Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

2
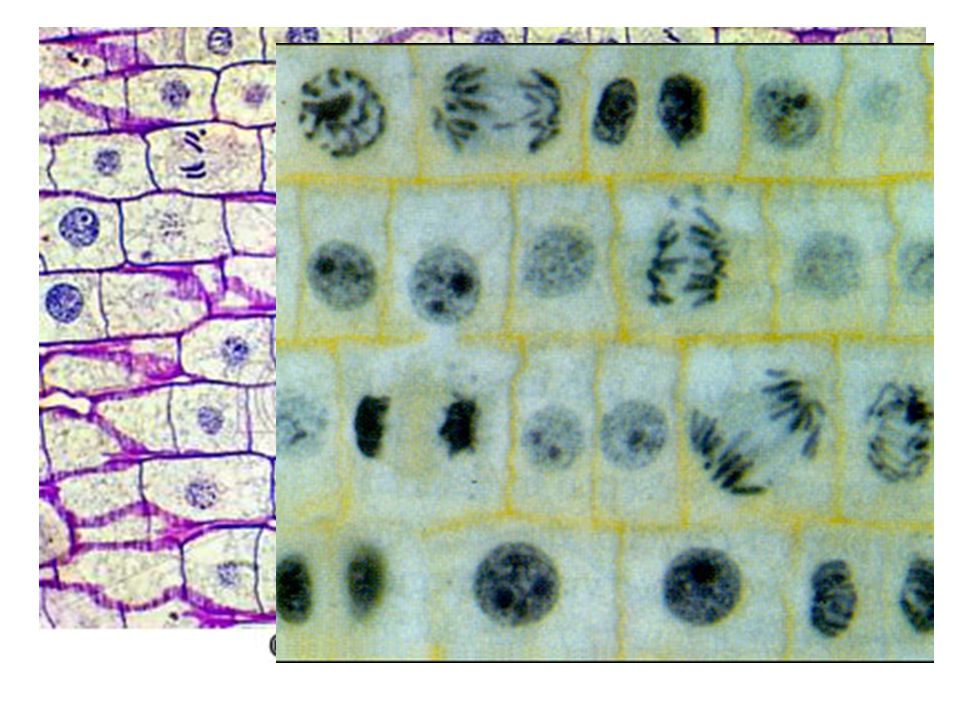
Ogni cellula deriva per divisione da una cellula preesistente: “omnis cellula e cellula” procarioti ed eucarioti hanno divisioni diverse. Negli eucarioti la divisione è più complicata Prima della divisione cellulare il DNA si duplica e, una volta duplicatosi il DNA, la cellula deve assolutamente dividersi. Tutti gli organismi pluricellulari si originano da una cellula uovo fecondata, attraverso una serie di divisioni cellulari (mitosi). Successivamente le cellule si specializzano per svolgere ruoli diversi. Mitosi: - interessa le cellule somatiche, ogni nuova cellula è uguale alla cellula madre. Riproduzione o divisione della cellula. Significato diverso e processi diversi si hanno con la riproduzione degli organismi (sessuata o asessuata). Nel primo caso interviene un tipo di divisione cellulare particolare: la Meiosi. Questa interessa le cell. germinali, vengono prodotte 4 cellule figlie: ogni cellula figlia ha metà dei cromosomi della cellula madre. Sono coinvolti 2 processi di divisione cellulare, riduzionale ed equazionale. Rudolf Virchow nel 1858 Il processo o l’insieme dei processi mediante i quali gli esseri viventi perpetuano la propria specie, producendo nuovi individui che ripetono ciclicamente le medesime fasi di sviluppo fino al raggiungimento della forma definitiva propria di quelli da cui sono derivati. La r. può essere asessuata o sessuata. La r. asessuata è prevalentemente (ma non esclusivamente) adottata da organismi unicellulari (Batteri ecc.) ma anche da alcuni organismi pluricellulari, come per es. l’idra. Essa avviene con modalità simili per cellule aploidi. In ogni caso, dà origine a una progenie di individui geneticamente identici (a meno di errori o mutazioni) tra loro e all’organismo parentale.
. Successivamente le cellule si specializzano per svolgere ruoli diversi. Mitosi: - interessa le cellule somatiche, ogni nuova cellula è uguale alla cellula madre. Riproduzione o divisione della cellula. Significato diverso e processi diversi si hanno con la riproduzione degli organismi (sessuata o asessuata). Nel primo caso interviene un tipo di divisione cellulare particolare: la Meiosi. Questa interessa le cell. germinali, vengono prodotte 4 cellule figlie: ogni cellula figlia ha metà dei cromosomi della cellula madre. Sono coinvolti 2 processi di divisione cellulare, riduzionale ed equazionale. Rudolf Virchow nel Il processo o l’insieme dei processi mediante i quali gli esseri viventi perpetuano la propria specie, producendo nuovi individui che ripetono ciclicamente le medesime fasi di sviluppo fino al raggiungimento della forma definitiva propria di quelli da cui sono derivati. La r. può essere asessuata o sessuata. La r. asessuata è prevalentemente (ma non esclusivamente) adottata da organismi unicellulari (Batteri ecc.) ma anche da alcuni organismi pluricellulari, come per es. l’idra. Essa avviene con modalità simili per cellule aploidi. In ogni caso, dà origine a una progenie di individui geneticamente identici (a meno di errori o mutazioni) tra loro e all’organismo parentale.")
4
La riproduzione agamica o asessuata non coinvolge due individui e non comporta lo scambio o la messa in comune di materiale genetico. La si riscontra certamente nei procarioti, ma anche in alcuni eucarioti specie nelle piante (talee). Esistono possibilità di trasferimenti orizzontali di DNA. La riproduzione sessuata è più evoluta e offre maggiori possibilità per avere individui più adatti ad un particolare ambiente. Si può pertanto affermare che quest’ultima ha creato le condizioni per una più rapida evoluzione in piante e animali superiori. Nel mondo vegetale la conservazione della specie è assicurata in molti modi che riflettono la più lunga storia naturale delle piante, e la loro maggiore flessibilità adattativa rispetto agli animali, in relazione sia alla minore differenziazione sia al regime più vario e rigoroso di rapporti con l’ambiente.

