Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Le strutture batteriche

2
Il peptidoglicano (PGN)
")
3
Il Peptidoglicano (PGN): [1]
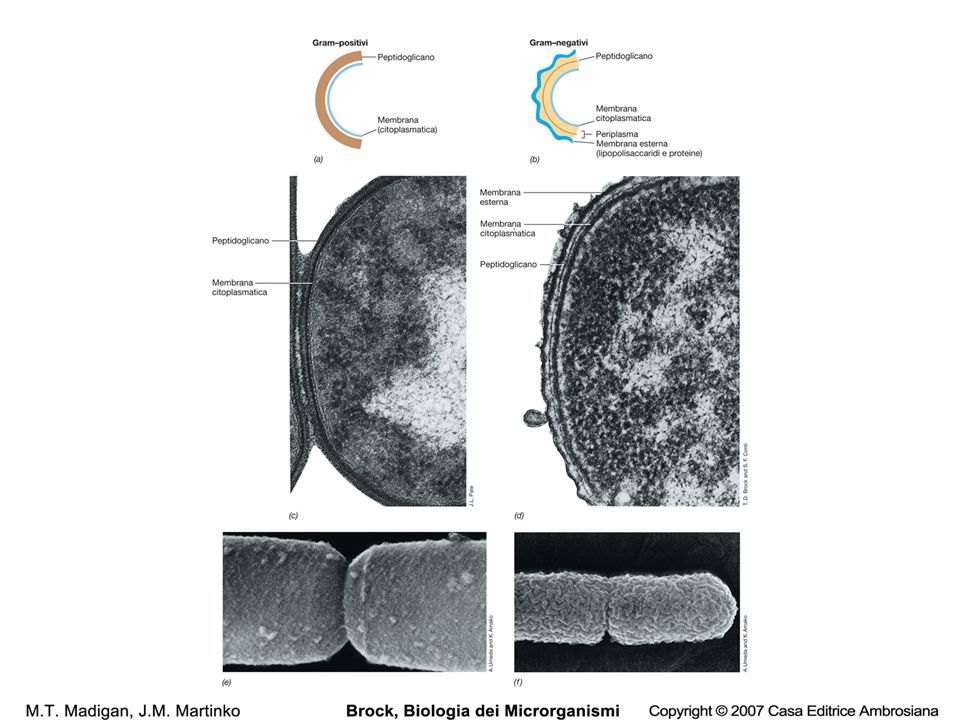
Il componente principale della parete batterica Responsabile della conformazione e della rigidità della parete Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [1]](http://slideplayer.it/slide/2348156/8/images/3/Il+Peptidoglicano+%28PGN%29%3A+%5B1%5D.jpg "Il componente principale della parete batterica. Responsabile della conformazione e della rigidità della parete. Esclusivo dei batteri…bersaglio degli antibiotici.")
6
Il Peptidoglicano (PGN): [2]
Preserva l’integrità della cellula batterica L’’inibizione della biosisntesi del PGN, sia attraverso mutazioni, sia attraverso il trattamento con antibiotici, o la rottura della struttura del PGN (lisozima) si accompagna alla lisi della cellula batterica. Contribuisce al mantenimento di una forma definita Serve come scaffold per le altre componenti della parete È assente in alcuni batteri: Mycoplasma, Planctomyces e in alcuni ceppi di Rickettsia In Chlamidia, seppur presenti i geni per la biosintesi del PGN, non è stato mai identificato Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [2]](http://slideplayer.it/slide/2348156/8/images/6/Il+Peptidoglicano+%28PGN%29%3A+%5B2%5D.jpg "Preserva l’integrità della cellula batterica. L’’inibizione della biosisntesi del PGN, sia attraverso mutazioni, sia attraverso il trattamento con. antibiotici, o la rottura della struttura del PGN (lisozima) si accompagna alla lisi della cellula. batterica. Contribuisce al mantenimento di una forma definita. Serve come scaffold per le altre componenti della parete. È assente in alcuni batteri: Mycoplasma, Planctomyces e in alcuni ceppi di. Rickettsia. In Chlamidia, seppur presenti i geni per la biosintesi del PGN, non è stato mai. identificato. Esclusivo dei batteri…bersaglio degli antibiotici.")
7
Resistenza del PGN Unica grossa molecola nelle tre direzioni dello spazio Legami b-1,4 particolarmente forti Presenza di zuccheri non comuni (MurNac) Alternanza di aminoacidi D e L risulta in una resistenza maggiore Sembra che tra i vari gruppi peptidici si stabiliscano numerosi legami idrogeno
 Alternanza di aminoacidi D e L risulta in una resistenza maggiore. Sembra che tra i vari gruppi peptidici si stabiliscano numerosi legami idrogeno.")
