Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
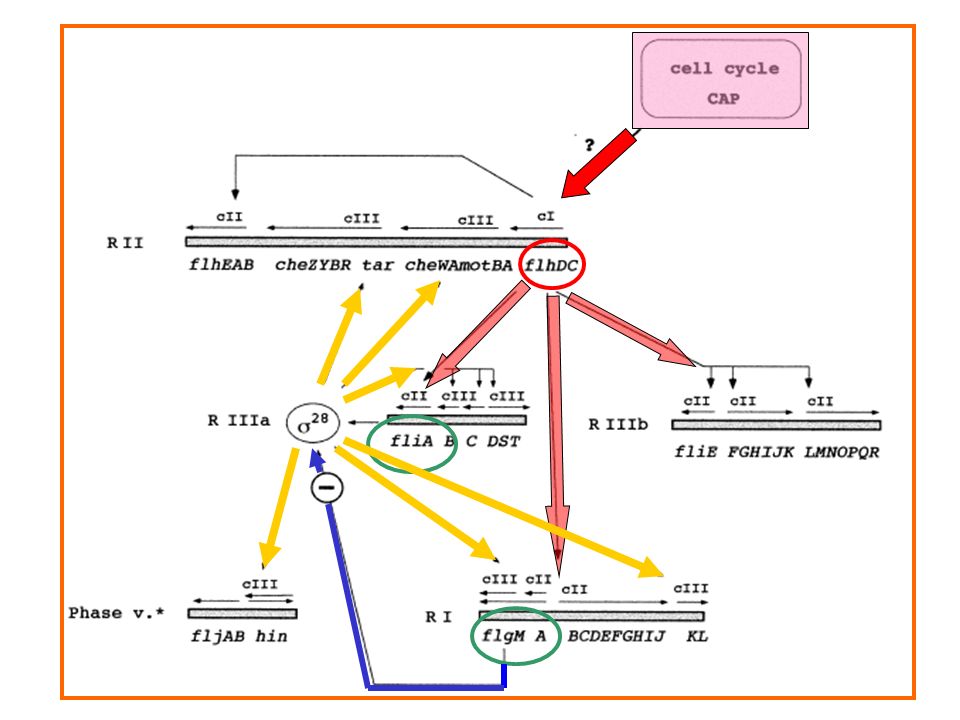
Il processo di CHEMIOTASSI

2
Two-component and chemotaxis signalling in bacteria
Two-component and chemotaxis signalling in bacteria. (A) Schematic representation of the canonical two-component system. Sensory histidine kinase (HK) is composed of the input (blue) and the autokinase (red) domains; the kinase is typically a dimer. The response regulator (RR) consists of the receiver (purple) and output (green) domains. The phosphate group is transferred from the histidine residue on the kinase to the asparate residue on the response regulator, activating the output domain, which typically regulates gene expression. The response regulator can be dephosphorylated by the phosphatase activity of the kinase. (B) Molecular composition of the chemotaxis pathway in E. coli. Receptors sense and transmit signals to regulate the activity of the cytoplasmic histidine kinase CheA. Receptors form trimers of dimers, where different types of receptors (light or dark blue) are mixed. CheA binding and regulation by receptors are aided by CheW. CheA transfers phosphate group to CheY, the single-domain response regulator controlling flagellar motor, and to CheB, composed of the regulatory receiver domain and the output methylesterase domain. Receptors are methylated on glutamate residues by the methyltransferase CheR. CheY is dephosphorylated by the phosphatase CheZ. Receptors, CheW, CheA and CheZ form a stable signalling core, to which CheR, CheB and CheY dynamically localize.
 Schematic representation of the canonical two-component system. Sensory histidine kinase (HK) is composed of the input (blue) and the autokinase (red) domains; the kinase is typically a dimer. The response regulator (RR) consists of the receiver (purple) and output (green) domains. The phosphate group is transferred from the histidine residue on the kinase to the asparate residue on the response regulator, activating the output domain, which typically regulates gene expression. The response regulator can be dephosphorylated by the phosphatase activity of the kinase. (B) Molecular composition of the chemotaxis pathway in E. coli. Receptors sense and transmit signals to regulate the activity of the cytoplasmic histidine kinase CheA. Receptors form trimers of dimers, where different types of receptors (light or dark blue) are mixed. CheA binding and regulation by receptors are aided by CheW. CheA transfers phosphate group to CheY, the single-domain response regulator controlling flagellar motor, and to CheB, composed of the regulatory receiver domain and the output methylesterase domain. Receptors are methylated on glutamate residues by the methyltransferase CheR. CheY is dephosphorylated by the phosphatase CheZ. Receptors, CheW, CheA and CheZ form a stable signalling core, to which CheR, CheB and CheY dynamically localize.")
4
Variazione della concentrazione di microrganismi in un capillare in presenza di attraente o repellente

5
Movimento dei microrganismi verso un alga unicellulare che produce per fotosintesi ossigeno sostanza attraente per questi batteri