5
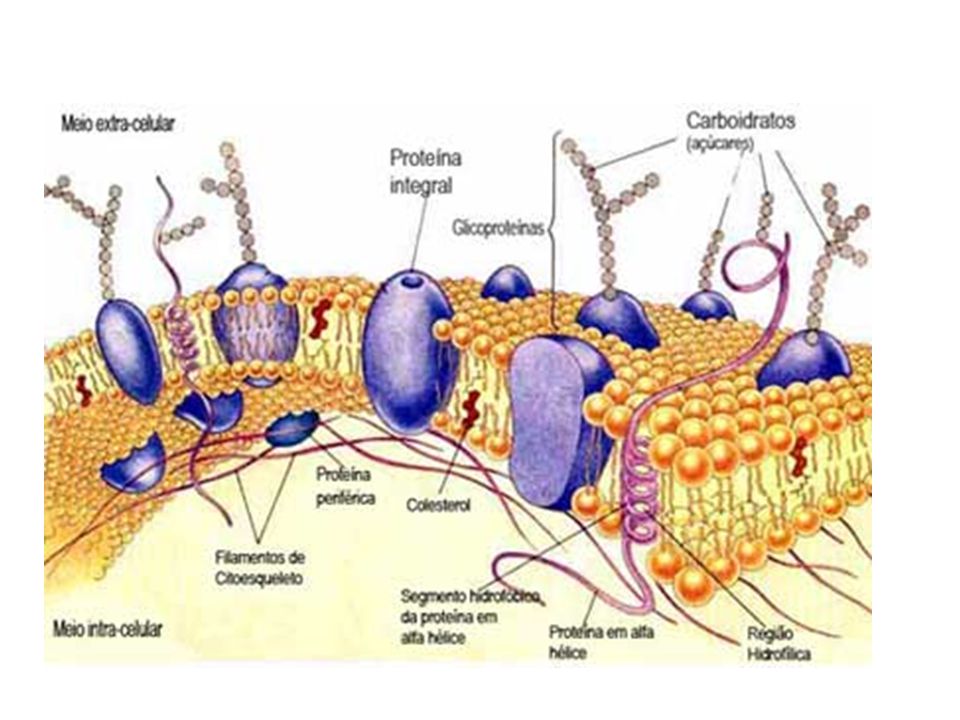
Membrana plasmatica, sistema di endomembrane, perossisomi, mitocondri, cloroplasti, ribosomi, citoscheletro

6
Tutte le cellule sono delimitate da una membrana detta membrana plasmatica

8
- La parete cellulare, esterna alla membrana, conferisce alla cellula una forma prismatica o poliedrica. - I vacuoli sono piccoli e numerosi nella cellula giovane, nella cellula matura si ingrandiscono e si fondono in un grande vacuolo che arriva ad occupare anche il 90% del volume cellulare. - I plastidi: cloroplasti (responsabili del colore verde delle foglie e coinvolti nella fotosintesi clorofilliana), cromoplasti (responsabile dei pigmenti dei fiori), leucoplasti (amiloplasti, lipoplasti etc. presenti nelle parti più interne della pianta e quindi meno esposte alla luce, come le radici).
, cromoplasti (responsabile dei pigmenti dei fiori), leucoplasti (amiloplasti, lipoplasti etc. presenti nelle parti più interne della pianta e quindi meno esposte alla luce, come le radici).")
9
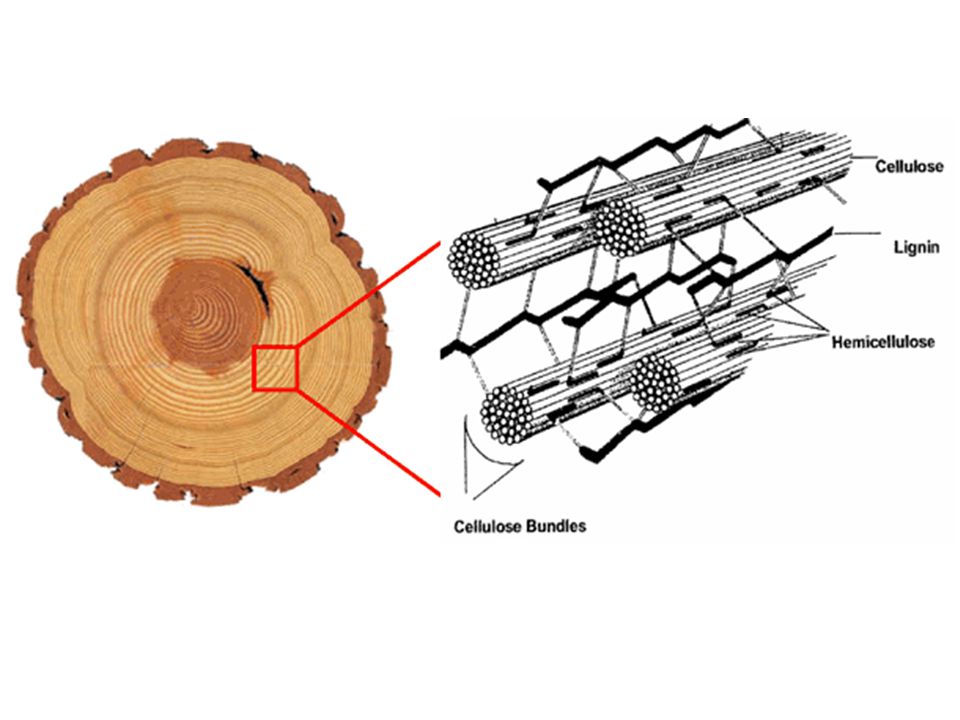
PARETE CELLULARE Costituisce il rivestimento esterno della cellula e rappresenta una sorta di involucro rigido. •Componente amorfa matrice • Emicellulose • Pectine • Proteine • Fenoli • Lignina •Componente fibrillare • Cellulosa

