Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
LEZIONE 7 Anno Accademico 2010/11
BIOTECNOLOGIE FARMACOLOGICHE CORSO DI LAUREA SPECIALISTICA IN BIOTECNOLOGIE DEL FARMACO LEZIONE 7 Anno Accademico 2010/11

2
LE BASI BIOLOGICHE DELL’INVECCHIAMENTO
e ambiente Invecchiamento e genetica Regolazione endocrina dell’invecchiamento

3
Non-Programmed Passive Aging Theories
Aging is a passive result of an organism’s inability to better resist fundamental deteriorative processes. Aging serves no purpose, is not an adaptation, is not programmed. Compatible with traditional evolutionary mechanics theory. Mammals needing more time for development needed a longer life span and therefore developed better maintenance and repair mechanisms that consequently delayed onset of age-related symptoms and diseases relative to shorter-lived mammals. Poor fit to many other observations of humans, other mammals, and other organisms particularly those that die suddenly from apparent biological suicide following reproduction rather than from gradual deterioration (e.g. Octopus, salmon)
")
4
Programmed Active Aging Theories
Organisms are purposely designed and genetically programmed to age or otherwise limit life span because the deterioration and life span limitation serves an evolutionary purpose. Aging is an adaptation, a purposeful design feature resulting from the evolution process. Aging is the result of a potentially complex active aging mechanism or “life span management system.” The mechanism could sense external conditions in order to adapt life span to local or temporary conditions and could operate by manipulating the maintenance and repair functions. Provides excellent fit to observations in humans, mammals, and other organisms. Incompatible with traditional “survival of the fittest” individual benefit requirement; requires an alternative mechanics theory. Supported and predicted by several alternative mechanics theories.

5
Planned Obsolescence Theory
Aging Theories Planned Obsolescence Theory Telomerase Theory of Aging The Neuroendocrine Theory The Free Radical Theory Mitochondrial Theory of Aging The Membrane Theory of Aging The Hayflick Limit Theory (The cell waste accumulation) Glycosylation Theory of Aging Immune system alterations
 Glycosylation Theory of Aging. Immune system alterations.")
6
Aging Theory Status “Main line” consensus of current gerontologists favors the passive theories. Earlier simple deterioration theories have little current scientific credibility in the biology community while still popular in the human-oriented (physician) community. Some relatively recent discoveries appear to favor aging-by-design theories. Efforts to explain aging based on traditional mechanics and efforts to explain other discrepancies with alternative mechanics cannot be simultaneously valid. Eventually there will be a unified theory.
 community. Some relatively recent discoveries appear to favor aging-by-design theories. Efforts to explain aging based on traditional mechanics and efforts to explain other discrepancies with alternative mechanics cannot be simultaneously valid. Eventually there will be a unified theory.")
7
Rougheye Rockfish 205 Years
“Non-Aging” Species Some species have been identified that apparently do not age or have negligible senescence. Older individuals do not appear to be weaker, less agile, less reproductive, more susceptible to disease, or otherwise less fit than younger animals. (Ages of some wild animals can be determined by annual marks in scales or bones similar to tree rings.) Some species with age of oldest recorded specimen: Rougheye Rockfish 205 Years Lake Sturgeon152 Years Aldabra Tortise152 Years Common U.S. Eastern Box Turtle is also long-lived (~100 years). Non-aging species tend to defeat simple deterioration theories and suggest dramatically longer human life spans are possible.
 Some species with age of oldest recorded specimen: Rougheye Rockfish 205 Years. Lake Sturgeon152 Years. Aldabra Tortise152 Years. Common U.S. Eastern Box Turtle is also long-lived (~100 years). Non-aging species tend to defeat simple deterioration theories and suggest dramatically longer human life spans are possible.")
8
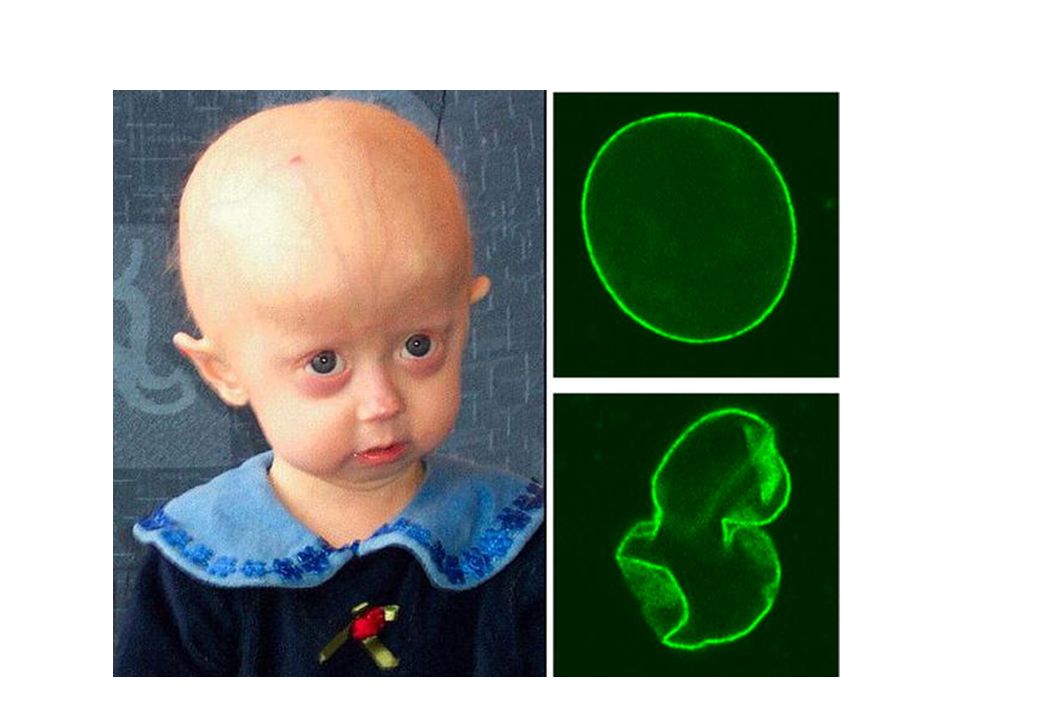
Hutchinson-Guilford Progeria and Werner syndrome
Hutchinson-Guilford Progeria, a very rare human genetic disease, accelerates many symptoms of aging including atherosclerotic heart disease. Victims usually die by age 13. Werner syndrome, another genetic disease, involves acceleration of most symptoms of aging including baldness, hair and skin conditions, heart disease, calcification of blood vessels, some cancers, cataracts, arthritis, diabetes, etc. Victims usually die by age 50. These conditions suggest aging is centrally controlled such that a single genetic defect could result in proportionally accelerating all of the expressed symptoms. Central control suggests aging-by-design