8
Catene lineare di n unità di un disaccaride:
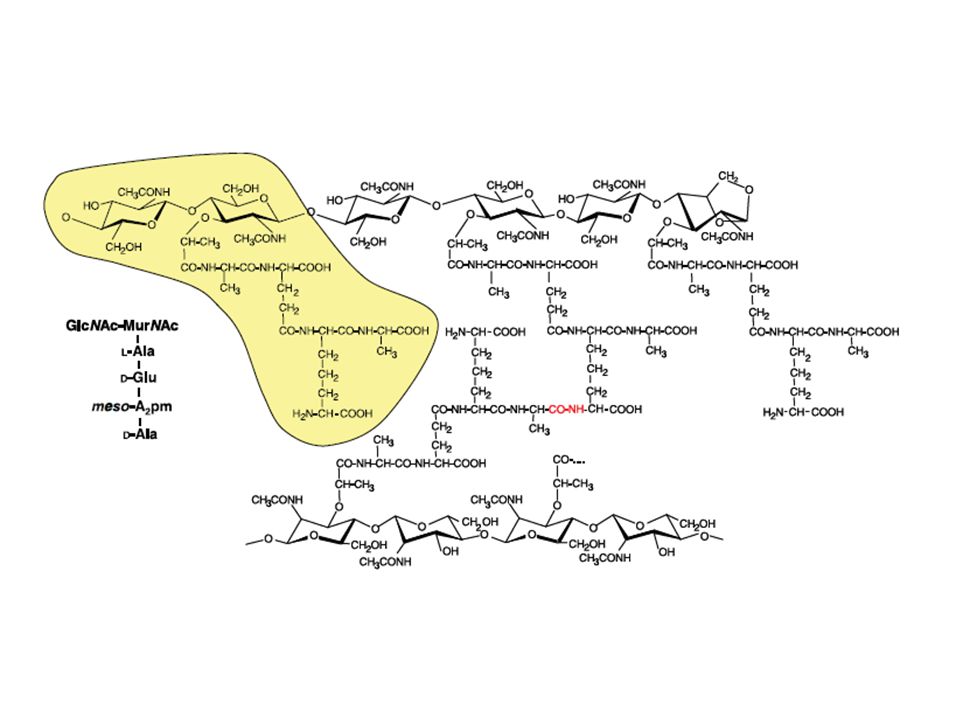
Nam nag formano catene di lunghezza variabile da 10 a 80 unità saccarisiche ripetute che vanno a formare un reticolato macromolecolare rigido e resistente alla trazione Il gruppo carbossilico del gruppo lattilico del NAM si lega al gruppo aminico del primo amminoacido del tetrapeptide mediante un legame carboaminico La sequenza di aminoacidi nei tetrapeptidi può variare da una specie batterica all’altra, ma contiene sempre in terza posizione un aminoacido BIBASICO lisina G+ dap G_ COMPONENTE GLUCIDICA Catene lineare di n unità di un disaccaride: N-acetil-D-glucosammina (NAG) Acido N-acetil-muramico (NAM) COMPONENTE PEPTIDICA Corta catena di aminoacidi (TETRAPEPTIDE) alternati negli stereoisomeri D e L
 Acido N-acetil-muramico (NAM) COMPONENTE PEPTIDICA. Corta catena di aminoacidi (TETRAPEPTIDE) alternati negli stereoisomeri D e L.")
9
Il Peptidoglicano (PGN)
")
11
Il Peptidoglicano (PGN): il legame delle catene peptidiche [1]
![Il Peptidoglicano (PGN): il legame delle catene peptidiche [1]](http://slideplayer.it/slide/2348156/8/images/11/Il+Peptidoglicano+%28PGN%29%3A+il+legame+delle+catene+peptidiche+%5B1%5D.jpg "Il Peptidoglicano (PGN): il legame delle catene peptidiche [1]")
12
Il Peptidoglicano (PGN): il legame delle catene peptidiche [2]
![Il Peptidoglicano (PGN): il legame delle catene peptidiche [2]](http://slideplayer.it/slide/2348156/8/images/12/Il+Peptidoglicano+%28PGN%29%3A+il+legame+delle+catene+peptidiche+%5B2%5D.jpg "Il Peptidoglicano (PGN): il legame delle catene peptidiche [2]")
13
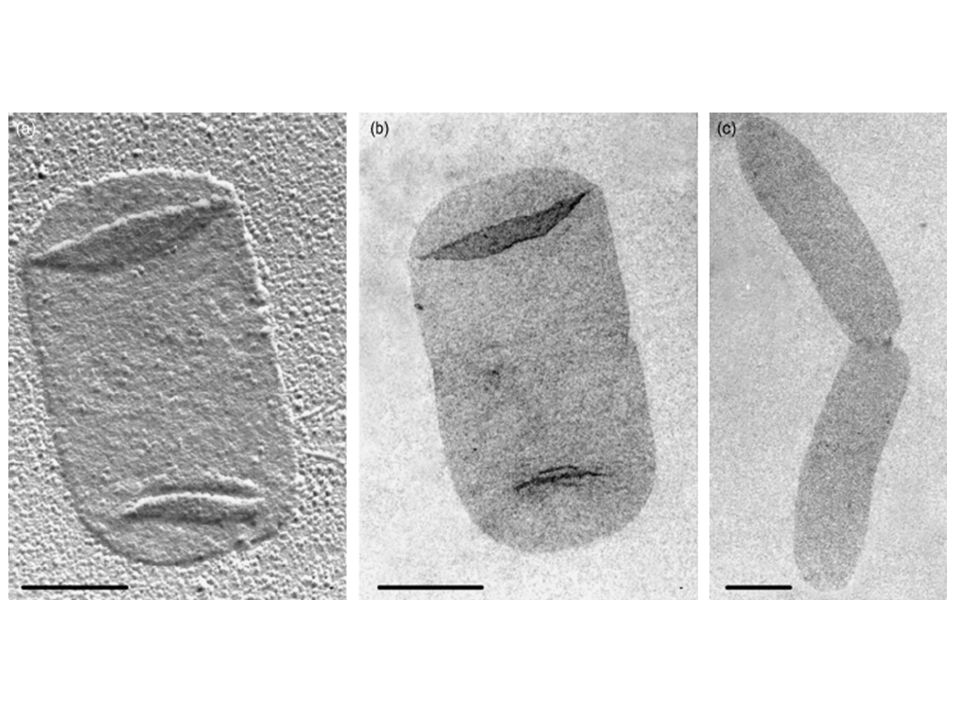
I tetrapeptidi I tetrapeptidi si legano tra di loro con due modalità principali: DIRETTAMENTE (nei Gram negativi) INDIRETTAMENTE (nei Gram positivi) Direttamente: tramite un legame peptidico tra il gruppo aminico dell’aminoacido bibasico di un tetrapeptide e il gruppo carbossilico dell’aminoacido terminale (D-ALA) INDIRETTAMENTE tramite un ponte di pentaglicina: il gruppo amminoco dell’aminoacdio bibasico di un tetrapeptide forma un legame peptidico con il gruppo carbossilico della pentaglicina Il gruppo aminico della pentaglicina fomra un legame peptidico con il gruppo carbossilico dell’ultima D-ALA