10
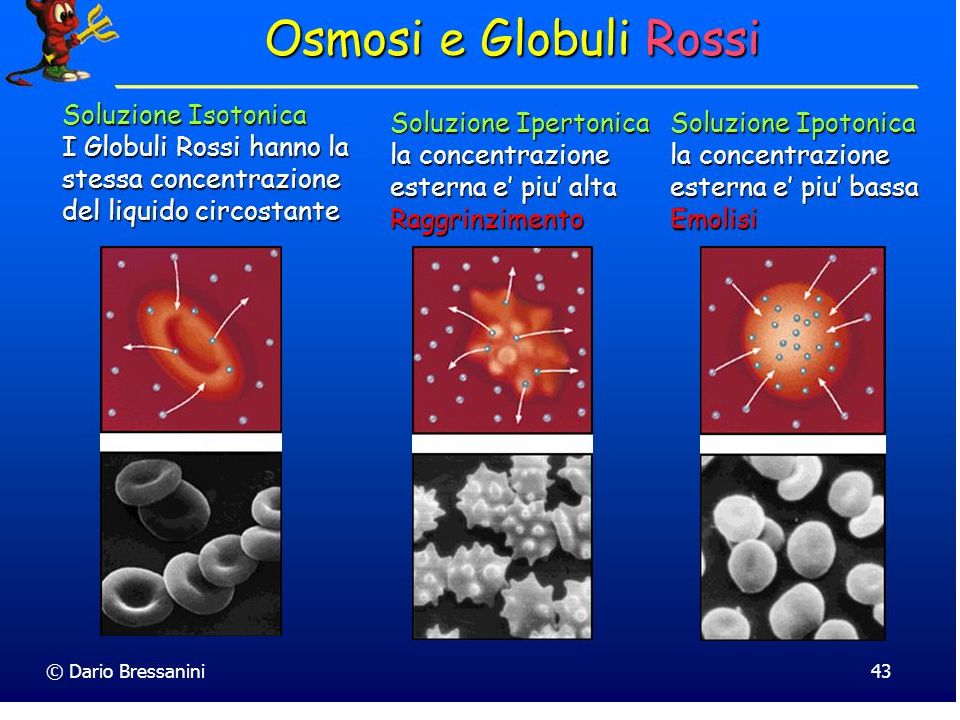
Si distingue in: Una sottile parete primaria Una più spessa parete secondaria; si trova in cellule differenziate come nel tessuto di conduzione o di sostegno. E’ situata all’interno della parete primaria. La porzione più esterna della parete, e quindi della cellula, è la lamella mediana, a contatto con la lamella mediana di altre cellule. E’ ricca in pectina e responsabile dell’adesione cellula-cellula. La parete previene lo scoppio di cellule poste in una soluzione ipotonica e determina la genesi della pressione di turgore data dalla pressione esercitata dal contenuto della cellule contro la parete cellulare stessa. Le cellule vegetali hanno un elevata conccentrazione di soluti nel citoplasma e quindi attirano acqua all’interno della cellula determinando rigonfiamento del vacuolo centrale che premerà contro la parete, impedendo una ulteriore entrata di acqua.

12
La parete primaria circonda le cellule che devono crescere e che si dividono (meristemi); fornisce resistenza meccanica ma permette alla cellula di espandersi per accrescersi e dividersi. E’ composta da pectina, emicellulosa, cellulosa (poca) e proteine. La parete primaria è il maggior componente degli alimenti vegetali. La maturazione dei frutti e delle verdure è associata a modificazioni che intervengono nella composizione e struttura della parete. I polisaccaridi della parete sono usati come addensanti e gelificanti Le pareti cellulari sono state sono paragonate a cemento armato; in esso sono immersi dei tondini di acciaio, (le microfibrille di cellulosa), che servono a rinforzare il cemento (la matrice). Disposizione fibrille di cellulosa!!!!!!!
, che servono a rinforzare il cemento (la matrice). Disposizione fibrille di cellulosa!!!!!!!")
13
Durante l’espansione cellulare la direzione predominante di crescita è influenzata dalla organizzazione della parete, tendendo ad essere perpendicolare alla direzione prevalente delle microfibrille. La disposizione delle microfibrille è determinata dal citoscheletro (microtubuli)
.")
14
La parete secondaria, più spessa e resistente, è deposta solo quando la cellula ha smesso di crescere. La si trova nelle cellule dello xilema, tracheidi e sclereidi. E’ composta di numerose fibre di cellulosa e piccole quantità di emicellulosa e pectine ed inoltre rinforzata da lignina, suberina, cere etc. La presenza di questa parete è stata di fondamentale importanza per la conquista della terra ferma facilitando il trasporto di acqua e di nutrienti e permettendo un crescita maggiore, protezione contro patogeni e perdita acqua. Tra le varie modificazioni cui essa può andare incontro sicuramente la più interessante è la lignificazione. La lignina è idrofoba e si sostituisce all’acqua nella parete cellulare

16
Per funzionare correttamente le cellule devono poter comunicare tra di loro, e la presenza di una parete, specie se secondaria, potrebbe impedire tale processo. Per ovviare al problema, la parete cellulare non è più una struttura continua ma presenta un numero più o meno elevato di interruzioni. Queste interruzioni sono dette plasmodesmi e mettono in comunicazione cellule vegetali vicine e lontane. Sono responsabili della comunicazione simplastica che mette ad esempio in comunicazione cellule della foglia con quelle della radice o viceversa.