9
Hutchinson-Gilford progeria syndrome
Una malattia autosomica dominante e sporadica e rara che determina invecchiamento precoce: in genere il paziente muore a 13 anni circa per patologie cardiache La base genetica per molti casi di questa patologia consiste nella mutazione della tripletta GGC in GGT nel codone 608 della laminina A (LMNA) . Questo determina l’insorgenza di un sito di splicing criptico porta alla sintesi di una proteina con una delezione di 50 aa. La regione deleta ha in se la sequenza riconosciuta da enzimi proteolitici che fanno maturare la Laminina. In mancanza di questa parte della proteina, questa viene carbossifarnesilata e si accumula a livello endocellulare e soprattutto a causa della farnesilazione, nella membrana nucleare.
 . Questo determina l’insorgenza di un sito di splicing criptico porta alla sintesi di una proteina con una delezione di 50 aa. La regione deleta ha in se la sequenza riconosciuta da enzimi proteolitici che fanno maturare la Laminina. In mancanza di questa parte della proteina, questa viene carbossifarnesilata e si accumula a livello endocellulare e soprattutto a causa della farnesilazione, nella membrana nucleare.")
10
Invecchiamento e genetica
Figure 1. Processing of lamin A in normal and HGPS cells Meshorer E., Gruenbaum Y. J. Cell Biol. 2008:181:9-13

11
La presenza di laminina mutata (progerin)altera le funzione della membrana nucleare, la sua permeabilità e la trascrizione genica.
altera le funzione della membrana nucleare, la sua permeabilità e la trascrizione genica.")
13
La laminina A è una proteina della membrana nucleare che si posizione nella porzione intranucleare e partecipa alla organizzazione dei processi che presiedono la biosintesi di RNA e DNA. La Prelaminina A contiene un CAAX box nella sua porzione carbossiterminale che ne permette la farnesilazione ed il suo legame con la membrana nucleare; l’intervento di una metalloproteasi specifica taglia il frammento farnesilato producendo la Laminina A che ha una legame meno forte con la membrana nucleare e puo’ svolgere la propria attività intranucleare.

