Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
L’impacchettamento del templato in nucleosomi influisce su
tutti gli stadi della trascrizione: legame con gli attivatori formazione del PIC allungamento I fattori di trascrizione sfruttano diverse strategie per ottenere un legame efficiente

2
Struttura della Cromatina ed Espressione Genica
Permanentemente represso Represso ma inducibile Attivo. Può essere modulato Legame di fattori trascrizionali

3
I meccanismi della trascrizione su DNA nudo sono simili
in tutti gli organismi nonostante l’aumento di complessità nel macchinario di trascrizione stesso 1°: Legame di attivatori a monte del promotore 2°: Reclutamento di complessi adattatori tipo SAGA, o Mediator 3°: Facilitazione del legame dei fattori di trascrizione generale 4°: Posizionamento di Pol II da parte di TFIID, TFIIA TFIIB Formazione del complesso di preinizio 5°: Fusione di 11-15bp da parte di TFIIH 6°: Fosforilazione del CTD di Pol II durante la sintesi delle prime 30bp 7°: Rilascio dei GTFs e inizio dell’allungamento

4
Studi genomici hanno dimostrato che la densità nucleosomale
è minore sui promotori che sulle regioni codificanti Microarrays ad alta risoluzione hanno mostrato che esistono regioni NUCLEOSOME-FREE di circa 200bp sulla maggioranza dei promotori Questi spesso corrispondono a siti DHS Di solito sono fiancheggiati da nucleosomi posizionati I siti DHS sono generati e mantenuti da diversi elementi: Attivatori della trascrizione (ad es. Pho4) Acetilazione (NuA4) Complessi di rimodellamento La combinazione dei tre elementi Legame di fattori di boundary (Rap1) e/o di elementi di sequenza che escludono i nucleosomi (dAdT)
 Acetilazione (NuA4) Complessi di rimodellamento. La combinazione dei tre elementi. Legame di fattori di boundary (Rap1) e/o di elementi di sequenza che. escludono i nucleosomi (dAdT)")
5
Un esempio di gene costitutivamente attivo in lievito: CYC 1
Dominio di Attivazione UAS 2 UAS 1 TATA Dominio stabile Le 60 bp tra i due fattori escludono i nucleosomi poiché i fattori hanno alta affinità per i siti. Il legame alle UAS è più dinamico, ma la perdita di un fattore non implica la riformazione del nucleosoma.

6
Non sempre la presenza del nucleosoma esclude la possibilità di trascrivere un gene
E’ possibile in certi casi che i siti per l’interazione con i fattori di trascrizione si trovino adiacenti sulla superficie del nucleosoma e che questo, di fatto, favorisca l’assemblaggio del macchinario di trascrizione

7
MMTV è come un gene inducibile nei mammiferi
(MMTV: mouse mammary tumor virus) -32 -210 -430 -600 -810 F E D C B A -200 NF1 HRE Otf 1 Questi fattori sono capaci di legare il DNA nudo, ma non tutti insieme. NF1 e Otf1 non possono legare da soli i nucleosomi fasati. Due siti HRE vengono occupati sulla superficie del nucleosoma. Ciò espone i successivi siti HRE ed avviene il legame. Questo permette il legame di ENTRAMBI NF1 e Otf1.
 F. E. D. C. B. A NF1. HRE. Otf 1. Questi fattori sono capaci di legare il DNA nudo, ma non tutti insieme. NF1 e Otf1 non possono legare da soli i nucleosomi fasati. Due siti HRE vengono occupati sulla superficie del nucleosoma. Ciò espone i successivi siti HRE ed avviene il legame. Questo permette il legame di ENTRAMBI NF1 e Otf1.")
8
Il Legame dipende dalla natura del promotore dell’ MMTV
I Fattori non possono legarsi tutti contemporaneamente sul DNA substrato I Fattori possono tutti legare il DNA substrato quando questo è in una conformazione nucleosomale.