7
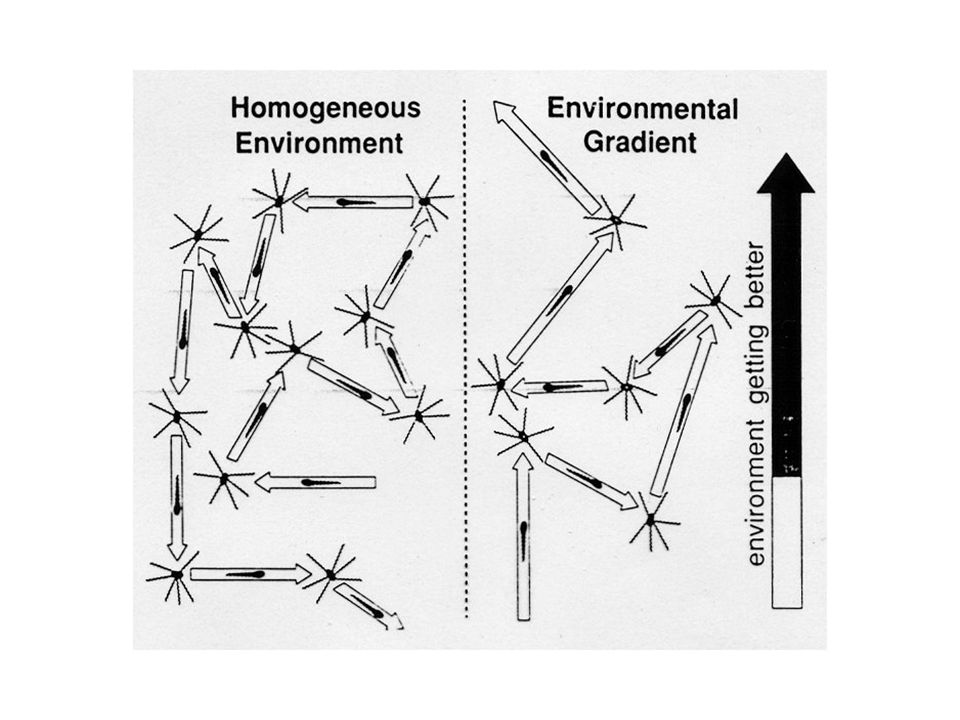
L’alternarsi tra le fasi di rotazione antiorarie ed orarie e la loro frequenza determina un orientamento generale

8
Il movimento dei batteri
Il batterio risponde ad un gradiente temporale registrando la concentrazione precedente e comparandola con l’attuale In assenza di un nutriente chemio-attraente i movimenti sono causali. In presenza di un nutriente chemio-attraente le cellule si orientano nella direzione del gradiente aumentando i moviementi lineari

11
Schematic representation of an E
Schematic representation of an E. coli cell swimming in a direction and responding to an increasing concentration of a chemical with a tumbling and a consequent change of direction. Tumbling is stimulated by a temporary accumulation ofCheY-P molecules (blue; unphosphorylated CheY are represented in purple), which communicate to the perithrichious flagellar motors to switch the sense of rotation.
, which communicate to the perithrichious flagellar. motors to switch the sense of rotation.")
12
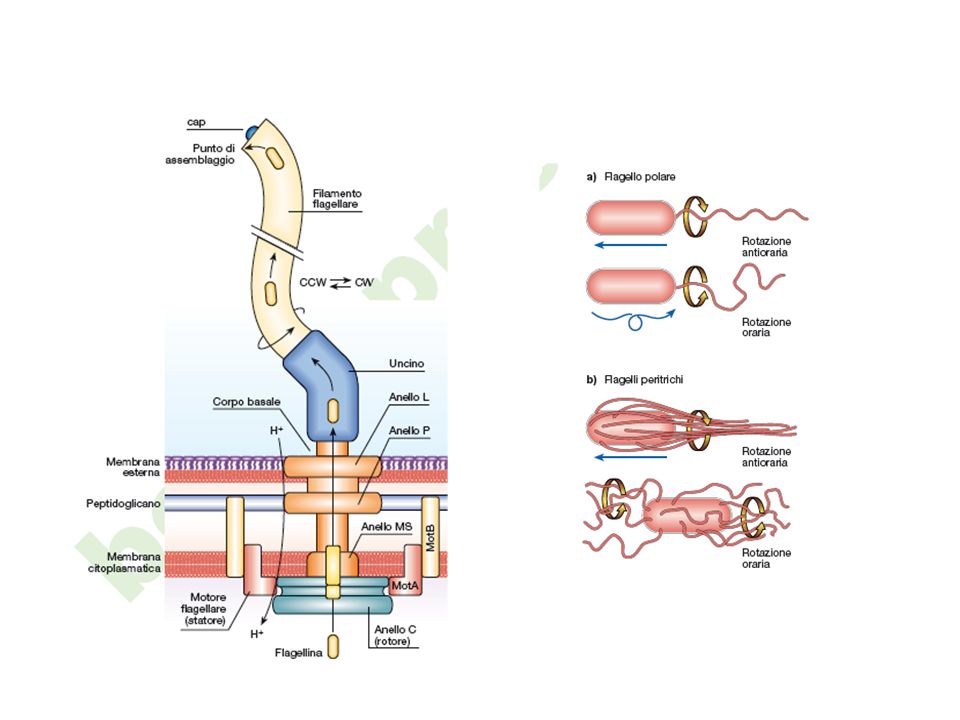
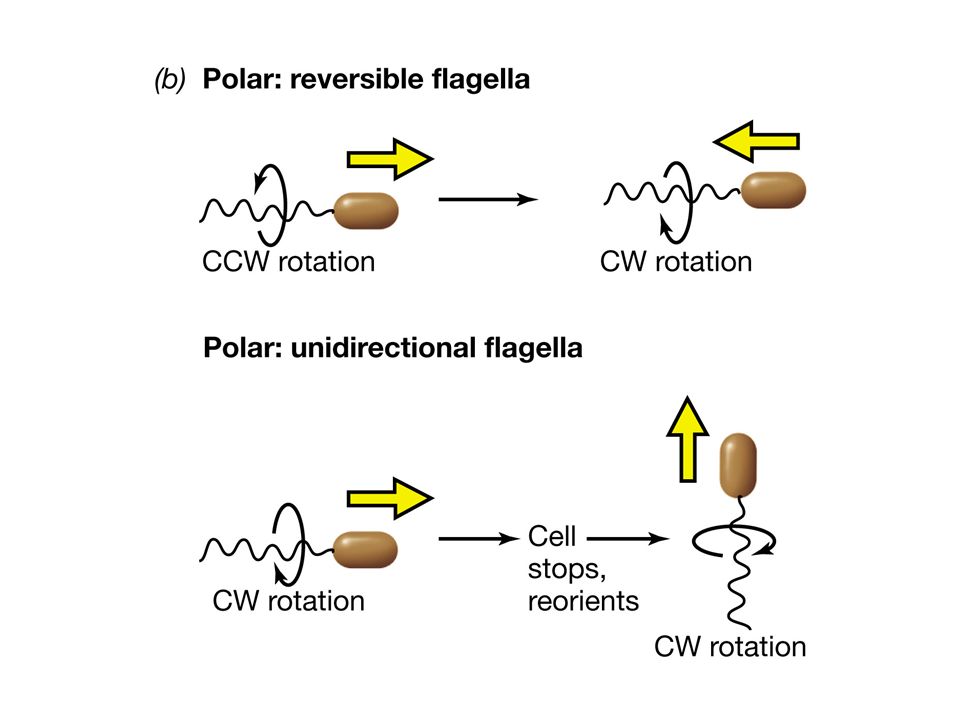
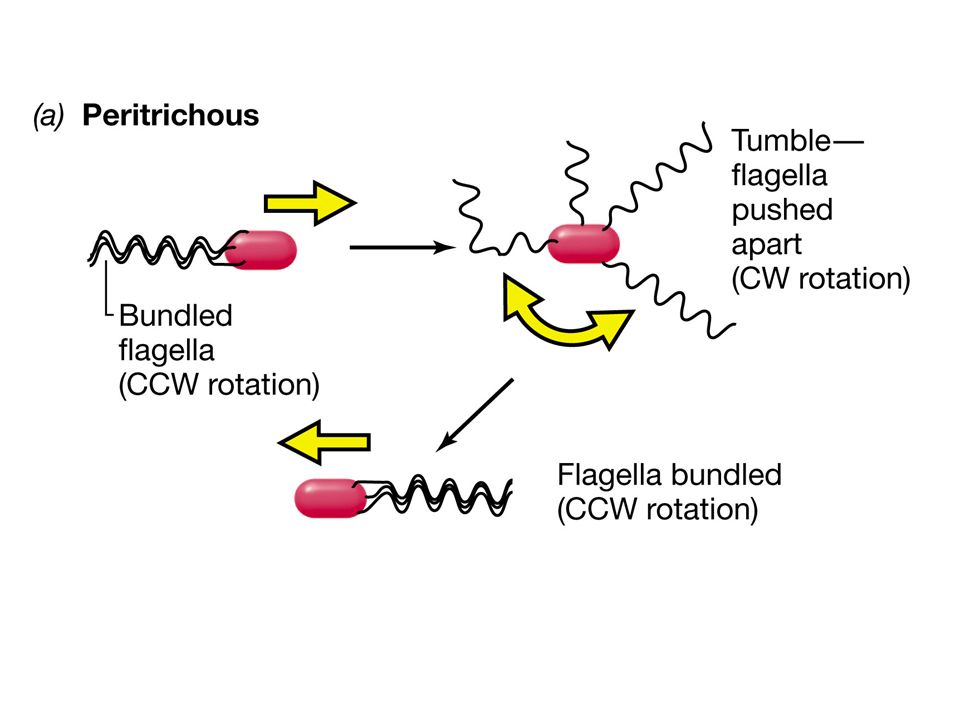
Flagelli ruotano in senso antiorario
Movimento rettilineo Flagelli ruotano in senso orario CAPRIOLA (thumble)
")
13
L’esempio meglio studiato e compreso di trasduzione del segnale e risposta cellulare è quello della chemiotassi. Nella chemiotassi si osserva come un batterio dotato di flagelli “proceda” in risposta a gradienti di molecole attraenti o molecole repellenti modificando la natura del movimento del flagello. Il flagello che ruota in senso antiorario determina un andamento rettilineo, il flagello che ruota in senso orario provoca invece il ribaltamento della cellula.

