella maniera corretta (4). In seguito i capsidi neosintetizzati incorporano al loro interno un singola copia del genoma virale(5), replicatosi sempre nella cellula. Una volta che i virus completi raggiungono un certo numero la cellula ospite subisce una lisi(6), cioè le sue pareti si rompono liberando una grande quantità di virus, ora virioni, all’esterno, pronti ad infettare le cellule vicine. Ciclo litico e lisogeno di un virus all’interno di una cellula ospite Il ciclo lisogeno, invece, differisce dal precedente in quanto il genoma virale non resta separato da quello dell’ospite ma è in grado di integrarsi al suo interno, mediante l’interazione con particolare sequenze genetiche che ne permettono inserzione (7). Il genoma virale in questa forma prende il nome di profago, e viene replicato insieme al resto del corredo genetico dell’ospite, risultando “invisibile” ad esso (8). In questo modo ogni cellula figlia della cellula originariamente infettata presenta il profago (9), il quale rimane in questa condizione di inattività fino a quando stimoli esterni non ne possono determinare l’uscita (escissione), alla quale segue la lisi della cellula con un meccanismo del tutto analogo a quello del ciclo litico. Fuoriuscita per gemmazione
, I VIRUS SENZA PERICAPSIDE PER LISI CELLULARE. Il virus infatti risulta in forma attiva soltanto all’interno della cellula ospite, mentre quando si trova all’esterno è come inerte e prende il nome di virione. Il genoma si trova isolato all’interno della testa del virus in una struttura che prende il nome di capside. Si conoscono virus diversi per tipologia e forma: fra i più studiati e usati in laboratorio come prototipo modello dei meccanismi virali vi sono i batteriofagi, o semplicemente fagi. Questi virus sono in grado di infettare cellule batteriche in maniera quasi totalmente esclusiva. I fagi, come tutti i virus, subiscono mutazioni casuali nel proprio corredo genomico e di conseguenza possono diventare in grado di attaccare anche cellule eucariotiche. I virus si replicano con due principali meccanismi alternativi: il ciclo litico e il ciclo lisogeno. Entrambi porteranno alla lisi della cellule ospite, ovvero alla sua degradazione, ma con modalità e tempistiche diverse. Il ciclo litico consiste nell’iniezione da parte del virus del proprio corredo genetico all’interno dell’ospite (1). Ciò è possibile grazie alle zampette proteiche di cui il fago dispone e che gli permettono l’aggancio alla cellula. Una volta all’interno, il genoma del virus viene replicato in modo da essere un doppio filamento (sempre che non lo fosse già in partenza) e va a costituire un plasmide (2), ovvero una molecola di DNA circolare. Nel caso in cui il genoma del virus sia ad RNA, esso verrà retrotrascritto a DNA a doppio filamento, che rappresenta la forma più stabile in cui il genoma del fago litico può trovarsi nell’ospite: in caso contrario gli enzimi in grado di digerire gli acidi nucleici (nucleasi) saranno in grado di eliminare il virus più facilmente. Il plasmide virale viene quindi replicato ma soprattutto trascritto e tradotto automaticamente dall’ospite, in quanto gli apparati trascrizionale e traduzionale non sono in grado di discriminare il proprio genoma da quello virale. Il genoma virale contiene i geni che codificano per le proteine che costituiscono i virus stesso, le quali verrano sintetizzate (3) e successivamente assemblate fra loro nella maniera corretta (4). In seguito i capsidi neosintetizzati incorporano al loro interno un singola copia del genoma virale(5), replicatosi sempre nella cellula. Una volta che i virus completi raggiungono un certo numero la cellula ospite subisce una lisi(6), cioè le sue pareti si rompono liberando una grande quantità di virus, ora virioni, all’esterno, pronti ad infettare le cellule vicine. Ciclo litico e lisogeno di un virus all’interno di una cellula ospite Il ciclo lisogeno, invece, differisce dal precedente in quanto il genoma virale non resta separato da quello dell’ospite ma è in grado di integrarsi al suo interno, mediante l’interazione con particolare sequenze genetiche che ne permettono inserzione (7). Il genoma virale in questa forma prende il nome di profago, e viene replicato insieme al resto del corredo genetico dell’ospite, risultando invisibile ad esso (8). In questo modo ogni cellula figlia della cellula originariamente infettata presenta il profago (9), il quale rimane in questa condizione di inattività fino a quando stimoli esterni non ne possono determinare l’uscita (escissione), alla quale segue la lisi della cellula con un meccanismo del tutto analogo a quello del ciclo litico. Fuoriuscita per gemmazione.")
