Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
FOSFORILAZIONE OSSIDATIVA
Tutte le vie metaboliche della degradazione ossidativa dei carboidrati, degli acidi grassi e degli amminoacidi convergono in una tappa finale, in cui l’energia prodotta dalle ossidazioni viene utilizzata per la sintesi di ATP. La fosforilazione ossidativa è la sintesi di ATP che si verifica in seguito al trasferimento degli elettroni, sottratti durante le ossidazioni, all’ossigeno. La fosforilazione ossidativa nelle cellule eucariotiche ha luogo nei mitocondri; nelle cellule procariotiche sulla membrana citoplasmatica.

2
I Mitocondri I mitocondri sono organelli cellulari provvisti di doppia membrana. La membrana esterna è permeabile a piccole molecole e ioni. La membrana interna è permeabile solo a CO2, H2O e O2, essendo IMPERMEABILE alla maggior parte delle piccole molecole e degli ioni, compresi i protoni (H+). Tuttavia, attraverso la membrana interna è possibile il passaggio di specie chimiche aventi specifici trasportatori di membrana. Sulla membrana mitocondriale interna sono localizzati i componenti della catena di trasporto degli elettroni e quelli deputati alla sintesi di ATP.
. Tuttavia, attraverso la membrana interna è possibile il passaggio di specie chimiche aventi specifici trasportatori di membrana. Sulla membrana mitocondriale interna sono localizzati i componenti della catena di trasporto degli elettroni e quelli deputati alla sintesi di ATP.")
3
I Mitocondri Lo spazio fra le due membrane è detto spazio intermembrana. Nella matrice mitocondriale hanno luogo le reazioni di: decarbossilazione ossidativa del piruvato; ciclo acido citrico; ossidazione degli acidi grassi; ossidazione degli amminoacidi.

4
Catena di trasporto degli elettroni
Le reazioni cataboliche determinano la sottrazione di elettroni dalle molecole di substrati ridotti. Questi elettroni vengono transitoriamente convogliati sui coenzimi specifici delle varie deidrogenasi: NAD+ , FAD e NADP+ La catena di trasporto degli elettroni è costituita da una serie di trasportatori di elettroni, la gran parte dei quali sono proteine integrali di membrana, contenenti gruppi prostetici capaci di accettare o donare elettroni. e- trasferiti a specifici trasportatori fino all’ossigeno (accettore finale) e- utilizzati durante reazioni anaboliche (biosintesi riduttive) citocromi proteine Fe-S ubichinone
 e- utilizzati durante reazioni anaboliche (biosintesi riduttive) citocromi. proteine Fe-S. ubichinone.")
5
Citocromi I citocromi sono proteine contenenti un gruppo eme (Fe2+ Fe3+) Vi sono 3 classi di citocromi: a e b: il gruppo eme è legato alla proteina non covalentemente; sono proteine integrali della membrana mitocondriale interna; nei citocromi b è presente lo stesso gruppo eme della mioglobina e della emoglobina c: il gruppo eme è legato covalentemente a residui di cisteina (Cys) della proteina; alcuni citocromi di tipo c sono proteine integrali della membrana mitocondriale interna; eccezione: il citocromo c è una proteina solubile che si lega alla superficie esterna della membrana mitocondriale interna. I citocromi trasportano elettroni.
 della proteina; alcuni citocromi di tipo c sono proteine integrali della membrana mitocondriale interna; eccezione: il citocromo c è una proteina solubile che si lega alla superficie esterna della membrana mitocondriale interna. I citocromi trasportano elettroni.")
6
Gruppo EME dei citocromi