14
Il segnale, di tipo chimico, viene trasdotto attraverso la membrana, dal sensore e tradotto in segnale chimico come fosforilazione Il segnale, sottoforma di fosforilazione, viene trasferito al regolatore che, modificato strutturalmente esercita il suo ruolo

15
La trasduzione del segnale nei batteri
Il sistema a due componenti Percezione dello stimolo e movimento

17
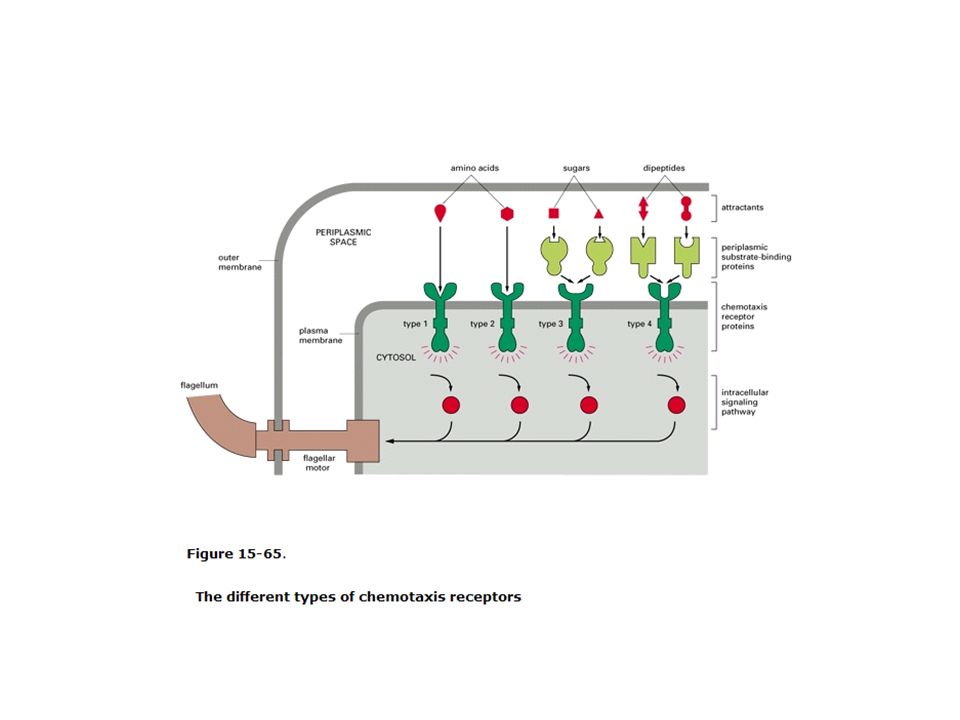
Struttura delle proteine MCP ( metil -accepting proteins for chemiotaxis)
")
19
Le MCP sono proteine della membrana INTERNA con un dominio rivolto verso lo spazio periplasmatico