 Direttamente: tramite un legame peptidico tra il gruppo aminico dell’aminoacido bibasico di un tetrapeptide e il gruppo carbossilico dell’aminoacido terminale (D-ALA) INDIRETTAMENTE tramite un ponte di pentaglicina: il gruppo amminoco dell’aminoacdio bibasico di un tetrapeptide forma un legame peptidico con il gruppo carbossilico della pentaglicina. Il gruppo aminico della pentaglicina fomra un legame peptidico con il gruppo carbossilico dell’ultima D-ALA.")
15
Il Peptidoglicano (PGN): [3]
Le catene glicaniche vengono assemblate attraverso l’oligomerizzazione di unità disaccaridiche/peptidiche. Queste reazioni sono catalizzate da numerosi enzimi e vengono indicate come reazioni di transglicosilazione. È posibile stabilire una lunghezza delle catene glicaniche? La lunghezza media delle catene varia considerevolmente da batterio a batterio e all’interno della stessa popolazione rispetto alla fase di crescita, alla composizione del mezzo di crescita, presenza/assenza di antibiotici, condizioni di crescita extra/intra cellulari Nel PGN dei Bacilli, B. subtilis, B. cereus, B. licheniformis, lo sheletro saccaridico ha una lunghezza compresa tra le 50 e le 250 unità saccaridiche Negli Staphilococchi, S. areus, le catene sembrano essere molto più corte con una lunghezza media di 18 unità Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [3]](http://slideplayer.it/slide/2348156/8/images/15/Il+Peptidoglicano+%28PGN%29%3A+%5B3%5D.jpg "Le catene glicaniche vengono assemblate attraverso l’oligomerizzazione di unità disaccaridiche/peptidiche. Queste reazioni sono catalizzate da numerosi enzimi e vengono indicate come reazioni di transglicosilazione. È posibile stabilire una lunghezza delle catene glicaniche La lunghezza media delle catene varia considerevolmente da batterio a batterio e all’interno della stessa popolazione rispetto alla fase di crescita, alla composizione del mezzo di crescita, presenza/assenza di