9
LA CROMATINA VIENE SOTTOPOSTA A RIMODELLAMENTO
GENI INDUCIBILI LA CROMATINA VIENE SOTTOPOSTA A RIMODELLAMENTO IN MODO CHE LE REGIONI REGOLATRICI DEI GENI POSSANO DIVENTARE ACCESSIBILI AL MACCHINARIO DELLA TRASCRIZIONE

10
Attivazione del gene PHO 5 in bassi livelli di fosfato
- 5 - 4 - 3 - 2 - 1 T A P H O 5 Sito Pho2 Siti Pho4 Pho4 viene defosforilato per potersi legare al suo sito A T P H O 5 La cromatina viene rimodellata in modo da lasciare scoperti i siti per l’attivazione della trascrizione

11
Dinamicità della cromatina
Nonostante la stabilità del nucleosoma e dell’alto grado di compattezza nel nucleo la cromatina è sorprendentemente dinamica. Esistono vari modi di modificare la cromatina e rispondere alle necessità della cellula di regolare i processi biologici dall’espressione genica, alla riparazione del DNA, alla replicazione e ricombinazione. Uso di varianti istoniche, in particolare varianti di H2A e H3. Modificazioni epigenetiche degli istoni: acetilazione, metilazione ed altre. Uso di sistemi di rimodellamento che cambiano la stabilità e/o la posizione del nucleosoma.

12
Il legame dei fattori e la perdita degli istoni possono anche essere consequenziali. Questo di solito coinvolge la partecipazione di FATTORI DI RIMODELLAMENTO

13
Le variazioni di struttura della cromatina si possono studiare usando la MNasi
Questo enzima non taglia sul nucleosoma ma solo tra particelle adiacenti. Si possono ottenere digesti parziali Dopo la digestione gli istoni vengono rimossi R R Sonda Taglio con R, elettroforesi e blot.

14
ATTIVAZIONE DEL GENE ADH2 IN ETANOLO
MN U MN U RIS TATA +1 +2 +3 +4 -1 Rep -2 TATA -1 +1 +2 +3 +4 +1 +2 +3 +4 -1 RIS Ind Modificazioni Nucleosomali nessuna media alta bassa Represso Indotto

15
RIMOZIONE DEGLI ISTONI
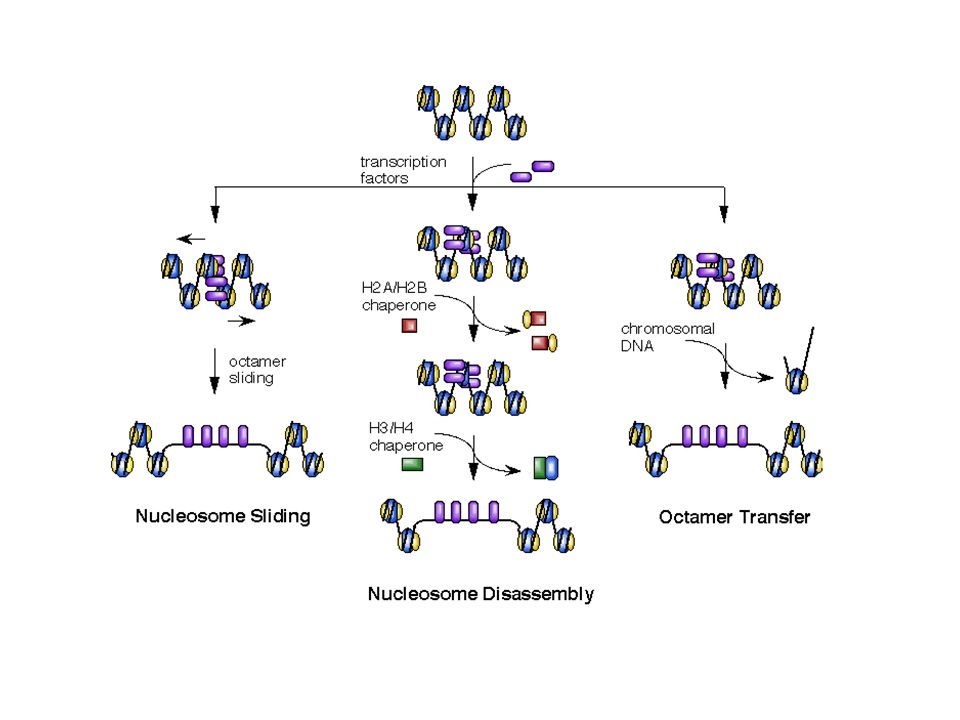
La rimozione degli istoni da una regione di DNA può avvenire tramite: Slittamento dei nucleosomi lungo una sequenza Disassemblaggio del nucleosoma tramite rimozione sequenziale degli istoni Trasferimento degli istoni su un’altra regione del DNA Rimodellamento è anche CAMBIAMENTO CONFORMAZIONALE SUL NUCLEOSOMA CON ESPOSIZIONE DI PORZIONI DI DNA