54
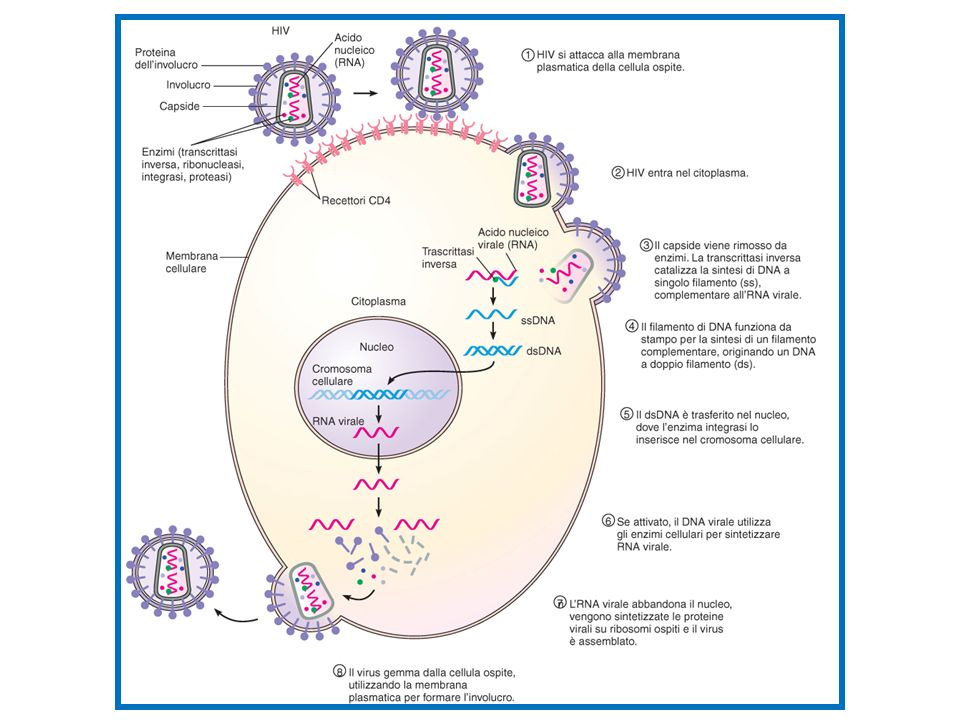
LATENZA I VIRUS CHE INTEGRANO I LORO GENI ALL’INTERNO DEL DNA DELLA CELLULA OSPITE SONO DI DUE TIPI: VIRUS A DNA E RETROVIRUS A RNA UN ESEMPIO DI VIRUS A DNA CHE ALTERNA LUNGHI PERIODI DI LATENZA A BREVI PERIODI RIPRODUTTIVI È L’HERPES SIMPLEX CHE VIENE ATTIVATO DA FATTORI LEGATI A STRESS FISICO O EMOTIVO UN ESEMPIO DI RETROVIRUS A RNA È IL VIRUS HIV CHE PROVOCA LA COSIDDETTA IMMUNODEFICIENZA UMANA, IN QUANTO COLPISCE LE CELLULE IMMUNITARIE (LINFOCITI T)
")
55
RETROVIRUS L’INTEGRAZIONE DI UN VIRUS A RNA IN UN CROMOSOMA COSTITUITO DA DNA PUÒ AVVENIRE GRAZIE A UN IMPORTANTE ENZIMA VIRALE: LA TRASCRITTASI INVERSA LA TRASCRITTASI INVERSA COPIA L’RNA VIRALE IN UN FILAMENTO SINGOLO DI DNA COMPLEMENTARE (CDNA) DAL CDNA SI ORIGINA UNA MOLECOLA DI DNA A DOPPIO FILAMENTO CHE SI INSERISCE NEL CROMOSOMA DELLA CELLULA OSPITE E SI REPLICA CON ESSA
 DAL CDNA SI ORIGINA UNA MOLECOLA DI DNA A DOPPIO FILAMENTO CHE SI INSERISCE NEL CROMOSOMA DELLA CELLULA OSPITE E SI REPLICA CON ESSA.")