antibiotici, condizioni di crescita extra/intra cellulari. Nel PGN dei Bacilli, B. subtilis, B. cereus, B. licheniformis, lo sheletro saccaridico ha una lunghezza compresa tra le 50 e le 250 unità saccaridiche. Negli Staphilococchi, S. areus, le catene sembrano essere molto più corte con una lunghezza media di 18 unità. Esclusivo dei batteri…bersaglio degli antibiotici.")
16
Biosintesi del PGN

17
Biosintesi del PGN La sintesi del PGN ha luogo in tre diversi compartimenti 1. Fase citoplasmatica 2. Fase di membrana 3. Fase periplasmatica Fase citoplasmatica Sintesi dei precursori di natura saccaridica e proteica Fase di membrana Assemblaggio del glican-pentapeptide e attraversamento della membrana citoplasmatica Fase periplasmatica Polimerizzazione del PGN (reazioni di transglicosilazione) e formazione dei legami crociati (reazioni di transpeptidazione)
 e formazione dei legami crociati (reazioni di transpeptidazione)")
18
Fase citoplasmatica (stage I)
Sintesi dei precursori di natura saccaridica e proteica Sintesi del NAG Si parte dal fruttosio-6 fosfato e attraverso l’azione coordinata di diversi enzimi si arriva alla formazione del primo substrato attivato del PGN: l’UDP-NAG

19
Fase citoplasmatica (stage I)
Sintesi dei precursori di natura saccaridica e proteica Sintesi del NAM Nucleotide di Park UDP-NAM

20
Fase di membrana (stage II)
Formazione del glican-pentapeptide e traslocazione nel periplasma Bactoprenolo

21
Fase di membrana (stage III)
Polimerizzazione del PGN e formazione dei legami crociati

22
Fase di membrana (stage III)
Polimerizzazione del PGN e formazione dei legami crociati Sintasi mureiniche Enzimi presenti fino a circa 200 copie per cellula batterica Possono essere monofunzionali e quindi agire o come transpeptidasi (TP) o come transglicosilasi (TG) Bifunzionali: hanno attività sia di TP che TG Tutte le sintasi mureiniche sono ancorate alla membrana citoplasmatica grazie a un dominio transmembrana adiacente la regione N-terminale (citoplasma) Il dominio catalitico è presene all’estremità C-terminale Un gruppo specifico di sintasi con attività TP sono le PBPs (penicillin-binding proteins)