17
L’accessibilità dei fattori di trascrizione al promotore e/o la
liberazione di questo dai nucleosomi si ottiene con la partecipazione di: SISTEMI DI RIMODELLAMENTO DELLA CROMATINA Di questi sistemi ne sono stati trovati finora due classi: Complessi di rimodellamento della cromatina ATP-dipendenti Complessi di rimodellamento che contengono Acetiltrasferasi o Deacetilasi Istoniche

18
ACETILTRASFERASI NUCLEARI (HAT)
Le HAT nucleari sono contenute in complessi multisubunità di massa molecolare molto alta. In lievito i principali sono: SAGA = 1.8MDa NuA4 = 1.4MDa NuA3 = 0.5MDa ADA = 0.8MDa

19
Deacetilasi e Acetiltrasferasi in lievito
Funzionano tutte attraverso il legame di altri fattori che le portano sul templato Rpd3 (correlata ad HDAC1,2,3) lega Sin3 è portata sul promotore da Ume6 Hda1 (correlata ad HDAC 4, 5, 6) lega Tup1 Gcn5 (subunità di SAGA con molti omologhi) lega Gcn4 Esa1 (subunità di NuA4 con molti omologhi) lega Gcn4 Altre deacetilasi di lievito: Hos1 e Hos3 lavorano principalmente sui geni ribosomali Hos2 deacetila le regioni codificanti
 lega Sin3. è portata sul promotore da Ume6. Hda1 (correlata ad HDAC 4, 5, 6) lega Tup1. Gcn5 (subunità di SAGA con molti omologhi) lega Gcn4. Esa1 (subunità di NuA4 con molti omologhi) lega Gcn4. Altre deacetilasi di lievito: Hos1 e Hos3 lavorano principalmente sui geni ribosomali. Hos2 deacetila le regioni codificanti.")
20
Specificità di azione degli enzimi HAT e HDAC in lievito

