Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Genetica dei Procarioti - 4
Dipartimento di Scienze della Vita CdS Biologia Molecolare e Cellulare (LM-6) A.A Genetica dei Procarioti - 4 Prof. Laura Marri ricevimento: previo appuntamento telefonico o
 A.A Genetica dei Procarioti - 4. Prof. Laura Marri. ricevimento: previo appuntamento telefonico o .")
2
Frequenza di mutazione
La frequenza con cui avvengono i diversi tipi di mutazioni è estremamente variabile. Errori nella replicazione del DNA ricorrono con una frequenza di per coppia di basi per singolo ciclo di replicazione. Aumenta in fase stazionaria: mutazione di una cellula ogni 300–400

3
Una coltura di E. coli in terreno complesso (LB) entra in fase di morte dopo tre giorni di incubazione. In questa fase, il 90-99% della popolazione muore 90-99% ossidazione di proteine e DNA, programma simile all’apoptosi eucariotica che determina la morte delle cellule al raggiungimento di una densità elevata

4
Le cellule sopravvissute possono utilizzare le sostanze rilasciate da quelle morte, inclusi gli amminoacidi dalle proteine, i lipidi dalla membrana cellulare e il DNA Dopo la fase di morte la vitalità delle cellule può essere mantenuta per mesi o addirittura per anni in una condizione definita fase stazionaria prolungata A partire dal 10° giorno delle colture in LB si osserva la comparsa, all’interno della popolazione batterica, del fenotipo GASP I fenotipi GASP mostrano una fitness migliore rispetto al ceppo parentale e il bilancio tra le cellule in crescita e morte fornisce un equilibrio dinamico il cui risultato finale è la vitalità stabile della popolazione

5
rpoS duplicazione di 46 bp all’estremità 3’ proteina in cui i quattro amminoacidi presenti a C-terminale sono sostituiti con 39 nuovi amminoacidi indebolimento dell’espressione del regulon (in E. coli RpoS controlla l’espressione di oltre il 10% dei geni) sbilanciamento della competizione tra i vari fattori sigma per il core della RNA polimerasi. Cellule con una ridotta espressione di rpoS favorirebbero, infatti, l’associazione dei fattori sigma RpoD (σ70 ) e RpoN (σ54) alla RNA polimerasi determinando una fitness migliore rispetto al ceppo wild-type
 sbilanciamento della competizione tra i vari fattori sigma per il core della RNA polimerasi. Cellule con una ridotta espressione di rpoS favorirebbero, infatti, l’associazione dei fattori sigma RpoD (σ70 ) e RpoN (σ54) alla RNA polimerasi determinando una fitness migliore rispetto al ceppo wild-type.")
6
lrp Lrp è un regolatore trascrizionale che può funzionare sia da attivatore che da repressore e controlla l’espressione di molti geni implicati nel metabolismo e nel trasporto degli amminoacidi maggiore capacità di recuperare gli amminoacidi rilasciati dalle cellule morte, in particolare serina, treonina e alanina Attivazione operone ybeJ-gltJKL riarrangiamento genomico prodotto dalla sequenza di inserzione IS5. Quattro geni di questo operone sono implicati nella regolazione del trasporto per l’aspartato e il glutammato, aumenta il potenziale di assorbimento degli amminoacidi per i mutanti

7
Un modello delle basi fisiologiche del fenotipo GASP
mutanti GASP: “truffatori evolutivi”, ignorano i segnali di inibizione della crescita e continuano a moltiplicarsi Un modello delle basi fisiologiche del fenotipo GASP