 o come transglicosilasi (TG) Bifunzionali: hanno attività sia di TP che TG. Tutte le sintasi mureiniche sono ancorate alla membrana citoplasmatica grazie a un dominio transmembrana adiacente la regione N-terminale (citoplasma) Il dominio catalitico è presene all’estremità C-terminale. Un gruppo specifico di sintasi con attività TP sono le PBPs (penicillin-binding proteins)")
23
PBPs in E. coli

24
Il Peptidoglicano (PGN): [4]
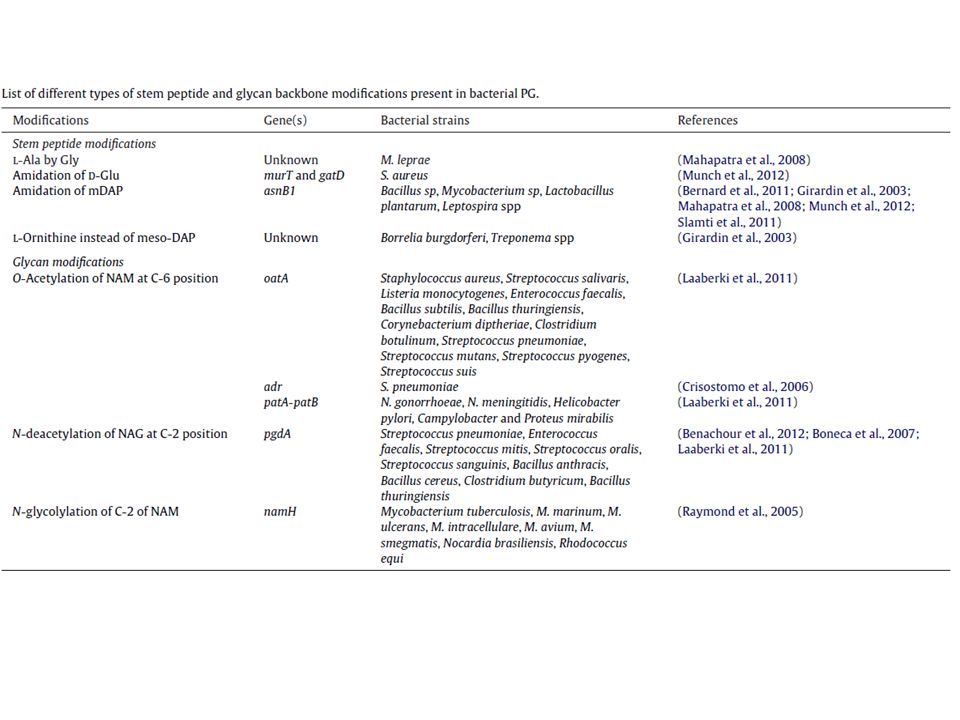
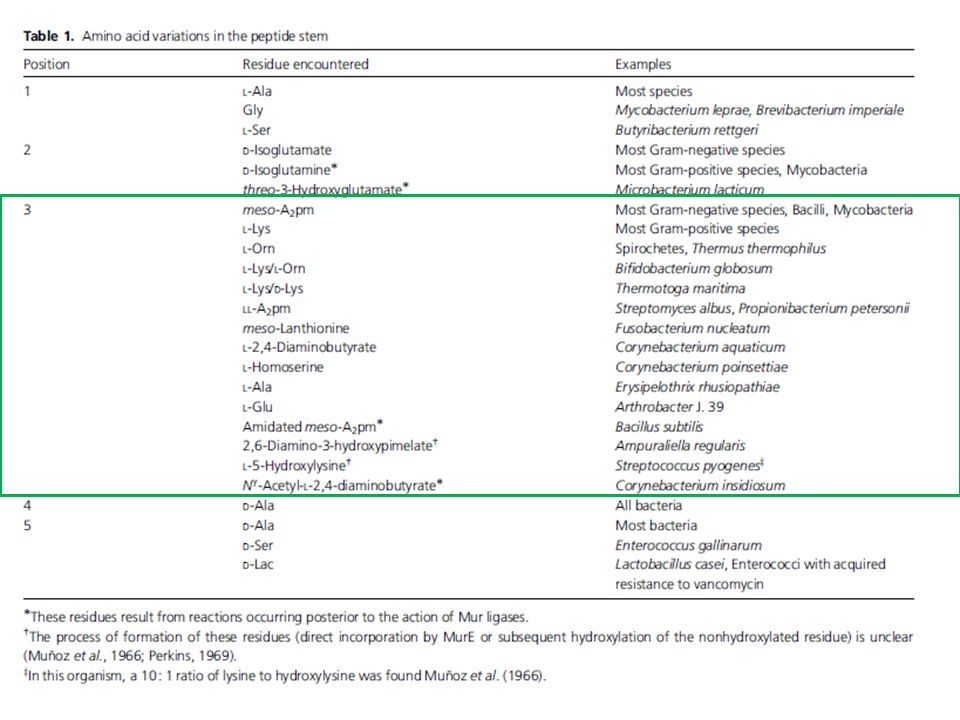
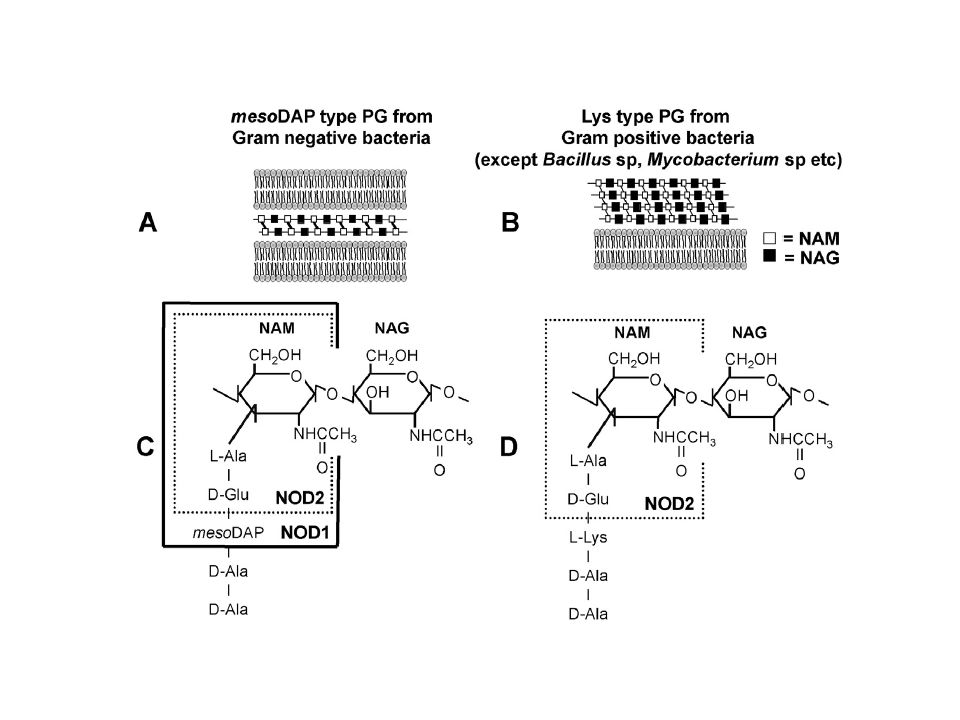
Come per l’LPS anche per il PGN esistono numerose modificazioni, che si realizzano sia a carico dello scheletro glucidico, sia a livello della componente proteica. 1. le modificazioni della componente saccaridica possono essere fatte rientrare nelle seguenti categorie: deacetilazione, acetilazione, glicolilazione 2. Le variazioni della componente peptidica (peptide stem) sono ascrivibili a due meccanismi: a) azione degli enzimi Mur (durante la biosintesi) b) fasi tardive della biosintesi: amidazione, acetilazione, idrossilazione, aggancio di gruppi proteici Le maggiori variazioni si osservano a livello dell’aminoacido in terza posizione! Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [4]](http://slideplayer.it/slide/2348156/8/images/24/Il+Peptidoglicano+%28PGN%29%3A+%5B4%5D.jpg "Come per l’LPS anche per il PGN esistono numerose modificazioni, che si realizzano sia a carico dello scheletro glucidico, sia a livello della componente proteica. 1. le modificazioni della componente saccaridica possono essere fatte rientrare nelle seguenti categorie: deacetilazione, acetilazione, glicolilazione. 2. Le variazioni della componente peptidica (peptide stem) sono ascrivibili a due meccanismi: a) azione degli enzimi Mur (durante la biosintesi) b) fasi tardive della biosintesi: amidazione, acetilazione, idrossilazione, aggancio di gruppi proteici. Le maggiori variazioni si osservano a livello dell’aminoacido in terza. posizione! Esclusivo dei batteri…bersaglio degli antibiotici.")