20
Le MCP (Methyl-accepting protein) sono suddivise in 4 gruppi e presentano probabilmente oltre 2000 differenti proteine per gruppo. Tipo I: serina ed alcuni repellenti Il tipo II: maltosio aspartato e alcuni repellenti Tipo III: galattosio e ribosio Tipo IV: alcuni dipeptidi

21
Esistono 8 siti di metilazione a carico di ogni dimero di MCP
Esistono 8 siti di metilazione a carico di ogni dimero di MCP. La riduzione dell’attraente determina la demetilazione delle MCP da parte di CheB-P

22
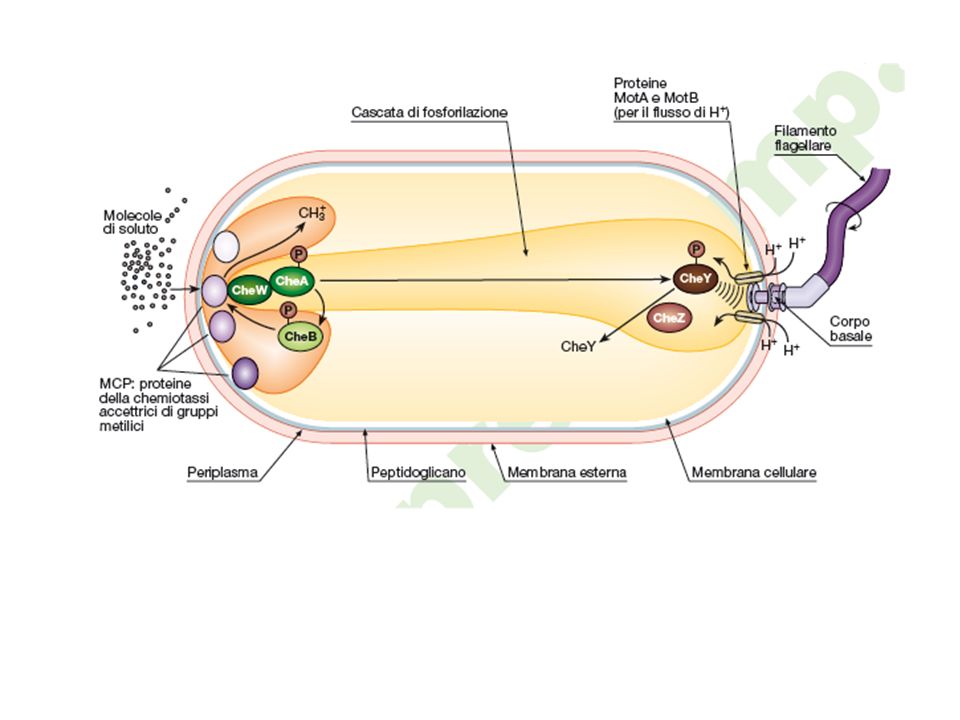
Rivediamo la cascata di regolazione che porterà al movimento come un circuito che riceve un segnale ( input ) e genera una risposta (output) tramite un sensore (MCP) ed un attuatore ( la fosforilazione CheA mediata)
 e genera una risposta (output) tramite un sensore (MCP) ed un attuatore ( la fosforilazione CheA mediata)")
23
La proteina citoplasmatica CheR continua lentamente ad aggiungere gruppi metilici alle MCP utilizzando la S-adenilmetosina come donatore di metili. Il livello di metilazione delle MCP influenza la loro conformazione e controlla l’adattamento a un determinato segnale. Se il livello di attraente si mantiene elevato il livello di fosforilazione di CheA si manterrà basso e la cellula manterrà il moto di avanzamento regolare. Il livello di metilazione delle MCP aumenterà perché vi è poca CheA-P ( e quindi poco CheY-P e CheB-P)
")
24
I trasduttori MCP sono a contatto con le proteine citoplasmatiche CheW e CheA.
Le MCP però ad un certo punto saranno completamente metilate e questo determina un cambiamento conformazionale che si ripercuote sulla fosforilazione di CheA ( CheA-P). CheA è una chinasi sensore che cambia conformazione coadiuvata da CheW quando il trasduttore si lega ad un attraente. Che-A si autofosforila CheA-P
. CheA è una chinasi sensore che cambia conformazione coadiuvata da CheW quando il trasduttore si lega ad un attraente. Che-A si autofosforila CheA-P.")
25
CheA è una chinasi in grado di autofosforilarsi in grado di fosforilare CheY - CheY-P in grado di fosforilare CheB - CheB-P (demetilasi)
 .")
26
CheA-P può fosforilare CheY (CheY-P) regolatore della risposta che interagisce con il motore del flagello solo se fosforilato determinando la rotazione in senso orario. CheZ può defosforilare CheY-P ( CheY) e quindi il movimento può procedere perché il flagello ruoterà in senso antiorario
 e quindi il movimento può procedere perché il flagello ruoterà in senso antiorario.")
27
CheR è una metil tranferasi in grado di metilare le MCP nei residui di glutammato aumentando cosi l’attività chinasica di CheA. Il livello di metilazione della MCP fornisce al batterio una misura del passato gradiente in quanto la metilazione avvine solo se vi è legame tra MCP e molecola attraente. Seguendo il pathway verde si arriva al movimento orario ( capriola) mentre il rosso indica movimento antiorario ( lineare)
 mentre il rosso indica movimento antiorario ( lineare)")
28
CheA fosforilata CheA-P
fosforila CheY che si lega al motore del flagello