7
Le proteine ferro-zolfo
Le proteine ferro-zolfo contengono ferro associato ad atomi di zolfo [FeS]. Gli atomi di zolfo possono essere residui di cisteina o zolfo inorganico. Le proteine ferro-zolfo possono avere strutture semplici (un solo atomo di ferro) o complesse (da due a quattro atomi di ferro). Nelle proteine ferro-zolfo di Rieske, un tipo particolare di proteina ferro-zolfo, l’atomo di ferro è coordinato con 2 residui di istidina invece che con 2 residui di cisteina. Nelle proteine ferro-zolfo l’atomo di ferro può assumere stati di ossidazione Fe2+ o Fe3+. Le proteine Fe-S trasportano elettroni.
 o complesse (da due a quattro atomi di ferro). Nelle proteine ferro-zolfo di Rieske, un tipo particolare di proteina ferro-zolfo, l’atomo di ferro è coordinato con 2 residui di istidina invece che con 2 residui di cisteina. Nelle proteine ferro-zolfo l’atomo di ferro può assumere stati di ossidazione Fe2+ o Fe3+. Le proteine Fe-S. trasportano elettroni.")
8
Ubichinone (coenzima Q)
L’ubichinone (coenzima Q) è un benzochinone con lunga catena isoprenoide. Il coenzima Q può accettare 1 elettrone e 1 protone (trasformandosi in radicale semichinonico) oppure 2 elettroni e 2 protoni (riducendosi completamente a ubichinolo QH2) Il coenzima Q è di piccole dimensioni e di natura idrofobica, pertanto può diffondere liberamente nel doppio strato lipidico della membrana mitocondriale interna fungendo da ponte tra i trasportatori di elettroni. Il coenzima Q trasporta elettroni e protoni.
 è un benzochinone con lunga catena isoprenoide. Il coenzima Q può accettare 1 elettrone e 1 protone (trasformandosi in radicale semichinonico) oppure 2 elettroni e 2 protoni (riducendosi completamente a ubichinolo QH2) Il coenzima Q è di piccole dimensioni e di natura idrofobica, pertanto può diffondere liberamente nel doppio strato lipidico della membrana mitocondriale interna fungendo da ponte tra i trasportatori di elettroni. Il coenzima Q trasporta elettroni e protoni.")
9
Catena di trasporto degli elettroni
I trasportatori di elettroni sono organizzati in quattro complessi multienzimatici posizionati in maniera ordinata sulla membrana mitocondriale interna . L’affinità per gli elettroni aumenta dal complesso I fino all’ossigeno. Ogni complesso è caratterizzato da una propria composizione in proteine e dalla presenza di determinati gruppi prostetici.