17
Come si formano i plasmodesmi
Come si formano i plasmodesmi? Di solito in seguito ad una divisione cellulare (mitosi).
.")
19
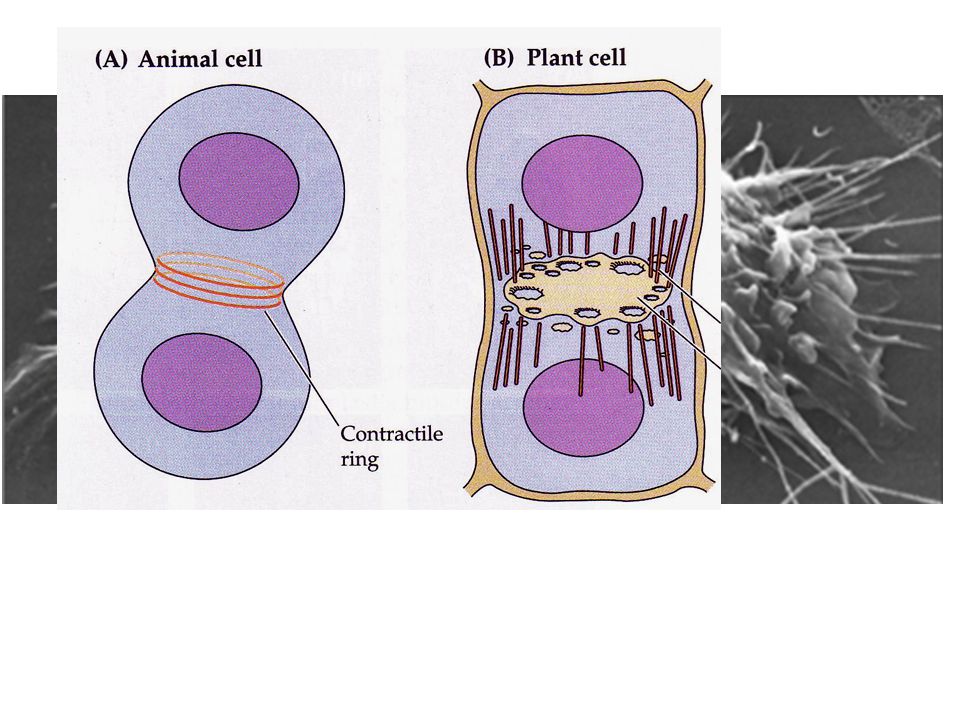
In telofase dai due apparati di Golgi si distaccano vescicole che guidate da microtubuli si portano nel piano equatoriale della cellula vescicole delimitate da membrana che formeranno la nuova membrana plasmatica e nel lume di queste viene secreta la parete primaria. Non tutte le vescicole si fondono e quindi i citoplasmi delle due cellule figlie rimangono in contatto. Fuso mitotico fragmoplasto Setto di divisione

20
Comunicazione simplastica (intracellulare) ed apoplastica (extracellulare tramite parete)
 ed apoplastica (extracellulare tramite parete)")
21
Plasmodesmata are now known to be structurally complex, with the capacity to dilate and facilitate the cell-to-cell transport of macromolecules, such as proteins, RNAs and protein–RNA complexes. Furthermore, they are also functionally dynamic channels, undergoing modifications in permeability, or changes in positional frequency along the cell wall, depending on the needs of the plant [12, 13, 14]. These characteristics allow plasmodesmata to establish domains of symplastically connected cells, so-called ‘symplastic domains,’ giving higher plants a means to produce different cell types, tissues and organs; they can also form cellular gateways to the vascular system, enabling plants to coordinate cellular responses at the whole-organism level [9, 15, 16]. The role of plasmodesmata in the symplastic pathway in plants. (a)A schematic diagram of a plasmodesma illustrating the ultrastructure and cell-to-cell trafficking of diffusible signaling molecules [28]. Orange-yellow spheres and short rods represent hypothetical proteinaceous and filamentous components observed within plasmodesma. (b) Plasmodesmal-mediated signaling among symplastically connected cells. Some signals (red arrows) move only into cells adjacent to the original cell that generated them for local communication, whereas systemic signals (black arrows) move farther to reach phloem for long-distance communication. (c)Environmental signals (e.g. day length or light intensity) or challenges (e.g. biotic stresses caused by microbial pathogen infection) perceived in the leaves are processed in the receptive cells (dark-green or yellow patch, respectively) and transported through plasmodesmata for local communication within a tissue [6]. These signals can then enter phloem (broken red arrows) for inter-organ signaling and are transported to distantly located target cells and tissues, such as the shoot or root tips, to bring about appropriate biochemical, physiological, and/or developmental changes. Broken blue arrows indicate xylem transport.
![Plasmodesmata are now known to be structurally complex, with the capacity to dilate and facilitate the cell-to-cell transport of macromolecules, such as proteins, RNAs and protein–RNA complexes. Furthermore, they are also functionally dynamic channels, undergoing modifications in permeability, or changes in positional frequency along the cell wall, depending on the needs of the plant [12, 13, 14]. These characteristics allow plasmodesmata to establish domains of symplastically connected cells, so-called ‘symplastic domains,’ giving higher plants a means to produce different cell types, tissues and organs; they can also form cellular gateways to the vascular system, enabling plants to coordinate cellular responses at the whole-organism level [9, 15, 16].](http://slideplayer.it/slide/2680820/10/images/21/Plasmodesmata+are+now+known+to+be+structurally+complex%2C+with+the+capacity+to+dilate+and+facilitate+the+cell-to-cell+transport+of+macromolecules%2C+such+as+proteins%2C+RNAs+and+protein%E2%80%93RNA+complexes.+Furthermore%2C+they+are+also+functionally+dynamic+channels%2C+undergoing+modifications+in+permeability%2C+or+changes+in+positional+frequency+along+the+cell+wall%2C+depending+on+the+needs+of+the+plant+%5B12%2C+13%2C+14%5D.+These+characteristics+allow+plasmodesmata+to+establish+domains+of+symplastically+connected+cells%2C+so-called+%E2%80%98symplastic+domains%2C%E2%80%99+giving+higher+plants+a+means+to+produce+different+cell+types%2C+tissues+and+organs%3B+they+can+also+form+cellular+gateways+to+the+vascular+system%2C+enabling+plants+to+coordinate+cellular+responses+at+the+whole-organism+level+%5B9%2C+15%2C+16%5D..jpg "The role of plasmodesmata in the symplastic pathway in plants. (a)A schematic diagram of a plasmodesma illustrating the ultrastructure and cell-to-cell trafficking of diffusible signaling molecules [28]. Orange-yellow spheres and short rods represent hypothetical proteinaceous and filamentous components observed within plasmodesma. (b) Plasmodesmal-mediated signaling among symplastically connected cells. Some signals (red arrows) move only into cells adjacent to the original cell that generated them for local communication, whereas systemic signals (black arrows) move farther to reach phloem for long-distance communication. (c)Environmental signals (e.g. day length or light intensity) or challenges (e.g. biotic stresses caused by microbial pathogen infection) perceived in the leaves are processed in the receptive cells (dark-green or yellow patch, respectively) and transported through plasmodesmata for local communication within a tissue [6]. These signals can then enter phloem (broken red arrows) for inter-organ signaling and are transported to distantly located target cells and tissues, such as the shoot or root tips, to bring about appropriate biochemical, physiological, and/or developmental changes. Broken blue arrows indicate xylem transport.")
22
AutoScreenRecorder_02 Oct. 13 10.46.avi