57
Penetrazione del virus attraverso lesioni di cute e membrane mucose
Penetrazione del virus attraverso lesioni di cute e membrane mucose. Il virus si riproduce alla base della lesione ed infetta i neuroni innervanti raggiungendo per trasporto assonale retrogrado i gangli: • gangli del trigemino per l’HSV-1 • gangli sacrali per HSV-2 Il virus poi ritorna nel sito iniziale di infezione e può rimanere inapparente o causare lesioni vescicolari. La lesione si rimargina senza lasciare la cicatrice.

59
VIROIDI I viroidi sono i più piccoli agenti patogeni conosciuti e agenti causali di malattie delle piante. La forma extracellulare del viroide consiste semplicemente di molecole di RNA nude, ovvero non racchiuse da un rivestimento proteico protettivo; tali agenti infettanti non codificano infatti neppure per una singola proteina. Un viroide è quindi solo una piccola molecola di RNA circolare arrotolata su se stessa a creare un esteso segmento a doppio filamento Vi sono in natura altri agenti, detti viroidi e prioni, che non sono conformi alla definizione di virus, anche se sono altrettanto piccoli e contagiosi. Esiste una stretta affinità fra virus e viroidi: i viroidi sono infatti molecole di acido nucleico nude composte interamente di RNA circolare. I prioni, d’altro canto, sono entità infettanti a parte: sono costituite soltanto di singole molecole proteiche. Come si riproducono i viroidi? Innanzitutto, poiché non debbono affrontare il problema di sintetizzare le proteine del capside, nel loro genoma non sono codificati i relativi geni. Tuttavia essi devono affrontare, al pari dei virus a RNA, il problema di sintetizzare l’RNA da RNA stampo, e inoltre di come renderlo circolare. Le modalità con cui i viroidi compiono tale sintesi è un evento abbastanza complesso. L’RNA dei viroidi infettanti viene replicato da una RNA polimerasi dell’ospite che può sintetizzare l’RNA usando un RNA stampo. Tali enzimi sono comuni nelle piante ma non lo sono negli animali e nei batteri non infetti. Il processo di replicazione viene attuato usando l’RNA circolare come stampo e formando una copia, complementare ad esso, spostandosi tutto intorno al cerchio; questo meccanismo di replicazione a cerchio rotante viene usato anche da alcuni virus a DNA e dai plasmidi. Il risultato di tale operazione non consiste in una singola copia dello stampo, ma in una lunga serie di copie ripetute in tandem, come un rotolo di carta igienica non svolto. Per produrre viroidi, questa lunga molecola deve essere scissa in segmenti di dimensioni adeguate; questo può essere fatto in uno dei due seguenti modi. In alcuni viroidi, lo stesso RNA presenta un’attività enzimatica, ossia è un ribozima, ed è quindi capace di tagliare la lunga catena in singole copie; in altri viroidi il taglio viene eseguito da una endonucleasi dell’ospite. In entrambi i casi le copie risultanti devono legarsi in cerchi, che poi assumono la struttura molecolare del viroide. Il solo agente infettante umano simile al viroide è detto erroneamente virus dell’epatite D (HDV); in realtà tale particella è qualcosa di intermedio fra un virus e un viroide e quindi un’altra possibile denominazione potrebbe essere quella di “virusoide” L’HDV siste di RNA nudo che differisce dai viroidi delle piante in quanto codifica per proteine. Tuttavia, a differenza dei veri e propri virus, l’agente dell’epatite D non codifica per il proprio capside, mentre le proteine che esso codifica sembrano svolgere un ruolo importante nell’impacchettamento delle particelle. L’agente dell’epatite D utilizza il capside di un virus autentico, ossia il virus dell’epatite B (HBV), per essere impacchettato. Da tutto ciò si evince che la malattia epatite D si verifica solo se l’ospite viene contemporaneamente coinfettato sia dal virus dell’epatite B che dall’agente dell’epatite D. Questo particolare tipo di infezione doppia comporta l’insorgenza di una malattia ben più grave di quella indotta dalla sola infezione da virus dell’epatite B. L’agente dell’epatite D può causare la malattia inattivando un componente essenziale delle cellule: il suo RNA presenta, infatti, un’ampia omologia di sequenza con una particella di RNA 7S citoplasmatica coinvolta nel riconoscimento del segnale e nella traslocazione di proteine secretorie e di quelle associate alla membrana; è probabile che l’RNA del virusoide sequestri o scinda la particella 7S, determinando la morte cellulare.