10
Catena di trasporto degli elettroni

11
Complesso I Il complesso I (NADH-coenzima Q ossido reduttasi) è costituito da diverse proteine Fe-S e da una flavo proteina contenente FMN. Questo complesso riceve gli elettroni dal NADH, prodotto durante le reazioni cataboliche, e li cede al coenzima Q. Al coenzima Q arrivano anche i 2 protoni (del NADH e H+). Il coenzima Q ridotto (QH2) diffonde nella membrana mitocondriale interna verso il complesso III. Il trasferimento di elettroni porta contemporaneamente alla fuoriuscita dalla matrice verso lo spazio intermembrana di protoni (H+). Il percorso degli elettroni è: NADH → flavoproteina → proteine Fe-S → coenzima Q.
 è costituito da diverse proteine Fe-S e da una flavo proteina contenente FMN. Questo complesso riceve gli elettroni dal NADH, prodotto durante le reazioni cataboliche, e li cede al coenzima Q. Al coenzima Q arrivano anche i 2 protoni (del NADH e H+). Il coenzima Q ridotto (QH2) diffonde nella membrana mitocondriale interna verso il complesso III. Il trasferimento di elettroni porta contemporaneamente alla fuoriuscita dalla matrice verso lo spazio intermembrana di protoni (H+). Il percorso degli elettroni è: NADH → flavoproteina → proteine Fe-S → coenzima Q.")
12
Complesso II Il complesso II (succinato-coenzima Q ossido reduttasi) è di piccole dimensioni e non attraversa completamente la mb mitocondriale interna. È costituto da: - SUCCINATO DEIDROGENASI (ciclo acido citrico ,ox del succinato a fumarato), coenzima FAD, - proteine FeS - citocromo di tipo b. Il coenzima Q riceve e- da: Complesso I e II - Acil CoA deidrogenasi-FAD dipendente (β-ox acidi grassi) - glicerolo-3P deidrogenasi (sistema navetta per traslocazione NADH da citosol a matrice mitocondriale) Il percorso degli elettroni è: Succinato → FAD → proteine Fe-S → coenzima Q
 è di piccole dimensioni e non attraversa completamente la mb mitocondriale interna. È costituto da: - SUCCINATO DEIDROGENASI (ciclo acido citrico ,ox del succinato a fumarato), coenzima FAD, - proteine FeS. - citocromo di tipo b. Il coenzima Q riceve e- da: Complesso I e II. - Acil CoA deidrogenasi-FAD dipendente (β-ox acidi grassi) - glicerolo-3P deidrogenasi. (sistema navetta per traslocazione NADH da citosol a matrice mitocondriale) Il percorso degli elettroni è: Succinato → FAD → proteine Fe-S → coenzima Q.")
13
Complesso III Il complesso III, detto Coenzima Q citocromo c ossido reduttasi, è costituto da proteine FeS, citocromo b (cit bH, cit bL), citocromo c1. Il complesso III riceve gli elettroni dal coenzima Q e li trasferisce al citocromo c. Il citocromo c non fa parte del complesso III ma si muove lungo la membrana mitocondriale interna verso il complesso IV. Il complesso III è un altro sito dove i protoni fuoriescono dalla matrice. Il percorso degli elettroni è: coenzima Q → proteine FeS → cit c1 → cit c
, citocromo c1. Il complesso III riceve gli elettroni dal coenzima Q e li trasferisce al citocromo c. Il citocromo c non fa parte del complesso III ma si muove lungo la membrana mitocondriale interna verso il complesso IV. Il complesso III è un altro sito dove i protoni fuoriescono dalla matrice. Il percorso degli elettroni è: coenzima Q → proteine FeS → cit c1 → cit c.")
14
Complesso IV Il complesso IV (citocromo c ossidasi) è costituto da proteine rame-zolfo (CuA, CuB), citocromo a, citocromo a3. Il complesso IV riceve gli elettroni dal citocromo c e li trasferisce all’ossigeno che si riduce ad H2O. Al trasferimento degli elettroni si associa la fuoriuscita di protoni (H+) dalla matrice verso lo spazio intermembrana. Il percorso degli elettroni è: Cit c → CuA → cit a → cit a3 → CuB → O2
 dalla matrice verso lo spazio intermembrana. Il percorso degli elettroni è: Cit c → CuA → cit a → cit a3 → CuB → O2.")
15
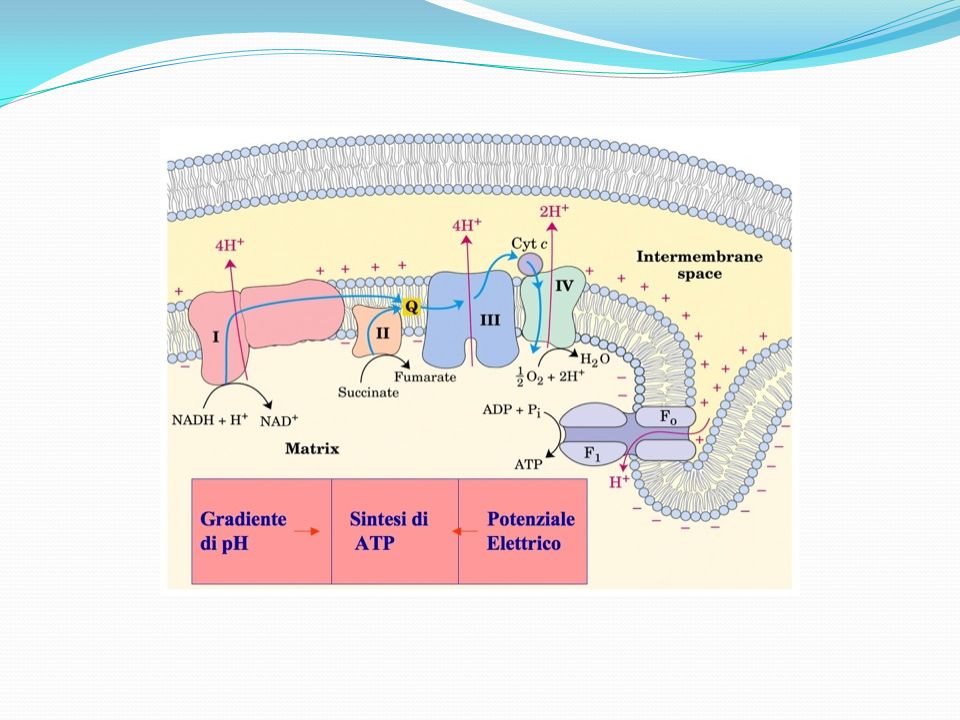
Flusso Di Protoni Per ogni coppia di elettroni trasferita all’ossigeno fuoriescono protoni (H+) dalla matrice verso lo spazio intermembrana. Poiché la membrana mitocondriale interna è impermeabile ai protoni, questi fuoriescono attraverso i complessi multiproteici I, III e IV. Vengono trasferiti: 4 protoni dal complesso I 4 dal complesso III 2 dal complesso IV In totale, per ogni coppia di elettroni fuoriescono 10 protoni. Si genera in questo modo un GRADIENTE DI CONCENTRAZIONE PROTONICA tra i due lati della membrana mitocondriale interna, con una concentrazione di protoni più alta nello spazio intermembrana che nella matrice.
 dalla matrice verso lo spazio intermembrana. Poiché la membrana mitocondriale interna è impermeabile ai protoni, questi fuoriescono attraverso i complessi multiproteici I, III e IV. Vengono trasferiti: 4 protoni dal complesso I. 4 dal complesso III. 2 dal complesso IV In totale, per ogni coppia di elettroni fuoriescono 10 protoni. Si genera in questo modo un GRADIENTE DI CONCENTRAZIONE PROTONICA tra i due lati della membrana mitocondriale interna, con una concentrazione di protoni più alta nello spazio intermembrana che nella matrice.")
17
Fosforilazione ossidativa
La membrana mitocondriale interna è impermeabile ai protoni e questi rientrano nella matrice attraverso i canali ionici dell’ATP sintasi. L’ATP sintasi è una proteina di membrana costituita da 2 componenti, F1 e Fo. Fo: attraversa la membrana e consiste di 3 subunità diverse (a, b, c), funge da canale proteico per il passaggio di H+. F1: sporge nella matrice e consiste di 5 subunità diverse, così organizzate α3β3γδε, contiene il sito catalitico per la sintesi di ATP. Quando i protoni rientrano nella matrice, il flusso di H+ determina un cambio conformazionale dell’ATP-sintasi che rende l’enzima attivo, capace di fosforilare l’ADP ad ATP. Quindi, l’energia elettrochimica del gradiente protonico fornisce l’energia per la sintesi di ATP da ADP e Pi. Teoria chemiosmotica che associa il flusso protonico alla sintesi di ATP (fosforilazione ossidativa).
, funge da canale proteico per il passaggio di H+. F1: sporge nella matrice e consiste di 5 subunità diverse, così organizzate α3β3γδε, contiene il sito catalitico per la sintesi di ATP. Quando i protoni rientrano nella matrice, il flusso di H+ determina un cambio conformazionale dell’ATP-sintasi che rende l’enzima attivo, capace di fosforilare l’ADP ad ATP. Quindi, l’energia elettrochimica del gradiente protonico fornisce l’energia per la sintesi di ATP da ADP e Pi. Teoria chemiosmotica che associa il flusso protonico alla sintesi di ATP (fosforilazione ossidativa).")
18
Cambi conformazionali nella molecola di ATP sintasi
Le modifiche della conformazione nell’ATP sintasi sono essenziali per la sintesi di ATP. Nella componente F1 ciascuna delle subunità β possiede un sito catalitico per la sintesi di ATP. Le subunità β possono assumere tre diverse conformazioni, interconvertibili tra loro: Nella conformazione β-ADP la subunità β lega ADP e Pi. Nella conformazione β-ATP la subunità β catalizza la sintesi di ATP. Nella conformazione β-vuota la subunità β rilascia l’ATP. In un determinato tempo ogni subunità è in uno dei tre diversi stati conformazionali, di conseguenza le tre subunità β catalizzano a turno la sintesi di ATP.