23
La parete cellulare svolge numerose funzioni.
Le principali sono: - Mantiene/determina la forma e la grandezza della cellula - Fornisce supporto e resistenza meccanica - Previene lo scoppio di cellule poste in una soluzione ipotonica e determina la genesi della pressione di turgore - Controlla la velocità e la direzione d’accrescimento - Ha una funzione di difesa; rappresenta una barriera fisica contro i patogeni - Riduce la perdita di H2O dai tessuti con opportune modificazioni secondarie (cuticola, cellule suberificate, ecc.) - Le proteine enzimatiche presenti nella parete permettono il rimodellamento della parete stessa e la formazione di molecole segnale
 - Le proteine enzimatiche presenti nella parete permettono il rimodellamento della parete stessa e la formazione di molecole segnale.")
24
VACUOLO È una cisterna tondeggiante rivestita da una membrana (tonoplasto ); a maturità un singolo vacuolo occupa la quasi totalità dello spazio intracellulare. La cellula immatura contiene numerosi piccoli vacuoli che, durante la fase di crescita per distensione, aumentano di dimensioni e si fondono in un singolo vacuolo. Le cellule usano i vacuoli per regolare la loro dimensione (senza modificare la quantità di citoplasma) e la pressione da turgore. I vacuoli sono compartimenti importanti per l’omeostasi cellulare, il turgore cellulare, i processi di detossificazione, le funzioni di riserva. Si ritengono simili ai lisosomi delle cellule animali, ma possono funzionare come deposito per sostanze di riserva o per metaboliti. Contengono acqua e varie sostanze come metaboliti vari, proteine, alcaloidi, pigmenti.
; a maturità un singolo vacuolo occupa la quasi totalità dello spazio intracellulare. La cellula immatura contiene numerosi piccoli vacuoli che, durante la fase di crescita per distensione, aumentano di dimensioni e si fondono in un singolo vacuolo. Le cellule usano i vacuoli per regolare la loro dimensione (senza modificare la quantità di citoplasma) e la pressione da turgore. I vacuoli sono compartimenti importanti per l’omeostasi cellulare, il turgore cellulare, i processi di detossificazione, le funzioni di riserva. Si ritengono simili ai lisosomi delle cellule animali, ma possono funzionare come deposito per sostanze di riserva o per metaboliti. Contengono acqua e varie sostanze come metaboliti vari, proteine, alcaloidi, pigmenti.")
25
Questi pigmenti presenti nelle foglie, nei fiori o nei frutti di diverse piante, possono funzionare da indicatori naturali di acidità perché sono in grado di cambiare colore al variare del pH. Essi assumono un colore completamente diverso in una soluzione acida rispetto ad una soluzione basica. L’estrazione dei pigmenti si ottiene con l’ebollizione in acqua. Le antocianine all’aumentare del pH cambiano colore passando dal rosso al blu. Il bicchiere con l’acqua di bollitura del cavolo rosso, ad un pH tra 6 e 7, dovrebbe avere un colore dal rosa al porpora. Possiamo aumentare ulteriormente il pH aggiungendo una sostanza alcalina. Sciogliendo un cucchiaino di bicarbonato di sodio nella soluzione il colore dovrebbe virare verso il blu, raggiungendo pH 8.. Si potrà misurare così il pH del succo di limone, dell’aceto, del bicarbonato, della birra, dell’ aspirina ecc. Scala