; in realtà tale particella è qualcosa di intermedio fra un virus e un viroide e quindi un’altra possibile denominazione potrebbe essere quella di virusoide L’HDV siste di RNA nudo che differisce dai viroidi delle piante in quanto codifica per proteine. Tuttavia, a differenza dei veri e propri virus, l’agente dell’epatite D non codifica per il proprio capside, mentre le proteine che esso codifica sembrano svolgere un ruolo importante nell’impacchettamento delle particelle. L’agente dell’epatite D utilizza il capside di un virus autentico, ossia il virus dell’epatite B (HBV), per essere impacchettato. Da tutto ciò si evince che la malattia epatite D si verifica solo se l’ospite viene contemporaneamente coinfettato sia dal virus dell’epatite B che dall’agente dell’epatite D. Questo particolare tipo di infezione doppia comporta l’insorgenza di una malattia ben più grave di quella indotta dalla sola infezione da virus dell’epatite B. L’agente dell’epatite D può causare la malattia inattivando un componente essenziale delle cellule: il suo RNA presenta, infatti, un’ampia omologia di sequenza con una particella di RNA 7S citoplasmatica coinvolta nel riconoscimento del segnale e nella traslocazione di proteine secretorie e di quelle associate alla membrana; è probabile che l’RNA del virusoide sequestri o scinda la particella 7S, determinando la morte cellulare.")
60
PRIONI la più nota patologia degenerativa cerebrale è stata la cosiddetta “malattia della mucca pazza”, ma ci sono anche altre manifestazioni cliniche simili, come la malattia di Creutzfeldt-Jakob (CJD) negli esseri umani e lo scrapie nelle pecore. Queste malattie sono denominate nel loro insieme encefalopatie spongiformi perché formano fori nel cervello, che fanno assumere al tessuto cerebrale un aspetto a spugna. il prione, è altamente insolito in quanto non contiene acidi nucleici, ma è composto interamente da una proteina La natura infettiva di una di queste malattie umane, il kuru, fu ipotizzata quando venne osservata l’associazione fra persone colpite e il loro consumo di carne di cervello umano a seguito di rituali tribali. È stato poi provato che alcune di queste malattie possono essere trasmesse anche in animali da laboratorio. In tutti questi esempi patologici, i ricercatori hanno sospettato che l’agente eziologico potesse essere un virus e tuttavia non riuscivano a isolare alcun agente infettivo conosciuto (virus, batteri o altri). In seguito, fu chiarito che l’agente infettivo, poi chiamato “prione”, è altamente insolito in quanto non contiene acidi nucleici, ma è composto interamente da una proteina. Ciò risulta sorprendente perché tutti gli altri agenti infettivi, ovvero tutte le entità in grado di replicarsi, possiedono genomi a base di acido nucleico. Tale mistero è stato parzialmente risolto con la scoperta che le proteine prioniche hanno proprietà insolite. Al pari delle altre proteine possono piegarsi in un certo numero di strutture tridimensionali differenti: in una configurazione, sono costituenti normali delle cellule del sistema nervoso centrale e non causano danno; in un’altra configurazione, invece, si trasformano in prioni e assumono la capacità insolita di fungere da stampo per convertire le molecole di proteine normali in prioni. I prioni e i loro precursori normali hanno la stessa sequenza aminoacidica e sono codificati dagli stessi geni: differiscono solamente nella conformazione molecolare, ovvero nel modo in cui tali componenti proteici sono ripiegati. Le proteine precursore normali dei prioni (proteina prionica cellulare o PrPc ) sono infatti ricche di siti molecolari nella conformazione α-elica, mentre tutte le proteine patologiche prioniche (PrPSc) mostrano una pre-valenza di siti molecolari che si presentano “srotolati” (unfolded) nella conformazione a foglietti-β. Contrariamente alle loro controparti normali, i prioni sono altamente resistenti alle proteasi, agli agenti chimici aggressivi e alle temperature elevate. La disinfezione