19
Regolazione della fosforilazione ossidativa
La fosforilazione ossidativa è regolata in modo tale da produrre un numero di molecole di ATP adeguato alle esigenze della cellula. Il principale regolare è la carica energetica cellulare. La concentrazione di ADP e il rapporto [ATP]/[ADP]+[P] sono misure dello stato energetico della cellula. Se la concentrazione di ATP diminuisce → la velocità del trasporto degli elettroni e della fosforilazione ossidativa aumentano. Se la concentrazione di ATP aumenta → la velocità del trasporto degli elettroni e della fosforilazione ossidativa diminuiscono.

20
ATP prodotto Il numero di molecole di ATP prodotte dipende dai protoni che fuoriescono dalla matrice verso lo spazio intermembrana. I protoni che fuoriescono sono: 4 dal complesso I 4 dal complesso III Totale 10 H+ per ogni coppia di e- 2 dal complesso IV da NADH 4 dal complesso III Totale 6 H+ per ogni coppia di e- 2 dal complesso IV da FADH2 4 protoni che rientrano attraverso l’ATP-sintasi producono l’energia per la sintesi di 1 molecola di ATP. Se il donatore di elettroni è il NADH si producono 2.5 molecole di ATP. Se il donatore di elettroni è il FADH2 si producono 1.5 molecole di ATP.

21
Resa totale di ATP

22
Radicali liberi e stress ossidativo
Durante il normale trasporto degli elettroni l’ossigeno: - può essere ridotto parzialmente; - può accettare singoli elettroni. In tal caso si possono formare derivati instabili, noti come specie reattive dell’ossigeno (ROS), contenenti uno o più elettroni spaiati. I principali radicali liberi prodotti sono: Gli antiossidanti proteggono la cellula dai radicali liberi agendo come “scavenger”. Se in eccesso i radicali liberi possono causare danni alle strutture cellulari: lipidi di membrana; proteine; acidi nucleici. radicale superossido O2- perossido di idrogeno H2O2 ossigeno singoletto 1O2 radicale ossidrile •OH
, contenenti uno o più elettroni spaiati. I principali radicali liberi prodotti sono: Gli antiossidanti proteggono la cellula dai radicali liberi agendo come scavenger . Se in eccesso i radicali liberi possono causare danni alle strutture cellulari: lipidi di membrana; proteine; acidi nucleici. radicale superossido O2- perossido di idrogeno H2O2. ossigeno singoletto 1O2. radicale ossidrile •OH.")
23
Schema del trasferimento di elettroni e protoni attraverso la membrana interna dei mitocondri

24
Schema riassuntivo del metabolismo ossidativo

Presentazioni simili