21
Geni regolati da Ume6 Es: INO1 Geni regolati da Gcn4 Es: HIS3

22
Membri di tre differenti
famiglie HAT Il dominio Acetiltrasferasico è conservato nelle famiglie e comune a tutte.

23
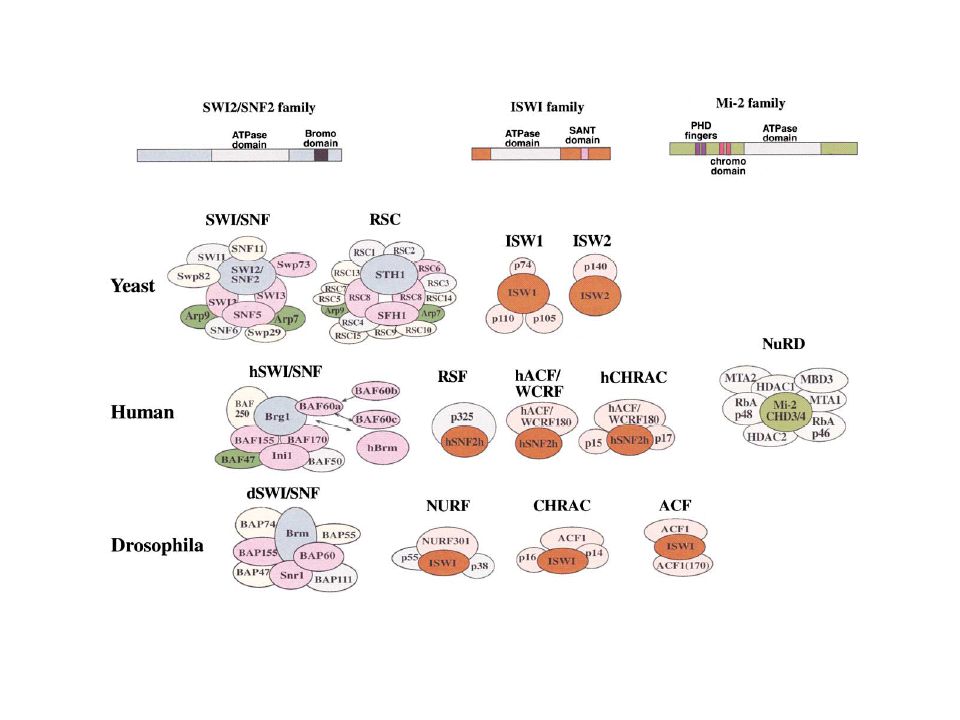
Complessi di Rimodellamento ATP-dipendenti
CLASSE SWI/SNF: SWI/SNF = 2Md (ATPasi Snf2) RSC = 1Md (ATPasi Sfh2) CLASSE ISWI: Tutti complessi di circa Kd ATPasi comune ISWI NURF CHRAC ACF CLASSE Mi2: ATPasi Mi2-CHD3/4
 RSC = 1Md (ATPasi Sfh2) CLASSE ISWI: Tutti complessi di circa Kd. ATPasi comune ISWI. NURF. CHRAC. ACF. CLASSE Mi2: ATPasi Mi2-CHD3/4.")
25
Il rimodellamento della cromatina
può essere un meccanismo che coinvolge anche diversi passaggi. Complessi di rimodellamento di tipo diverso possono cooperare per aprire la cromatina.

26
Cascata di eventi nell’inizio
della trascrizione del gene per IFN-β In seguito a infezione virale: il promotore diventa progressivamente più acetilato da parte di GCN5 umano su Lys8H4 e Lys9,14H3 AcLys8H4 recluta SWI/SNF attraverso il BROMODOMINIO della subunità BRG1 segue il reclutamento di TFIID attraverso il BROMODOMINIO di TAFII250 che lega AcLys9,14H3

27
I complessi di rimodellamento ATP-dipendenti hanno molteplici
attività biochimiche studiate in vitro a) Alterazione dei profili di taglio della DNasiI b) Formazione di nucleosomi rimodellati d) Trasferimento dell’ottamero istonico IN TUTTI I CASI IL PROFILO DNasiI è ALTERATO e) Alterazione delle posizioni traslazionali degli ottameri f) Assemblaggio e spaziatura dei nucleosomi g) estrusione di struture cruciformi
 Alterazione dei. profili di taglio. della DNasiI. b) Formazione di. nucleosomi. rimodellati. d) Trasferimento. dell’ottamero. istonico. IN TUTTI I CASI. IL PROFILO. DNasiI è ALTERATO. e) Alterazione delle. posizioni traslazionali. degli ottameri. f) Assemblaggio e. spaziatura dei. nucleosomi. g) estrusione di. struture. cruciformi.")
28
h) Rimodellamento topologico
in molecole circolari chiuse nucleosomizzate g) Accesso alterato agli enzimi di restrizione IL NUMERO DI SUPERAVVOLGIMENTI PUO’ ESSERE RIDOTTO SENZA LA PERDITA DI OTTAMERI ISTONICI
 Accesso alterato agli. enzimi di restrizione. IL NUMERO DI SUPERAVVOLGIMENTI. PUO’ ESSERE RIDOTTO SENZA LA. PERDITA DI OTTAMERI ISTONICI.")
29
Modelli per la regolazione della cromatina durante l’inizio della trascrizione

30
Le proteine regolatrici e i
complessi di rimodellamento possono agire in combinazione e in ordine differente, e alla fine comportano sempre lo stesso risultato: un substrato pronto per la trascrizione.

Presentazioni simili