26
Plastidi -I cloroplasti di colore verde contenenti i pigmenti clorofilla a, b e carotenoidi (e ficobiline nei plastidi fotosintetici delle alghe rosse); -i cromoplasti di colore giallo, arancio o rosso mancano di clorofilla ma hanno carotenoidi; -i leucoplasti incolori per l’assenza di pigmenti. Come nei procarioti e nei mitocondri, i plastidi presentano una o più molecole di DNA e ribosomi deputati alla sintesi proteica. Si duplicano per scissione. I diversi plastidi possono trasformarsi l’uno nell’altro e tutti derivano dalla stessa forma embrionale, il proplastidio. Attività metaboliche, come la fotosintesi, la biosintesi degli acidi grassi, degli amminoacidi e dell’amido, si svolgono all’interno dei plastidi. a) Cloroplasti b) Cromoplasti c) Leucoplasti.
; -i cromoplasti di colore giallo, arancio o rosso mancano di clorofilla ma hanno carotenoidi; -i leucoplasti incolori per l’assenza di pigmenti. Come nei procarioti e nei mitocondri, i plastidi presentano una o più molecole di DNA e ribosomi deputati alla sintesi proteica. Si duplicano per scissione. I diversi plastidi possono trasformarsi l’uno nell’altro e tutti derivano dalla stessa forma embrionale, il proplastidio. Attività metaboliche, come la fotosintesi, la biosintesi degli acidi grassi, degli amminoacidi e dell’amido, si svolgono all’interno dei plastidi. a) Cloroplasti b) Cromoplasti. c) Leucoplasti.")
27
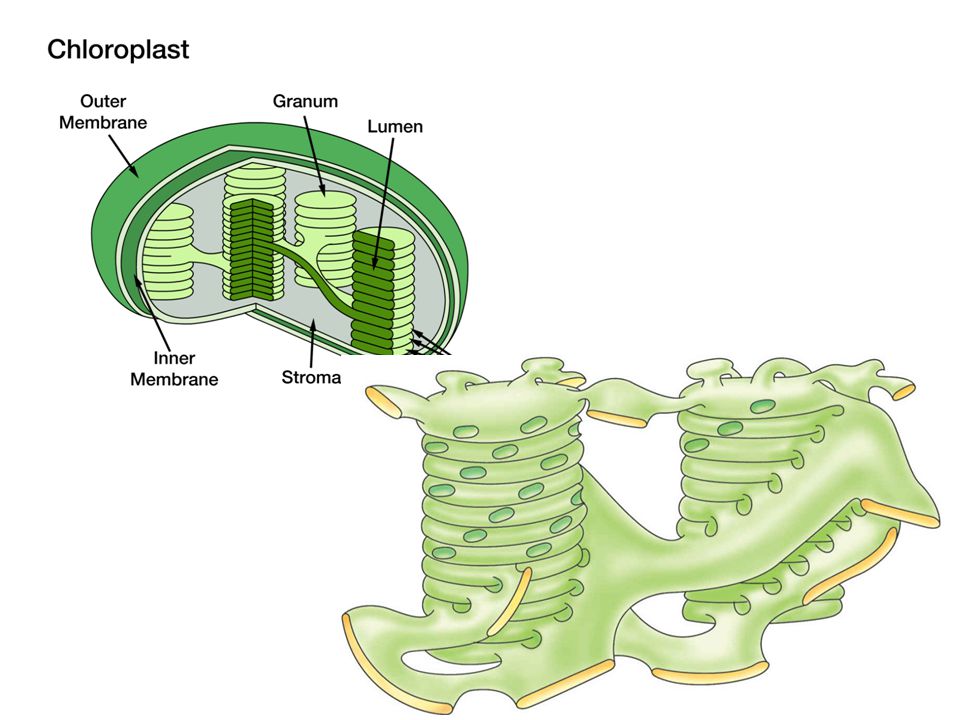
A differenza di tutti gli altri compartimenti citoplasmatici, l’involucro dei plastidi è costituito da un insieme di due membrane unitarie, la membrana esterna e la membrana interna; il compartimento tra le due membrane è detto spazio intermembrana. I cloroplasti sono gli organelli in cui avviene la fotosintesi e sono presenti nelle cellule delle parti verdi della pianta. Sono di colore verde in quanto assorbono, e quindi utilizzano, solo la luce rossa e blu. I pigmento in essi contenuto è principalmente la clorofilla, ma si riscontrano anche carotenoidi Internamente al cloroplasto si trova un ulteriore compartimento membranoso che va a costituire i tilacoidi; questi sono immersi in una sostanza amorfa, detta stroma. I tilacoidi si dispongono in pile a formare i grana. Nelle membrane dei grana avvengono i processi di fotosintesi e contengono clorofilla. Nello stroma risiedono gli enzimi coinvolti nell’organicazione del carbonio. Es. la clorofilla, nei cloroplasti, si forma solo in presenza della luce; i cloroplasti si trovano solo nelle parti della pianta che sono esposte alla luce. Foglia sviluppata al buio: i suoi proplastidi non diventano leucoplasti, ma sono sempre cloroplasti incolori ed anomali

30
La fotosintesi può essere rappresentata da questa equazione generale:
6CO2 + 6H2O + energia → C6H12O6 + 6O2 che corrisponde a: anidride carbonica + acqua + energia solare → glucosio + ossigeno In realtà, il processo di fotosintesi è molto più articolato e riunisce una lunga serie di reazioni complesse, suddivise in una fase luminosa e in una fase oscura.