28
Il Peptidoglicano (PGN): Modificazioni nella componente glicanica
: Modificazioni nella componente glicanica")
29
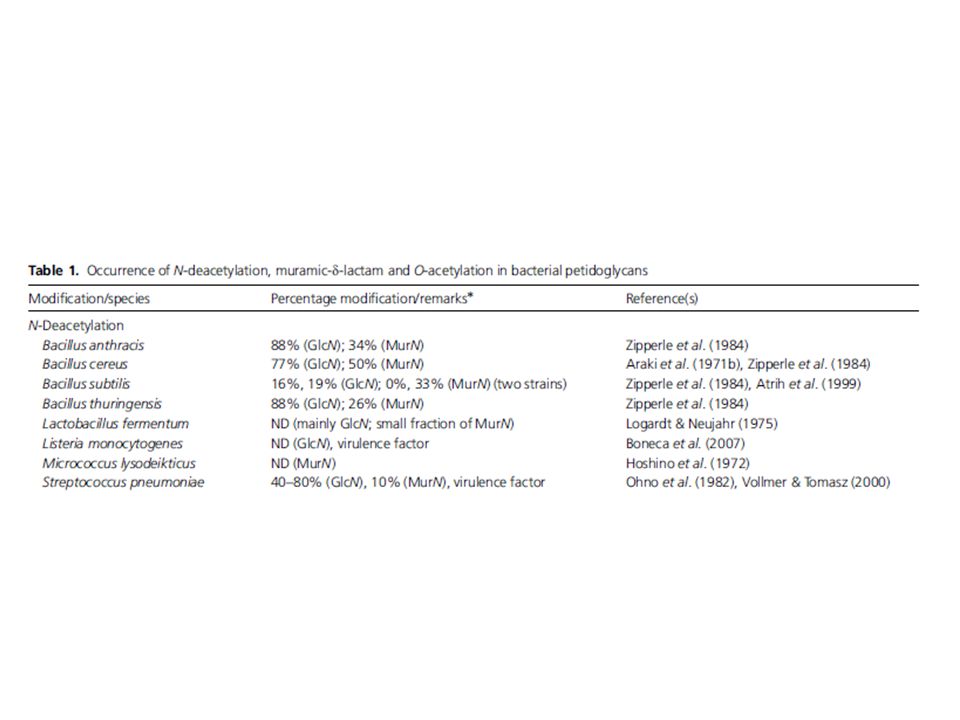
Modificazioni nella componente glicanica [1]: la deacetilazione
Osservata inizialmente in ceppi di S. pneumoniae reisistenti all’attività del lisozima: una significativa proporzione dei residui di GlcNAC risulta deacetilata B cereus, B. anthracis, L. monocytogenes, E. faecalis, H. pylori In S. pneuomniae è stato identificato il gene responsabile di questa modifica: pgdA (peptidoglycan deacetilase A) PgdA appartiene a una superfamiglia di proteine: le esterasi saccardidiche di classe 4 I membri di questa famiglia presentano due caratteristiche: 1) vengono espresse come proteine con una sequenza segnale di taglio; 2) necessitano, per l’attivazione, del legame a ioni (Zn, Co, Mg)
![Modificazioni nella componente glicanica [1]: la deacetilazione](http://slideplayer.it/slide/2348156/8/images/29/Modificazioni+nella+componente+glicanica+%5B1%5D%3A+la+deacetilazione.jpg "Osservata inizialmente in ceppi di S. pneumoniae reisistenti all’attività del lisozima: una significativa proporzione dei residui di GlcNAC risulta deacetilata. B cereus, B. anthracis, L. monocytogenes, E. faecalis, H. pylori. In S. pneuomniae è stato identificato il gene responsabile di questa modifica: pgdA (peptidoglycan deacetilase A) PgdA appartiene a una superfamiglia di proteine: le esterasi saccardidiche di classe 4. I membri di questa famiglia presentano due caratteristiche: 1) vengono espresse come proteine con una sequenza segnale di taglio; 2) necessitano, per l’attivazione, del legame a ioni (Zn, Co, Mg)")
31
Qual è il ruolo biologico della deacetilazione del PGN?
La presenza di amminozuccheri deacetilati nella composizione del PGN riduce drasticamente l’attività del lisozima Lisozima: enzima con attività muramidasica ubiquitariamente presente nei liquidi corporei e nelle secrezioni, nei fagociti (granuli), cellule del Paneth
, cellule del Paneth.")
32
Qual è il ruolo biologico della deacetilazione del PGN?
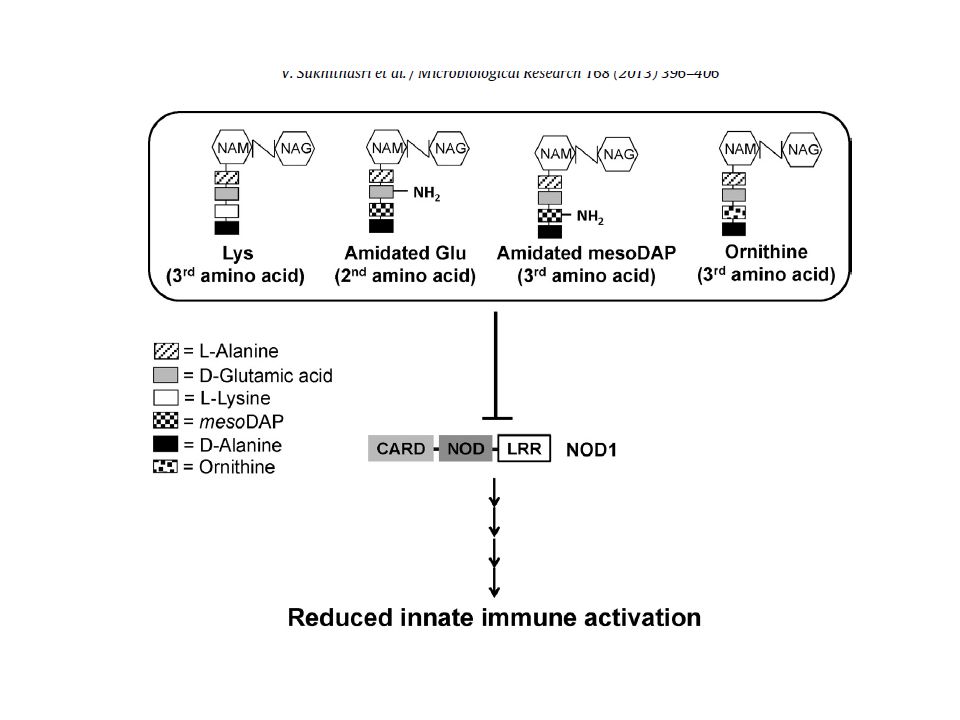
È possibile che l’effetto della deacetilazione del PGN non sia limitato alla riduzione dell’attività enzimatica del lisozima La deacetilazione introduce cariche positive nella parete cellulare con conseguente modulazione dei legami con altre strutture batteriche di superficie (proteine) o altri componenti batterici (capsula polisaccaridica) Maggiore resistenza del batterio ai peptidi cationici antimicrobici (AMPs)) dell’ospite La deacetilazione può alterare/modulare il riconoscimento di un patogeno attraverso i PRRs: L. monocytogenes, mutante pgdA risposta interferonica (IFN-b) attraverso il riconoscimento mediato da TLR2 e NOD1
 o altri componenti batterici (capsula polisaccaridica) Maggiore resistenza del batterio ai peptidi cationici antimicrobici (AMPs)) dell’ospite. La deacetilazione può alterare/modulare il riconoscimento di un patogeno attraverso i PRRs: L. monocytogenes, mutante pgdA risposta interferonica (IFN-b) attraverso il riconoscimento mediato da TLR2 e NOD1.")