29
Ruolo delle istidin protein chinasi ( HPK) e dei regolatori della risposta con dominio aspartato (RR) nel sistema di trasduzione del segnale basato sul fosforilazione istidina aspartato (HAP)
 e dei regolatori della risposta con dominio aspartato (RR) nel sistema di trasduzione del segnale basato sul fosforilazione istidina aspartato (HAP)")
30
Some of the different combinations of histidine protein kinase (HPK) and aspartate response regulator (RR) domains in histidine–aspartate phosphorelay (HAP) systems. a | The EnvZ/OmpR pathway of Escherichia coli, which is involved in regulating the expression of the two outer-membrane porins OmpF and OmpC. A membrane-bound HPK (EnvZ) controls the activity of the RR OmpR in response to changes in osmolarity. b | The complex ArcB–ArcA HAP system of E. coli. The membrane-bound HPK ArcB senses changes in the redox state of components of the respiratory electron-transport chain through its PAS (PER, ARNT, SIM) domain. The phosphoryl group is then passed from the conserved His in the ArcB kinase domain to a fused RR domain, then to a fused histidine-containing phosphotransfer (HPt) domain and finally to a DNA-binding RR ArcA. ArcA regulates microaerophilic gene expression. c | The chemosensory pathway of E. coli. The soluble HPK chemotaxis protein (Che)A has five domains per monomer that are designated P1–P5 from the N terminus to the C terminus. CheA senses changes through transmembrane chemoreceptors, which induce the trans-autophosphorylation of dimeric CheA on a His residue of the HPt domain. Two RRs compete for this phosphoryl group: CheY, a single-domain, motor-binding protein, which controls flagellar motor switching, and CheB, which controls the adaptation of the chemoreceptors. d | Part of the complex system that regulates sporulation in Bacillus subtilis. A single-domain RR, Spo0F, is regulated by two HPKs, one of which has numerous transmembrane domains (KinB), the other of which is soluble with numerous PAS domains (KinA). Spo0F indirectly phosphorylates a DNA-binding RR, Spo0A, by way of a His residue in Spo0B. Throughout this figure, light-green rectangles highlight conserved, phosphorylatable His residues, light-blue rectangles highlight conserved, phosphorylatable Asp residues, and orange circles highlight phosphoryl (P) groups. NGFG represents the kinase domain and, with the exception of CheA, the conserved His residue that precedes the kinase domain is contained within the dimerization domain. Despite being dimeric in nature, HPKs are shown here as monomers for simplicity, and the HAMP domain ('histidine kinases, adenylyl cyclases, methyl-binding proteins and phosphatases' domain) is a linker domain. Download file

31
CheA-P può fosforilare anche la proteina CheB
CheB-P svolge un ruolo di demetilasi sulle MCP cruciale per il fenomeno dell’adattamento. A questo punto la fosforilazione di CheB-P riporta le MCP al loro livello iniziale di metilazione ( molto basso ) e quindi può riprendere il movimento rettilineo. I repellenti aumentano il livello di CheY fosforilata e quindi provocano il susseguirsi di capriole
 e quindi può riprendere il movimento rettilineo. I repellenti aumentano il livello di CheY fosforilata e quindi provocano il susseguirsi di capriole.")
32
Schematic representation of the E. coli chemosensory system
Schematic representation of the E. coli chemosensory system. Trasmembrane receptors (Mcp) perceive signals directly (red flash) or via ligand binding proteins (LBP).). CheR and CheB proteins (light or dark green) add to or remove from Mcps metil groups.
 perceive signals directly (red. flash) or via ligand binding proteins (LBP).). CheR and CheB proteins (light or dark green) add to or remove from Mcps metil groups.")
33
ADATTAMENTO Quando il batterio si trova in concentrazioni elevate di attraente può saturare le MCP. In questo caso tali proteine non sarebbero in grado di indicare ulteriori variazioni delle concentrazioni delle molecole nell’ambiente. Questo fenomeno è impedito dall’adattamento. Quando un attraente si lega ad una MCP ne determina una alterazione strutturale. La proteina CheB-P, demetilando le MCP le riporta alla struttura originaria adattandola alle nuove concentrazioni di attraente