14
Capell and Collins, Nature Reviews Genetics 2006.

15
The is a mouse model of progeria where the prelamin A is not mutated
The is a mouse model of progeria where the prelamin A is not mutated. Instead, the metallopeptidase ZMPSTE24, the specific protease that is required to remove the C-terminus of prelamin A, is missing.

18
Una patologia autosomica recessiva
Sindrome di Werner Una patologia autosomica recessiva La mutazione genica è a carico della DNA elicasi (cromosoma 8 braccio corto) che accorcia la lunghezza dei telomeri. La malattia si manifesta alla pubertà e i portatori della mutazione vivono fino circa 40 anni di età.
 che accorcia la lunghezza dei telomeri. La malattia si manifesta alla pubertà e i portatori della mutazione vivono fino circa 40 anni di età.")
19
A yeast protein similar to the human WRN protein, called SGS1, has been found.
Mutations in SGS1 cause yeast to have a shorter lifespan than yeast cells without the mutation, and shown other signs typical of aging in yeast, such as an enlarged and fragmented nucleolus. Using yeast as a model for human aging in general, may give insight into the mechanisms of Werner syndrome and related diseases

20
When replication forks stall, the stable maintenance of replisome components requires the ATR kinase Mec1/Ddc2 and the RecQ helicase Sgs1.

21
A. Topo I usually found in eukaryotes binds the 3’ end of the broken DNA strand, and removes (+) or (-) supercoils. As replicating DNA moves through the structure, the two parental strands (black) are separated by the helicase, while positive supercoiling is removed by the 3’ topoisomerase. B. A machine able to separate the daughter molecules at the end of replication is formed by a helicase (red) removing the last turns of parental DNA and a type II topoisomerase (green) untangling the daughter duplexes.
 removing the last turns of parental DNA and a type II topoisomerase. (green) untangling the daughter duplexes.")
22
C. Nucleosome disruption.
The positive supercoiling produced by the translocating helicase H (red) destabilizes the nucleosome, while a topoisomerase T (5’ or 3’ Topo I, or eukaryotic topo II, green) efficiently relaxes the negative supercoiling, reforming the normal duplex behind the helicase.
 destabilizes the nucleosome, while a topoisomerase T (5’ or 3’ Topo I, or eukaryotic topo II, green) efficiently relaxes the negative supercoiling, reforming the normal duplex behind the helicase.")
23
STUDIARE VERMI E INSETTI PER CAPIRE L’UOMO
Drosophila melanogaster STUDIARE VERMI E INSETTI PER CAPIRE L’UOMO Coenorabditis elegans