33
Modificazioni nella componente glicanica [2]: la glicolilazione del MurNAC
Osservata inizialmente in ceppi di M. smegmatis Rappresenta un marker di molti generi batterici appartenenti al phylum degli Actinobatteri (Gordonia, Nocardia, Rhodococcus) L’aggiunta di un ulteriore gruppo idrossile si realizza durante il pathway biosintetico del PGN
![Modificazioni nella componente glicanica [2]: la glicolilazione del MurNAC](http://slideplayer.it/slide/2348156/8/images/33/Modificazioni+nella+componente+glicanica+%5B2%5D%3A+la+glicolilazione+del+MurNAC.jpg "Osservata inizialmente in ceppi di M. smegmatis. Rappresenta un marker di molti generi batterici appartenenti al phylum degli Actinobatteri (Gordonia, Nocardia, Rhodococcus) L’aggiunta di un ulteriore gruppo idrossile si realizza durante il pathway biosintetico del PGN.")
34
Qual è il ruolo biologico della glicolilazione del PGN?
Il gruppo idrossilico sembra partecipare alla formazione di nuovi legami idrogeno con componenti della parete, conferendo maggiore stabilità e resistenza alla parete stessa Mutanti di M smegmatis nel gene namH sono più suscettibile all’azione del lisozima e degli antibiotici b-lattamici

35
Modificazioni nella componente glicanica [3]: l’acetilazione del MurNAC
Aggiunta di un secondo gruppo idrossilico al C6 con formazione di un residuo di diacetil muramico L’acetilazione del residuo di MurNAC è la modificazione che si presenta con maggiore frequenza nei Gram positivi e negativi Il grado di acetilazione del residuo di MurNAC varia considerevolmente tra le specie batteriche e tra i diversi ceppi batterici (dal 20 al 70%)
![Modificazioni nella componente glicanica [3]: l’acetilazione del MurNAC](http://slideplayer.it/slide/2348156/8/images/35/Modificazioni+nella+componente+glicanica+%5B3%5D%3A+l%E2%80%99acetilazione+del+MurNAC.jpg "Aggiunta di un secondo gruppo idrossilico al C6 con formazione di un residuo di diacetil muramico. L’acetilazione del residuo di MurNAC è la modificazione che si presenta con maggiore frequenza nei Gram positivi e negativi. Il grado di acetilazione del residuo di MurNAC varia considerevolmente tra le specie batteriche e tra i diversi ceppi batterici (dal 20 al 70%)")
37
Modificazioni nella componente glicanica [3]: l’acetilazione del MurNAC
Gene oatA (O-acetil transferasi) di S. aureus e adrA di S. pneumoniae Enzimi transmembrana (11 regioni predette transmembrana) con la regione carbossi-terminale extra-citoplasmatica con attività catalitica
![Modificazioni nella componente glicanica [3]: l’acetilazione del MurNAC](http://slideplayer.it/slide/2348156/8/images/37/Modificazioni+nella+componente+glicanica+%5B3%5D%3A+l%E2%80%99acetilazione+del+MurNAC.jpg "Gene oatA (O-acetil transferasi) di S. aureus e adrA di S. pneumoniae. Enzimi transmembrana (11 regioni predette transmembrana) con la regione carbossi-terminale extra-citoplasmatica con attività catalitica.")
38
Qual è il ruolo biologico dell’acetilazione del PGN?
Maggiore resistenza all’azione del lisozima Negli Staphilococchi sembra esserci una chiara correlazione tra patogenicità, resistenza all’attività muramidasica del lisozima e grado di acetilazione del MurNAC Persistenza di frammenti di PGN con residui di MurNAC acetilati: modulazione del riconoscimento dei patogeni da parte del sistema immunitario innato Il grado di acetilazione dei residui di MurNAC sembra essere ridotto in seguito al trattamento con antibiotici b-lattamici (Neisseria spp., S. aureus, P. mirabilis) In N. gonorrhoeae esiste una proporzionalità diretta tra il grado di acetilazione del MurNAC e l’attività della carbossipeptidasi PBP2
 In N. gonorrhoeae esiste una proporzionalità diretta tra il grado di acetilazione del MurNAC e l’attività della carbossipeptidasi PBP2.")