8
La competizione osservata all’interno di popolazioni batteriche è stata interpretata facendo riferimento alla “teoria dei giochi” i “cooperatori” (WT-WT) coesistono, entrambi bloccano anticipatamente la crescita e sono premiati attraverso la redistribuzione delle risorse necessarie al mantenimento della popolazione per lungo tempo i “cooperatori” (WT) che incontrano i “truffatori” (GASP) sono imbrogliati e le loro risorse vengono consumate prematuramente dai secondi i “truffatori” che coesistono tra loro (GASP & GASP) sono puniti poichè continuano a crescere senza controllo mettendo a rischio la vitalità a lungo termine
 coesistono, entrambi bloccano anticipatamente la crescita e sono premiati attraverso la redistribuzione delle risorse necessarie al mantenimento della popolazione per lungo tempo. i cooperatori (WT) che incontrano i truffatori (GASP) sono imbrogliati e le loro risorse vengono consumate prematuramente dai secondi. i truffatori che coesistono tra loro (GASP & GASP) sono puniti poichè continuano a crescere senza controllo mettendo a rischio la vitalità a lungo termine.")
9
in un habitat artificiale spazialmente strutturato (micro lastre in silicone) in cui la disponibilità dei nutrienti è omogenea, WT e GASP coesistono
 in cui la disponibilità dei nutrienti è omogenea, WT e GASP coesistono")
11
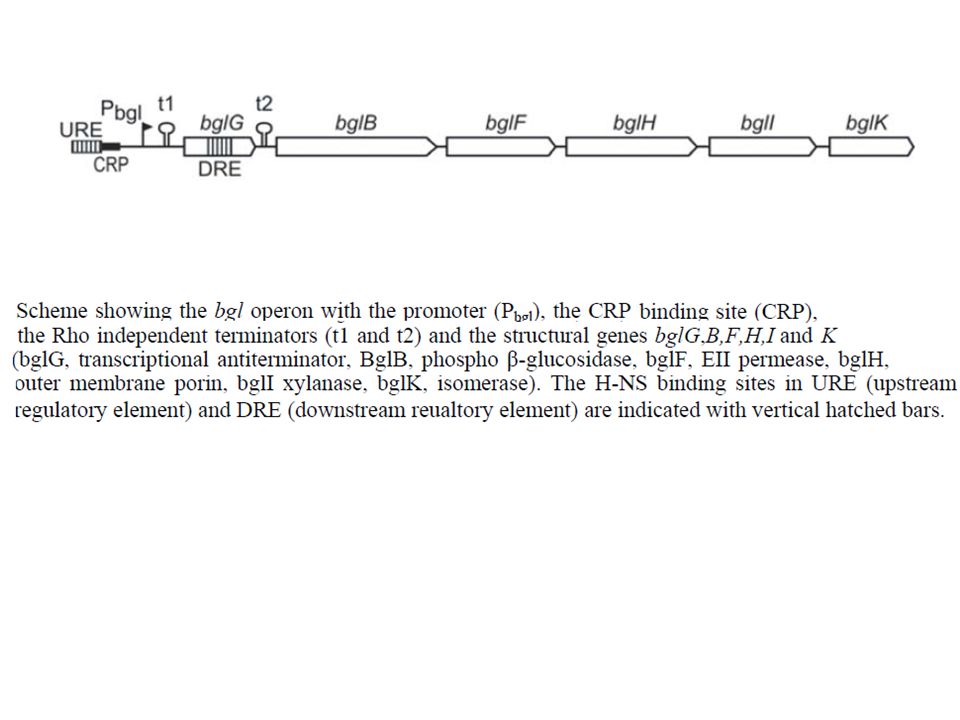
operone criptico sequenze di DNA fenotipicamente silenti normalmente non espresse durante il ciclo vitale dell’organismo Sistemi criptici in Escherichia coli K12 bgl metabolismo arbutina e salicina cel metabolismo arbutina, salicina, cellobiosio arbT assunzione arbutina asc metabolismo arbutina, salicina, cellobiosio