di materiali contaminati da prioni richiede misure di disinfezione specifiche: per esempio, gli oggetti contaminati devono essere impregnati di idrossido di sodio 1N (normale) per almeno un’ora per poter distruggere le proteine prioniche. Come fanno i prioni a imporre il loro tipo di ripiegamento alle molecole normali? Si pensa che i prioni inducano le proteine normali a ripiegarsi nella forma di prione. Non è noto, tuttavia, se i prioni agiscano su proteine che sono ancora nel processo di piegamento o su quelle che già si sono piegate nella loro configurazione naturale. Tale processo non è stato ancora riprodotto in vitro; si ritiene che esso sia irreversibile: ossia i prioni non ritornano a essere proteine normali. Qualunque sia il meccanismo, l’accumulo di quantità sufficienti di prioni e la loro diffusione alle cellule adiacenti comporta l’alterazione delle normali funzioni cerebrali. Nelle pecore la malattia è denominata scrapie perché gli animali ammalati si strofinano intensamente su qualsiasi superficie, scorticando la propria lana e la pelle (scrape, scorticare). Con la morte di ampi aggregati cellulari nervosi, il cervello assume sempre più la forma di un formaggio svizzero con i buchi (Figura 17.11). Le proteine prioniche si aggregano in fibrille traslucide cerose dette amiloidi, che inducono la morte programmata delle cellule. Il termine “encefalopatia spongiforme” si riferisce all’aspetto spugnoso del tessuto cerebrale che consegue alla estesa necrosi cellulare. Le malattie da prioni mostrano sempre uno sviluppo lento nel tempo; è opportuno sottolineare che la funzione della forma normale dei prioni è ancora sconosciuta. I topi mutanti privi di questa proteina non mostrano sintomi. È possibile ipotizzare che se i prioni interessassero una proteina essenziale, le manifestazioni della malattia sarebbero più immediate.
 negli esseri umani e lo scrapie nelle pecore. Queste malattie sono denominate nel loro insieme encefalopatie spongiformi perché formano fori nel cervello, che fanno assumere al tessuto cerebrale un aspetto a spugna. il prione, è altamente insolito in quanto non contiene acidi nucleici, ma è composto interamente da una proteina. La natura infettiva di una di queste malattie umane, il kuru, fu ipotizzata quando venne osservata l’associazione fra persone colpite e il loro consumo di carne di cervello umano a seguito di rituali tribali. È stato poi provato che alcune di queste malattie possono essere trasmesse anche in animali da laboratorio. In tutti questi esempi patologici, i ricercatori hanno sospettato che l’agente eziologico potesse essere un virus e tuttavia non riuscivano a isolare alcun agente infettivo conosciuto (virus, batteri o altri). In seguito, fu chiarito che l’agente infettivo, poi chiamato prione , è altamente insolito in quanto non contiene acidi nucleici, ma è composto interamente da una proteina. Ciò risulta sorprendente perché tutti gli altri agenti infettivi, ovvero tutte le entità in grado di replicarsi, possiedono genomi a base di acido nucleico. Tale mistero è stato parzialmente risolto con la scoperta che le proteine prioniche hanno proprietà insolite. Al pari delle altre proteine possono piegarsi in un certo numero di strutture tridimensionali differenti: in una configurazione, sono costituenti normali delle cellule del sistema nervoso centrale e non causano danno; in un’altra configurazione, invece, si trasformano in prioni e assumono la capacità insolita di fungere da stampo per convertire le molecole di proteine normali in prioni. I prioni e i loro precursori normali hanno la stessa sequenza aminoacidica e sono codificati dagli stessi geni: differiscono solamente nella conformazione molecolare, ovvero nel modo in cui tali componenti proteici sono ripiegati. Le proteine precursore normali dei prioni (proteina prionica cellulare o PrPc ) sono infatti ricche di siti molecolari nella conformazione α-elica, mentre tutte le proteine patologiche prioniche (PrPSc) mostrano