39
Il Peptidoglicano (PGN): [5]
La componente peptidica rappresenta il punto di ancoraggio di proteine della parete cellulare al PGN: nei batteri Gram negativi, la lipoproteina di Brown CH2 CH CH2 S Cys O NH (aa)56 Lys NH2 COOH PEPTIDOGLICANO XXXXXXXXXXXXXX GlcNAc MurNAc XXXXXXXXXXXXXXXXXX D-Ala L-Ala D-Glu m-Dap NH2 Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [5]](http://slideplayer.it/slide/2348156/8/images/39/Il+Peptidoglicano+%28PGN%29%3A+%5B5%5D.jpg "La componente peptidica rappresenta il punto di ancoraggio di proteine della parete cellulare al PGN: nei batteri Gram negativi, la lipoproteina di Brown. CH2 CH CH2 S Cys. O. NH. (aa)56. Lys NH2. COOH. PEPTIDOGLICANO. XXXXXXXXXXXXXX GlcNAc MurNAc XXXXXXXXXXXXXXXXXX. D-Ala. L-Ala. D-Glu. m-Dap NH2. Esclusivo dei batteri…bersaglio degli antibiotici.")
40
Il Peptidoglicano (PGN): [6]
La componente peptidica rappresenta il punto di ancoraggio di proteine della parete cellulare al PGN: nei batteri Gram negativi, la lipoproteina di Brown Il legame della BLP al PGN avviene attraverso reazioni di transpeptidazione Nei batteri Gram positivi ancorate al PGN numerose componenti. Le reazioni di ancoraggio sono catalizzate dalla SortasiA Esclusivo dei batteri…bersaglio degli antibiotici
![Il Peptidoglicano (PGN): [6]](http://slideplayer.it/slide/2348156/8/images/40/Il+Peptidoglicano+%28PGN%29%3A+%5B6%5D.jpg "La componente peptidica rappresenta il punto di ancoraggio di proteine della parete cellulare al PGN: nei batteri Gram negativi, la lipoproteina di Brown. Il legame della BLP al PGN avviene attraverso reazioni di transpeptidazione. Nei batteri Gram positivi ancorate al PGN numerose componenti. Le reazioni di ancoraggio sono catalizzate dalla SortasiA. Esclusivo dei batteri…bersaglio degli antibiotici.")
41
Metabolismo del PGN Gram positivi: le unità neosintetizzate vengono inserite nel versante interno, prima che la degradazione possa iniziare (INSIDE TO OUTSIDE) Gram negativi: tre nuovi filamenti vengono inseriti contemporaneamente alla rimozione di un filamento vecchio (THREE FOR ONE) Nei g+ la parete non è mai indebolita in quanto i klegami covalenti si formano prima della rimozione degli strati più esterni
 Gram negativi: tre nuovi filamenti vengono inseriti contemporaneamente alla rimozione di un filamento vecchio (THREE FOR ONE) Nei g+ la parete non è mai indebolita in quanto i klegami covalenti si formano prima della rimozione degli strati più esterni.")
42
“Three for one”

43
Complesso “Yin-Yang” Complesso multienzimatico che combina l’attività di: Transglicosilasi litiche (LT) Endopeptidasi (EP) Amidasi (AM) Transpeptidasi (TP) Transglicosilasi (TG)
 Amidasi (AM) Transpeptidasi (TP) Transglicosilasi (TG)")
44
Processo di recycling nei Gram-
Durante la crescita batterica, circa il 40-50% del PGN viene degradata nel corso di ogni generazione I prodotti di turnover si accumulano nello spazio periplasmatico Il 90% dei prodotti di turnover viene riciclato e reincorporato nel sacculo stesso Il 10% del materiale degradato viene rilasciato nell’ambiente Per coli è stato dimostrato che…. Il recycling è un processo attraverso il quale la cellula batterica riutilizza e modifica i prodotti del turnover reinserendoli nella via biosintetica del PGN Nei Gram Positivi circa il 70% del vecchio PGN viene liberato all’esterno durante la crescita e solo una piccola parte entra nella via del recycling

45
“Three for one”

46
Il PGN: gli enzimi del recycling
Membrana esterna , AmiB, AmiC Amidase SltY, MltA, MltB Lytic trasglycosylases AmiA PERIPLASMA MUREINA ANHYDRO MUROPEPTIDE TETRAPEPTIDE TRIPEPTIDE MppA periplasmic murein peptide binding AmpG muropeptide permease Opp oligopeptide permease Membrana interna CITOPLASMA NAG – anh-NAM – L-Ala – D-Glu – meso-DAP – D-Ala UDP-NAM – pentapeptide MurF D-Ala – D-Ala UDP-NAM – D-Ala – L-Glu – meso-DAP Mpl murein peptide ligase UDP-NAM La via del recycling è costituita da una serie di enzimi localizzati nel periplasma, a livello della membrana plasmatica e nel citoplasma Periplasma: enzimi che degradano lo strato di PGN per permettere sia l’inserimento di nuovo materiale sintetizzato che il rimodellamento continuo dello strato di PGN Membrana: permeasi che mediano il passaggio dei muropeptidi Citoplasma: enizmi che rimodellano NagZ Β-N-acetyl-glucosamidase AmpD anhMurNAc-Lala amidase LdcA L,D-carboxypeptidase L-Ala – D-Glu meso-DAP L-Ala – D-Glu – meso-DAP – D-Ala LdcA L,D-carboxypeptidase MpaA Murein peptie amidase L-Ala – D-Glu-meso-DAP

Presentazioni simili