36
CheR transfers methyl groups from S-
adenosylmethionine (AdoMet) to conserved membrane receptors. The methyltransferase The changes in attractant or repellent glutamate residues on the cytoplasmic signaling domains of the chemotaxis the phosphoenolpyruvate (PEP)-dependent phosphotransferase system (PTS) binds and phosphorylation by CheA. Enzyme I (Pts I) of these groups when activated through receptors. The methylesterase CheB removes chain. The chemotaxis system can adapt to concentrations by covalently modifying the indirectly via periplasmic binding proteins. The to their respective receptors either directly or thought to interact with the electron transport molecules diffuse through the outer chemotaxis receptors signal through the inner membrane into the periplasm, where they bind membrane into the cytoplasm, where they through its flavin-binding domain, and is apparently receives intracellular redox signals interact with the adaptor protein CheW and the kinase CheA. The aerotaxis receptor, Aer, phosphorylation of the response regulator CheY, which directly interacts with the E. coli chemotaxis system. Stimulus transported into the cell. All chemotaxis inhibits CheA when a variety of hexoses are signals are ultimately integrated at the level of The E. colic hemotaxis system. Stimulimolecules diffuse through the outermembrane into the periplasm, where they bindto their respective receptors either directly orindirectly via periplasmic binding proteins. Thechemotaxis receptors signal through the innermembrane into the cytoplasm, where theyinteract with the adaptor protein CheW andthe kinase CheA. The aerotaxis receptor, Aer,apparently receives intracellular redox signalsthrough its flavin-binding domain, and isthought to interact with the electron transportchain. The chemotaxis system can adapt to changes in attractant or repellentconcentrations by covalently modifying the membrane receptors. The methyltransferaseCheR transfers methyl groups from S-adenosylmethionine (AdoMet) to conserved glutamate residues on the cytoplasmicsignaling domains of the chemotaxisreceptors. The methylesterase CheB removesthese groups when activated throughphosphorylation by CheA. Enzyme I (Pts I) of the phosphoenolpyruvate (PEP)-dependentphosphotransferase system (PTS) binds and inhibits CheA when a variety of hexoses aretransported into the cell. All chemotaxis signals are ultimately integrated at the level ofphosphorylation of the response regulatorCheY, which directly interacts with the flagellar motor.
 to conserved. membrane receptors. The methyltransferase. The. changes in attractant or repellent. glutamate residues on the cytoplasmic. signaling domains of the chemotaxis. the phosphoenolpyruvate (PEP)-dependent. phosphotransferase system (PTS) binds and. phosphorylation by CheA. Enzyme I (Pts I) of. these groups when activated through. receptors. The methylesterase CheB removes. chain. The chemotaxis system can adapt to. concentrations by covalently modifying the. indirectly via periplasmic binding proteins. The. to their respective receptors either directly or. thought to interact with the electron transport. molecules diffuse through the outer. chemotaxis receptors signal through the inner. membrane into the periplasm, where they bind. membrane into the cytoplasm, where they. through its flavin-binding domain, and is. apparently receives intracellular redox signals. interact with the adaptor protein CheW and. the kinase CheA. The aerotaxis receptor, Aer, phosphorylation of the response regulator. CheY, which directly interacts with the. E. coli. chemotaxis system. Stimulus. transported into the cell. All chemotaxis. inhibits CheA when a variety of hexoses are. signals are ultimately integrated at the level of. The E. colic hemotaxis system. Stimulimolecules diffuse through the outermembrane into the periplasm, where they bindto their respective receptors either directly orindirectly via periplasmic binding proteins. Thechemotaxis receptors signal through the innermembrane into the cytoplasm, where theyinteract with the adaptor protein CheW andthe kinase CheA. The aerotaxis receptor, Aer,apparently receives intracellular redox signalsthrough its flavin-binding domain, and isthought to interact with the electron transportchain. The chemotaxis system can adapt to changes in attractant or repellentconcentrations by covalently modifying the membrane receptors. The methyltransferaseCheR transfers methyl groups from S-adenosylmethionine (AdoMet) to conserved. glutamate residues on the cytoplasmicsignaling domains of the chemotaxisreceptors. The methylesterase CheB removesthese groups when activated throughphosphorylation by CheA. Enzyme I (Pts I) of the phosphoenolpyruvate (PEP)-dependentphosphotransferase system (PTS) binds and inhibits CheA when a variety of hexoses aretransported into the cell. All chemotaxis signals are ultimately integrated at the level ofphosphorylation of the response regulatorCheY, which directly interacts with the flagellar motor.")
37
Modello generale della regolazione della chemiotassi

38
Meccanismo di regolazione della chemiotassi
Repellente Le MCP completamente metilate rispondono meglio ai repellenti

39
Il motore del flagello visto dall’interno della cellula
CheY-P si lega a FliM provocando la rotazione in senso orario ( capriola)
")
40
FOTOTASSIA Il movimento dei microrganismi verso la luce per ottimizzare il processo di fotosintesi. Con uno spettro di luce orientato verso un vetrino si può osservare come i batteri si accumulino in corrispondenza della lunghezza d’onda ideale per l’assorbimento da parte dei loro pigmenti fotosintetici

41
La fototassi è un movimento attivo verso un gradiente di
aumentata intensità della luce. Colonie di Rhodospirillum che si muovono all’unisono verso la luce

42
I componenti del sistema regolativo coinvolti nella chemiotassi sono probabilmente coinvolti anche nella fototassi. Mutanti difettivi nella fototassi sono difettivi anche nella chemiotassi Fotorecettore analogo ai chemiorecettore (MCP) potrebbe funzionare come sensore della luce. Fotorecettore interagisce con proteine che agiscono sul motore flagellare al fine di mantenere le cellule in uno stato di avanzamento verso zone di maggiore intensità di luce.
 potrebbe funzionare come sensore della luce. Fotorecettore interagisce con proteine che agiscono sul motore flagellare al fine di mantenere le cellule in uno stato di avanzamento verso zone di maggiore intensità di luce.")
43
SCOTOFOBOTASSI Fenomeno osservabile in laboratorio quando un batterio esce dal campo illuminato e si dirige verso il buio. L’entrata nel buio si ripercuote negativamente sullo stato energetico della cellula e le segnala di capovolgersi e di riprendere il moto rettilineo verso la sorgente di luce.

44
Movimento per scivolamento
è ampiamente diffuso , Cianobatteri Mixobatteri ( Batteri Gram-) non richiede flagello avviene lungo l’asse maggiore del microrganismo. I batteri che si muovono per scivolamento sono bastoncellari o filamentosi La colonia assume una morfologia particolare in quanto ogni cellula può allontanarsi dal centro della colonia
 non richiede flagello. avviene lungo l’asse maggiore del microrganismo. I batteri che si muovono per scivolamento sono bastoncellari o filamentosi. La colonia assume una morfologia particolare in quanto ogni cellula può allontanarsi dal centro della colonia.")
45
Nei cianobatteri il movimento per scivolamento è accompagnato dalla secrezione dello SLIME, una sostanza mucosa di natura polisaccaridica. Lo SLIME ( lett. BAVA) man mano che viene secreto aderisce alla superficie e il batterio scivola su questa sostanza viscida da lui secreta.
 man mano che viene secreto aderisce alla superficie e il batterio scivola su questa sostanza viscida da lui secreta.")
46
Lo scivolamento in Flavobacterium non è indotto dalla escrezione di slime.
Le proteine specifiche per la motilità sono localizzate sia nella membrana interna che esterna. Colonie mutanti

47
Movimento a RUOTA DENTATA
L’energia rilasciata dalla forza protonmotrice permette il movimento delle proteine della I.M. che si trasmette a quelle della O.M. Il movimento delle proteine della O.M. avviene in direzione opposta rispetto al movimento che effettuerà la cellula

48
Modello di scivolamento di Flavobacterium johnsoniae
Lo scivolamento è indotto da un movimento delle proteine della superficie della cellula. Si crea un contatto tra le proteine della IM e della OM Le proteine specifiche per la motilità sono ancorate nella membrana citoplasmatica e nella membrana esterna e sospingono le cellule in avanti e indietro con un meccanismo a ruota dentata