una pre-valenza di siti molecolari che si presentano srotolati (unfolded) nella conformazione a foglietti-β. Contrariamente alle loro controparti normali, i prioni sono altamente resistenti alle proteasi, agli agenti chimici aggressivi e alle temperature elevate. La disinfezione di materiali contaminati da prioni richiede misure di disinfezione specifiche: per esempio, gli oggetti contaminati devono essere impregnati di idrossido di sodio 1N (normale) per almeno un’ora per poter distruggere le proteine prioniche. Come fanno i prioni a imporre il loro tipo di ripiegamento alle molecole normali Si pensa che i prioni inducano le proteine normali a ripiegarsi nella forma di prione. Non è noto, tuttavia, se i prioni agiscano su proteine che sono ancora nel processo di piegamento o su quelle che già si sono piegate nella loro configurazione naturale. Tale processo non è stato ancora riprodotto in vitro; si ritiene che esso sia irreversibile: ossia i prioni non ritornano a essere proteine normali. Qualunque sia il meccanismo, l’accumulo di quantità sufficienti di prioni e la loro diffusione alle cellule adiacenti comporta l’alterazione delle normali funzioni cerebrali. Nelle pecore la malattia è denominata scrapie perché gli animali ammalati si strofinano intensamente su qualsiasi superficie, scorticando la propria lana e la pelle (scrape, scorticare). Con la morte di ampi aggregati cellulari nervosi, il cervello assume sempre più la forma di un formaggio svizzero con i buchi (Figura 17.11). Le proteine prioniche si aggregano in fibrille traslucide cerose dette amiloidi, che inducono la morte programmata delle cellule. Il termine encefalopatia spongiforme si riferisce all’aspetto spugnoso del tessuto cerebrale che consegue alla estesa necrosi cellulare. Le malattie da prioni mostrano sempre uno sviluppo lento nel tempo; è opportuno sottolineare che la funzione della forma normale dei prioni è ancora sconosciuta. I topi mutanti privi di questa proteina non mostrano sintomi. È possibile ipotizzare che se i prioni interessassero una proteina essenziale, le manifestazioni della malattia sarebbero più immediate.")
61
Le proteine prioniche si aggregano in fibrille traslucide cerose dette amiloidi, che inducono la morte programmata delle cellule. Il termine “encefalopatia spongiforme” si riferisce all’aspetto spugnoso del tessuto cerebrale che consegue alla estesa necrosi cellulare. Le malattie da prioni mostrano sempre uno sviluppo lento nel tempo; è opportuno sottolineare che la funzione della forma normale dei prioni è ancora sconosciuta

62
TAKE HOME MESSAGE ENTITÀ BIOLOGICHE SUBCELLULARI
PARASSITI ENDOCELLULARI OBBLIGATI POSSIEDONO UN SOLO TIPO DI ACIDO NUCLEICO (DNA O RNA) HANNO DIMENSIONI PICCOLISSIME (SONO FILTRABILI) POSSIEDONO UN INVOLUCRO PROTEICO (CAPSIDE) E, A VOLTE, UN INVOLUCRO LIPOPROTEICO(PERICAPSIDEO PEPLOS) PRESENTANO IN SUPERFICIE STRUTTURE PROTEICHE (ANTIRECETTORI) CHE SI ANCORANO A SPECIFICI RECETTORI DELLA CELLULA BERSAGLIO
 HANNO DIMENSIONI PICCOLISSIME (SONO FILTRABILI) POSSIEDONO UN INVOLUCRO PROTEICO (CAPSIDE) E, A VOLTE, UN INVOLUCRO LIPOPROTEICO(PERICAPSIDEO PEPLOS) PRESENTANO IN SUPERFICIE STRUTTURE PROTEICHE (ANTIRECETTORI) CHE SI ANCORANO A SPECIFICI RECETTORI DELLA CELLULA BERSAGLIO.")
63
TAKE HOME MESSAGE NON SONO ESSERI VIVENTI
PER ESISTERE IN NATURA DEVONO ESSERE INFETTANTI DEVONO UTILIZZARE I MECCANISMI DELLA CELLULA OSPITE PER PRODURRE I PROPRI COMPONENTI (MRNA VIRALI, PROTEINE E COPIE IDENTICHE DEL GENOMA) DEVONO CODIFICARE PROPRIE PROTEINE SPECIFICHE PER OGNI PROCESSO DA LORO RICHIESTO E NON EFFETTUABILE DALLA CELLULA LA PRODUZIONE DI NUOVI VIRIONI AVVIENE PER ASSEMBLAGGIO DELLE COMPONENTI VIRALI
 DEVONO CODIFICARE PROPRIE PROTEINE SPECIFICHE PER OGNI PROCESSO DA LORO RICHIESTO E NON EFFETTUABILE DALLA CELLULA. LA PRODUZIONE DI NUOVI VIRIONI AVVIENE PER ASSEMBLAGGIO DELLE COMPONENTI VIRALI.")
64
GRAZIE PER L’ATTENZIONE















 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")