24
CICLO VITALE DI C.ELEGANS
adulto L4 embrioni Circa 3 giorni a 22°C L3 L1 L2

25
CICLO VITALE DI C.ELEGANS
adulto L4 embrioni MANCANZA DI ALIMENTI STADIO DAUER L1

26
LARVA LARVA DAUER

28
Studio di processi biologici legati a una maggiore morbidità
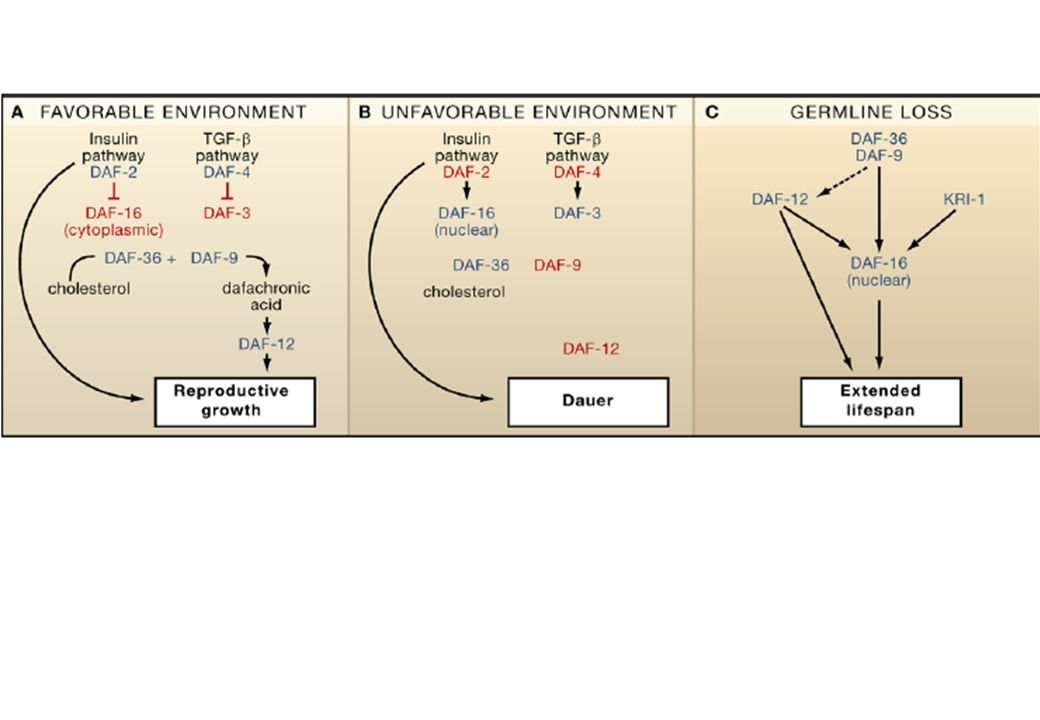
l’esempio dell’invecchiamento DAF1 (IGF-R) DAF 7 ( TGFb ligand) DAF 4 (Type II TGFbR) AGE 1 (IP3-K) DAF 3, DAF 5 (SMAD prot) DAF 12 DAF 16* 3-keto-cholestenoic acid metabolite DAUER SIR2 (deacetilasi attiva di DAF 16) DAF9 (cytochrome C CYP27A1) * Proteine della famiglia FOXO coinvolte nel metab del glucosio
 DAF 7 ( TGFb ligand) DAF 4 (Type II TGFbR) AGE 1 (IP3-K) DAF 3, DAF 5 (SMAD prot) DAF 12. DAF 16* 3-keto-cholestenoic acid metabolite. DAUER. SIR2 (deacetilasi attiva di DAF 16) DAF9 (cytochrome C. CYP27A1) * Proteine della famiglia FOXO coinvolte nel metab del glucosio.")
29
DAF 16/FOXO (adip. Tissue)
GH Insulin Insulin/IGF-1 Insulin/IGF-1 IGF-1 DAF 2 receptor IGF-1R 1R dFOXO TOR DAF 16/FOXO (adip. Tissue) TOR LONGEVITY LONGEVITY LONGEVITY germline germline
 TOR. LONGEVITY. LONGEVITY. LONGEVITY. germline. germline.")
30
Invecchiamento e ambiente

31
LAND OF BIOLOGICAL EQUAL OPPORTUNITIES
EVOLUTION: LAND OF BIOLOGICAL EQUAL OPPORTUNITIES SEXUAL REPRODUCTION “EFFECTOR” NUTRIMENT AGE “REGULATORS”

32
but AGE NUTRIENTS SEXUAL REPRODUCTION
EVOLUTION: LAND OF BIOLOGICAL EQUAL OPPORTUNITIES FECUNDITY SHOULD BE DIRECTLY PROPORTIONAL TO NUTRIENT AVAILABILITY, but DEATH: A TOOL INDISPENSABLE TO ENSURE THE CONTINUATION OF THE SPECIE HIGH NUTRIENT AVAILABILITY, FAVORING FECUNDITY, SHOULD SHORTEN THE LIFE SPAN AGE NUTRIENTS SEXUAL REPRODUCTION

33
AS NECESSITY FOR THE CONTINUATION OF LIFE
AGING AS NECESSITY FOR THE CONTINUATION OF LIFE and AS A MEAN TO GIVE TO EACH INDIVIDUAL EQUAL POSSIBILITIES TO GIVE HIS GENETIC CONTRIBUTION TO THE NEXT GENERATION Intrinsic program for aging aiming at increasing the fraility of the organism: a biological clock(telomers length, mitochondrial viability; DNA replication errors, loss of immune control and inflammation…) sex-dependent (male fecundity cannot be limited as well as in females) Fertility-driven Extrinsic factors nutrition adaptable environment
 sex-dependent (male fecundity cannot be limited as well as in females) Fertility-driven. Extrinsic factors. nutrition. adaptable environment.")
Presentazioni simili



 064825120 - fax.>")













 abbiamo cominciato a lavorare utilizzando i maniera didattica tecnologie di tipo hardware.>")