50
Svolgono importanti funzioni nella cellula
I pili sono strutture proteiche non flagellari presenti sulla superificie delle cellule batteriche Svolgono importanti funzioni nella cellula quali adesione a superfici, formazione di biofilm. Sono suddivisi in base al tipo di meccanismo di formazione: Pili di tipo I ( Chaperon/Usher) Pili di tipo IV. I pili di tipo IV sono molto diffusi ed importanti e mediano oltre ai processi di adesione anche una particolare forma di movimento cellulare detto twitching o movimento a scatto . Pili e fimbrie sono designati ad indicare la medesime strutture, fimbrie sono più corte, pili più lunghi ma i due termini si usano spesso indifferentemente
 Pili di tipo IV. I pili di tipo IV sono molto diffusi ed importanti e mediano oltre ai processi di adesione anche una particolare forma di movimento cellulare detto twitching o movimento a scatto . Pili e fimbrie sono designati ad indicare la medesime strutture, fimbrie sono più corte, pili più lunghi ma i due termini si usano spesso indifferentemente.")
51
Pili di tipo IV ed il movimento a scatti
Sono costituiti dal ripetersi di subunità di pilina PilA che viene sintetizzata in forma di precursore con la sequenza segnale all’estremità N -terminale. La sequenza segnale verrà rimossa dalla Prepilina peptidasi (PilD) omologo della peptidasi che rimuove la sequenza segnale della preflagellina negli Archea. La regione N-terminale della pilina forma una elica idrofobica mentre la porzione C terminale idrofilica è rivolta nel periplsma. Nei batteri Gram- è presente un canale di membrane esterna PilQ che permette il passaggio del pilo.
 omologo della peptidasi che rimuove la sequenza segnale della preflagellina negli Archea. La regione N-terminale della pilina forma una elica idrofobica mentre la porzione C terminale idrofilica è rivolta nel periplsma. Nei batteri Gram- è presente un canale di membrane esterna PilQ che permette il passaggio del pilo.")
52
Ruolo fondamentale viene svolto dalle 2 ATPasi PilB e PilT con funzione antagonista. L’ATPasi di assemblaggio PilB dopo idrolisi di ATP spinge il pilo verso l’esterno creando un vuoto che viene riempito dalle nuove subunità di pilina. L’idrolisi di ATP mediata da PilT invece determina la rimozione dei monomeri di pilina dalla base del pilo creando retrazione del pilo.

53
In seguito a cicli di estensione e ritrazione alcuni batteri si possono muovere a scatti grazie ai Pili di Tipo IV presenti sulla loro superficie. Le subunità di pilina eliminate durante la ritrazione possono poi essere riutilizzate per il successivo processo di estensione

54
. Vi sono diverse omologie tra il pilo di tipo IV ed il flagello degli Archea

55
E’ possibile vedere l’allungamento e la ritrazione dei pili di Tipo IV

56
Myxococcus xanthus ed il movimento a scatti
Un esempio interessante di movimento a scatti è rappresentato da Myxococcus xanthus, un microrganismo del suolo. Questo microrganismo è in grado di muoversi per scivolameneto utilizzando i pili di tipo IV che si allungano e ritraggono. Ogni 6 –8 minuti M.xanthus è in grado di revertire il movimento cambiando cosi direzione.