31
La fotosintesi avviene nelle parti verdi della pianta, in particolare nelle foglie. Qui abbiamo uno schema dei tessuti di una foglia Dicotiledone con epidermide superiore e ep. inferiore con gli stomi; nel mesofillo cellule con cloroplasti e fasci vascolari con cellule a palizzata

33
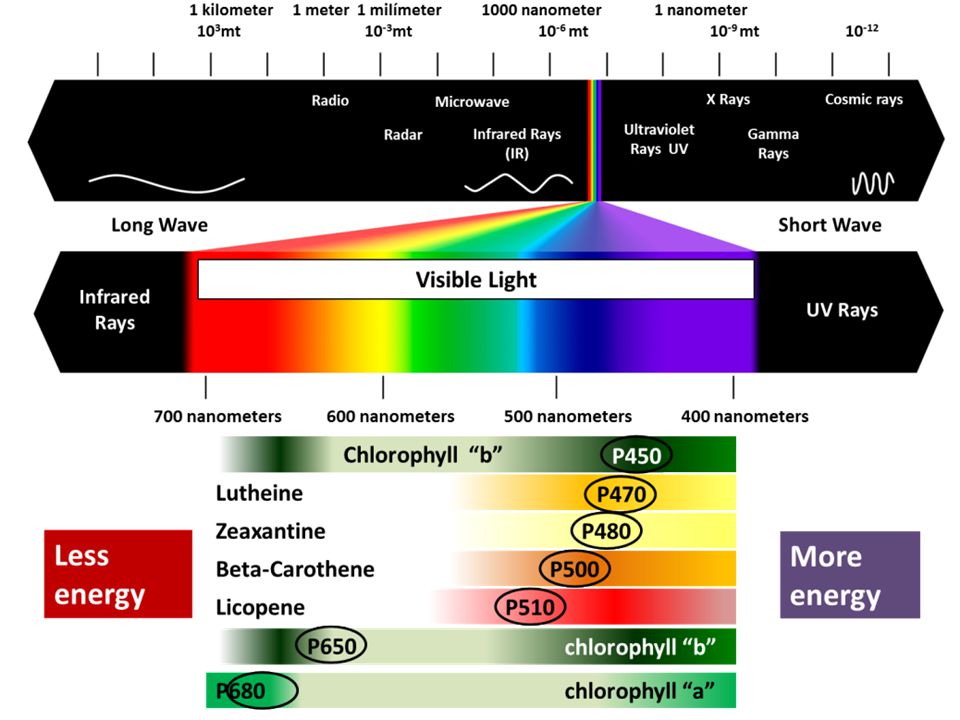
La fase luminosa avviene sulla membrana dei grana e trasforma energia luminosa in energia chimica. La clorofilla, con la collaborazione di altri pigmenti (fotosistemi), viene eccitata dalla luce blu e rossa e attiva la scissione dell’acqua determinando la liberazione di ossigeno molecolare, protoni ed elettroni. La molecola della clorofilla è particolare e ricorda quella della emoglobina, con il Mg al posto del ferro. Each of these differently-colored pigments can absorb a slightly different color of light and pass its energy to the central chlorphyll molecule to do photosynthesis. The central part of the chemical structure of a chlorophyll molecule is a porphyrin ring, which consists of several fused rings of carbon and nitrogen with a magnesium ion in the center. H2O→ 2 e- + 2 H+ + ½ O2 Questa scomposizione dell’acqua, che avviene grazie all’energia della luce, viene denominata fotolisi.

34
I protoni ed elettroni liberati dall’acqua servono per la produzione di una molecola ad alto potere riducente (NADPH) e per la sintesi di ATP. La prima ridurrà il C dell’anidride carbonica , mentre la seconda fornirà l’energia per la formazione dei legami carbonio carbonio necessari per la sintesi di precursori del glucosio (molecola a tre atomi di carbonio , gliceraldeide-3-fosfato (piante C3). Questa seconda serie di reazioni che avvengono nello stroma del cloroplasto, vanno a costituire la fase oscura della fotosintesi
. Questa seconda serie di reazioni che avvengono nello stroma del cloroplasto, vanno a costituire la fase oscura della fotosintesi.")
35
Accanto alle piante C3 esistono anche piante (granoturco) indicate come C4. Queste risultano meglio adattate al caldo e, pur mantenedo gli stomi quasi chiusi per impedire la perdita di acqua, hanno sviluppato sistemi che consentono di catturare e avere una maggior quantità di anidride carbonica. In particolare nel citosol delle cellule con cloroplasti le C4 hanno un particolare enzima che può, anche a livelli bassi di CO2 , sintetizzare un composto a quattro atomi di C detto ossalacetato . L’ossalacetato non viene usato per la sintesi di glucosio, ma degradato per fornire CO2 . Per questo motivo queste piante rimangono verdi quando le altre muoiono. Esistono anche piante CAM La fotosintesi avviene nel mesofillo - cloroplasti - clorofilla Nel cloroplasto: assorbimento della luce, ossidazione H2O Þ O2, riduzione CO2 Þ C6H12O6 Reazioni luminose (nei tilacoidi) prodotto finale Þ ATP, NADPH Reazione al buio (nello stroma) prodotto finale Þ zuccheri Fotosistemi (sistemi di cattura) Trasferimento e- Þ NADP Þ NADPH l Þ Forza motrice protonica membrana tilacoidale Þ gradiente H+ sintesi ATP Ciclo delle CAM Durante la notte: apertura degli stomi fissazione della CO2 da parte della PEPcarbossilasi con formazione dell'acido ossalacetico la PEPcarbossilasi è inibita durante il giorno dall'acido malico l'acido ossalacetico è poi ridotto ad acido malico (enzima malico deidrogenasi NADH dipendente) l'acido malico a pH cellulare si scinde in ione malato- e 2 protoni H+ , entrambi vengono accumulati nel vacuolo il trasporto notturno nel vacuolo è contro gradiente di concentrazione richiede quindi un dispendio di ATP trasporto attivo primario di protoni H+ trasporto attivo secondario di malato l'accumulo di acido malico nel vacuolo determina la caratteristica acidità notturna della pianta, determina inoltre una costante diminuzione notturna del potenziale idrico, che minimo al mattino, consente l'assorbimento di rugiada da parte di radici superficiali Durante il giorno chiusura degli stomi per evitare lo stress idrico per l'aumentata concentrazione di CO2 (stimolo che chiude gli stomi) non si forma più acido malico nel citosol, la pompa protonica verso il vacuolo si arresta l'acido malico accumulato nel vacuolo esce per diffusione l'acido malico viene decarbossilato con formazione di piruvato e di CO2 la CO2 entra nel ciclo di Calvin il piruvato viene trasformato in PEP (così riparte il ciclo)
 prodotto finale Þ ATP, NADPH Reazione al buio (nello stroma) prodotto finale Þ zuccheri Fotosistemi (sistemi di cattura) Trasferimento e- Þ NADP Þ NADPH. l Þ Forza motrice protonica membrana tilacoidale Þ gradiente H+ sintesi ATP. Ciclo delle CAM Durante la notte: apertura degli stomi. fissazione della CO2 da parte della PEPcarbossilasi con formazione dell acido ossalacetico. la PEPcarbossilasi è inibita durante il giorno dall acido malico. l acido ossalacetico è poi ridotto ad acido malico (enzima malico deidrogenasi NADH dipendente) l acido malico a pH cellulare si scinde in ione malato- e 2 protoni H+ , entrambi vengono accumulati nel vacuolo. il trasporto notturno nel vacuolo è contro gradiente di concentrazione richiede quindi un dispendio di ATP. trasporto attivo primario di protoni H+ trasporto attivo secondario di malato. l accumulo di acido malico nel vacuolo determina la caratteristica acidità notturna della pianta, determina inoltre una costante diminuzione notturna del potenziale idrico, che minimo al mattino, consente l assorbimento di rugiada da parte di radici superficiali Durante il giorno. chiusura degli stomi. per evitare lo stress idrico. per l aumentata concentrazione di CO2 (stimolo che chiude gli stomi) non si forma più acido malico nel citosol, la pompa protonica verso il vacuolo si arresta. l acido malico accumulato nel vacuolo esce per diffusione. l acido malico viene decarbossilato con formazione di piruvato e di CO2. la CO2 entra nel ciclo di Calvin. il piruvato viene trasformato in PEP (così riparte il ciclo)")
37
Le piante C3 fissano la CO2 attraverso il ciclo di Calvin, questo prevede il legame con il ribulosio-1,5-difosfato, catalizzato dall'enzima RuBisCO che può addizionare al ribulosio o la CO2 oppure l'O2, che competono tra loro per il sito attivo dell'enzima; in questo secondo caso si ha il fenomeno della FOTORESPIRAZIONE con ossidazione del substrato senza guadagno di energia e diminuzione dell'efficienza della fissazione del C. Le piante C4 riducono la fotorespirazione attraverso un particolare meccanismo; queste piante hanno una particolare anatomia della foglia con due tipi di cellule: mesofillo e guaina del fascio attorno ai fasci vascolari. Nelle cellule del mesofillo avviene la fissazione della CO2 con formazione di un prodotto ossalacetato ad opera di un enzima che ha una maggiore affinità per la CO2 rispetto alla RuBisCO. L’ossalacetato passa nelle cellule della guaina del fascio, dove rilascia la CO2 che reagisce con il ribulosio difosfato, iniziando il ciclo di Calvin: visto che l'ossalacetato "pompa" CO2 dal mesofillo alle cellule della guaina, qui la concentrazione di CO2 è molto maggiore rispetto a quella dell'O2 e così la fotorespirazione è molto ridotta e l'efficienza di fissazione del C è molto elevata. Le piante CAM invece possono vivere in ambienti con poca acqua, perché aprono i loro stomi (per far entrare la CO2, ma questo fa aumentare la perdita di acqua da parte delle foglie) di notte, quando la temperatura è più bassa e quindi l'evaporazione dell'acqua è ridotta. In pratica queste piante svolgono la fase luminosa di giorno (ovviamente) e la fase oscura di notte. Crassulacean acid metabolism

38
Cosa succede all’ossigeno molecolare liberato dai cloroplasti
Cosa succede all’ossigeno molecolare liberato dai cloroplasti? E al glucosio? L’ossigeno in parte si libera dagli stomi, ma parte serve ai mitocondri per la fosforilazione ossidativa e produzione di ATP che servirà alle cellule per tutte le altre reazioni sintetiche o altro. Il glucosio in eccesso viene temporaneamente polimerizzato a formare granuli d’amido (AMIDO PRIMARIO) all’interno dei cloroplasti stessi. Quando è in quantità viene risolubilizzato e trasferito in vacuoli e/o organi di accumulo Anche i cloroplasti ,come i mitocondri, sono organelli semiautonomi nello stroma hanno DNA RNA ed avvengono replicazione, trascrizione e traduzione. Come i mitocondri vengono ereditati per linea femminile (c’è qualche eccezione)
 all’interno dei cloroplasti stessi. Quando è in quantità viene risolubilizzato e trasferito in vacuoli e/o organi di accumulo. Anche i cloroplasti ,come i mitocondri, sono organelli semiautonomi nello stroma hanno DNA RNA ed avvengono replicazione, trascrizione e traduzione. Come i mitocondri vengono ereditati per linea femminile (c’è qualche eccezione)")
Presentazioni simili





















 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")