58
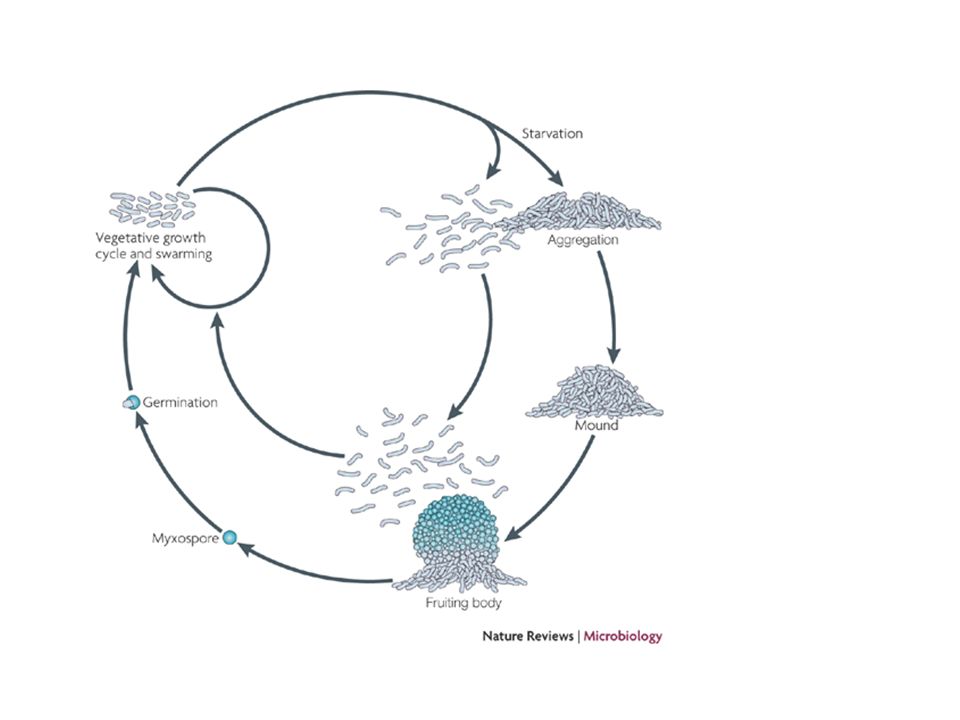
Life cycle of M. xanthus. (Vegetative growth) On a solid surface with soluble nutrients, groups of M. xanthus cells (swarms) grow, divide, and move outward. On a solid surface in the presence of lysing cells or prey, M. xanthus cells form “accordion waves” known as ripples. (Low-nutrient development) On a solid surface upon nutrient step-down or starvation, 105 to 106 cells aggregate to form mounds and then fruiting bodies. The rod-shaped cells in the fruiting bodies undergo morphogenesis and form spherical spores that are metabolically inactive but more resistant to desiccation and heat. Peripheral rods, a subpopulation of stressed cells, remain outside fruiting bodies in search of food. When nutrients become available, the spores germinate and complete the life cycle.
 On a solid surface with soluble nutrients, groups of M. xanthus cells (swarms) grow, divide, and move outward. On a solid surface in the presence of lysing cells or prey, M. xanthus cells form accordion waves known as ripples. (Low-nutrient development) On a solid surface upon nutrient step-down or starvation, 105 to 106 cells aggregate to form mounds and then fruiting bodies. The rod-shaped cells in the fruiting bodies undergo morphogenesis and form spherical spores that are metabolically inactive but more resistant to desiccation and heat. Peripheral rods, a subpopulation of stressed cells, remain outside fruiting bodies in search of food. When nutrients become available, the spores germinate and complete the life cycle..")
59
| Myxococcus xanthus cells move as groups by social motility (black arrow) and as single cells by adventurous motility (white arrow). Cells preferentially follow the paths of other cells. b | M. xanthus cells moving in ripples to consume Escherichia coli as prey11. Ripples have a wavelength of m.. c | Peripheral rods form a monolayer of cells between fruiting bodies that can move as reversing accordion waves d | A M. xanthus fruiting body on animal dung95. Fruiting bodies are 0.1–0.2 m in diameter. e | An optical slice of a fruiting body. Note that the spores are contained in the fruiting body and peripheral rods appear as a monolayer around the fruiting body.. f | A fruiting body that was cut to reveal the spores within. All scale bars are 15 m. Le cellule di Mixococcus xanthus si muovono come gruppi nella modalità sociale ( freccia nera) o come siLe cellule di ngole cellule nella modalità avventurosa( freccia bianca) Le cellule in genere seguono traccie di altre cellule.b) M.xanthuseffettuano un movimento ad onde per nutrirsi di cellule di E.coli Le onde hanno un ampiezza di 100 m.c) i batteri periferici formano
 o come siLe cellule di ngole cellule nella modalità avventurosa( freccia bianca) Le cellule in genere seguono traccie di altre cellule.b) M.xanthuseffettuano un movimento ad onde per nutrirsi di cellule di E.coli Le onde hanno un ampiezza di 100 m.c) i batteri periferici formano.")
61
Social motility is mediated by the extension and retraction of type IV pili (black tendrils) at the leading pole of a cell. Adventurous motility involves multiple transient adhesion complexes (coloured ovals on the bottom of cells) that are located throughout the length of a cell
 that are located throughout the length of a cell.")
62
(A)PilA è la subunità principale del pilo di tipo IV ed è localizzata al polo anteriore della cellula. E’ una proteina solubile che viene secreta attraverso il canale formato da PilQ. Tgl è una lipoproteina necessaria per il corretto assemblaggio del canale PilQ. Tgl svolge un ruolo cruciale in quanto si localizza solo ad un polo della cellula In mutanti Tgl-difettivi non si ha assemblaggio del pilo a causa della mancanza di canale PilQ. PilQ in questi mutanti si trova in forma monomerica ma può ripolimerizzare se il mutante Tgl – viene messo in contatto con cellule Tgl+.

63
(B) Quando la cellula cambia orientamento Tgl si sposta dall’altro polo , la traslocazione di Tgl è facilitata dal contatto cellula-cellula. (C)Una volta arrivato al polo opposto Tgl attiva il canale PilQ ed induce il trasporto e l’assemblaggio del filamento PilA. La presenza dei componenti disassemblati ad entrambi i poli facilita la riorganizzione del pilo all’altra estremità.
Una volta arrivato al polo opposto Tgl attiva il canale PilQ ed induce il trasporto e l’assemblaggio del filamento PilA. La presenza dei componenti disassemblati ad entrambi i poli facilita la riorganizzione del pilo all’altra estremità.")
64
Schematic representation of FrzCD-GFP clusters
stimulated by cell contacts and aligning. The stimulus generating from the cell contact is translated in the two adjacent cells and one or two of them will reverse the direction of movement in response.

65
Two dimeric chemoreceptors — methyl-accepting chemotaxis proteins (MCPs) — are shown, one of which is interacting with a periplasmic binding protein (PBP). In addition, two chemotaxis protein (Che)W monomers and a CheA dimer are shown interacting with the highly conserved signalling domain of the MCPs in the cytoplasm. It should be noted that, given the packing within MCP clusters and the calculations of the number of chemosensory proteins, the actual arrangement will be different. One CheA monomer will probably not interact with one MCP dimer, and instead a CheA dimer might span several receptors. A decrease in attractant concentration induces trans-autophosphorylation of the CheA dimer, which phosphorylates the response regulator CheY. Phosphorylated CheY then binds to the flagellar motor to bring about a change in direction. Phosphorylated CheA also phosphorylates another response regulator — the methylesterase CheB. Phosphorylated CheB competes with a constitutive methyltransferase, CheR, to control the degree of methylation of specific glutamates in the MCPs. This resets the signalling state of the receptors and allows them to adapt to the present concentration of attractant and to sense subsequent changes. The dephosphorylation of phosphorylated CheY is accelerated by the phosphatase CheZ. P, phosphoryl grou
W monomers and a CheA dimer are shown interacting with the highly conserved signalling domain of the MCPs in the cytoplasm. It should be noted that, given the packing within MCP clusters and the calculations of the number of chemosensory proteins, the actual arrangement will be different. One CheA monomer will probably not interact with one MCP dimer, and instead a CheA dimer might span several receptors. A decrease in attractant concentration induces trans-autophosphorylation of the CheA dimer, which phosphorylates the response regulator CheY. Phosphorylated CheY then binds to the flagellar motor to bring about a change in direction. Phosphorylated CheA also phosphorylates another response regulator — the methylesterase CheB. Phosphorylated CheB competes with a constitutive methyltransferase, CheR, to control the degree of methylation of specific glutamates in the MCPs. This resets the signalling state of the receptors and allows them to adapt to the present concentration of attractant and to sense subsequent changes. The dephosphorylation of phosphorylated CheY is accelerated by the phosphatase CheZ. P, phosphoryl grou.")
Presentazioni simili