12
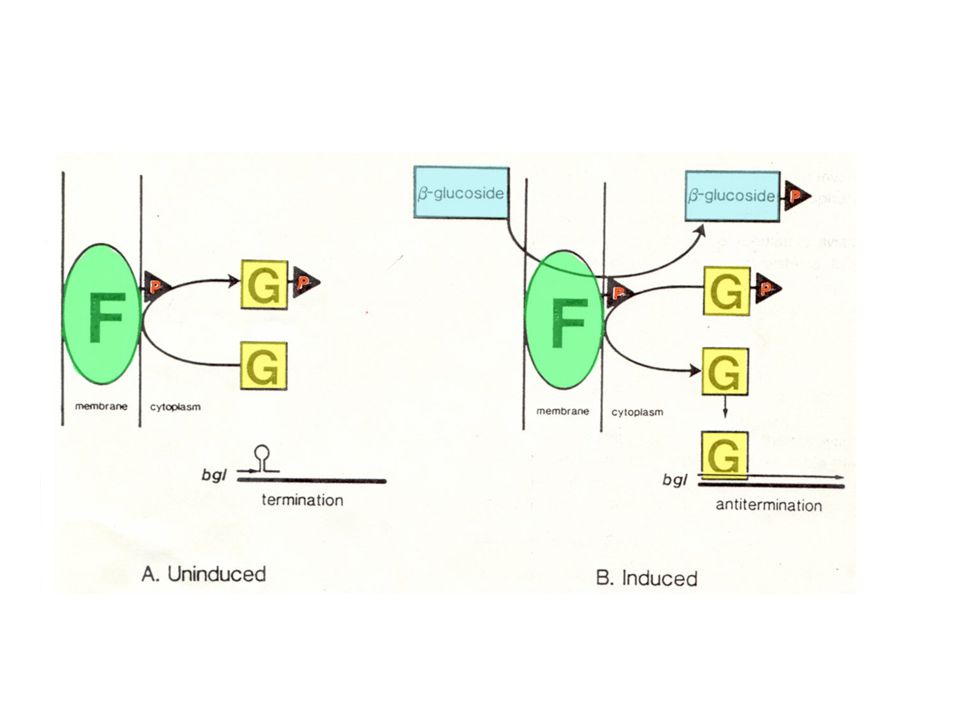
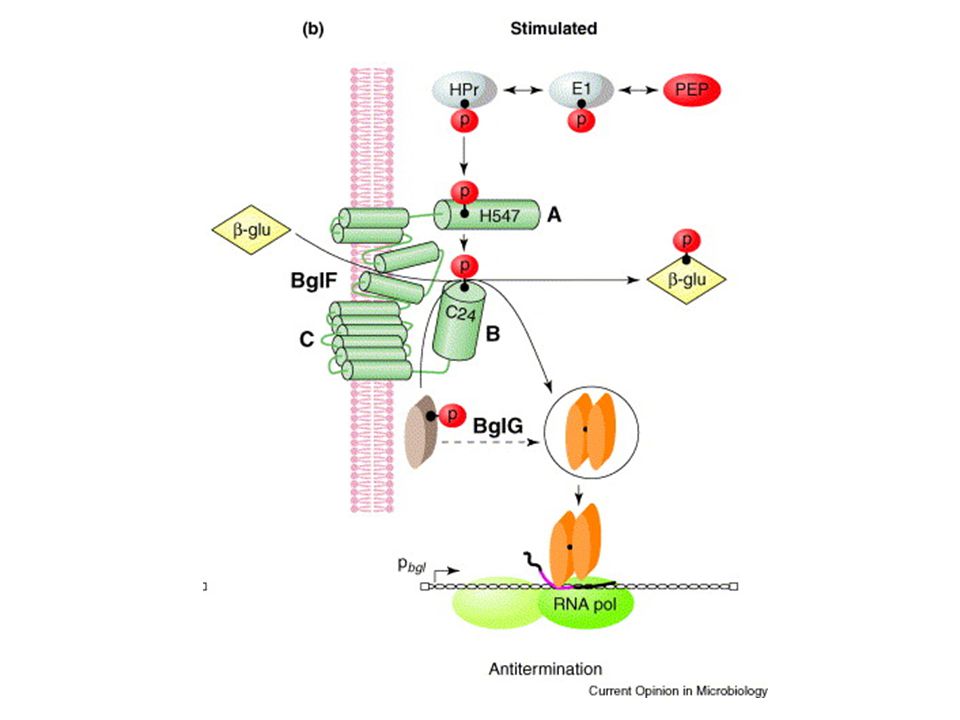
utilizzo beta-glucosidi aromatici
operone criptico Bgl utilizzo beta-glucosidi aromatici PHLORIZIN SALICIN ARBUTIN

14
Operone bgl *

15
H-NS binds to the upstream regulatory element and the promoter
RNA polimerasi CAP-cAMP CF CF CF CF CF CF CF -35 -10 regione AT CAP bglG t1 Pbgl +132 H-NS binds to the upstream regulatory element and the promoter

16
Figure: 07-10d Caption: Supercoiled DNA. (d) In actuality, the double-stranded DNA in the bacterial chromosome is arranged not in one supercoil but in several supercoiled domains, as shown here. In Escherichia coli over 50 supercoiled domains are thought to exist, each of which is stabilized by binding to specific proteins.
 In actuality, the double-stranded DNA in the bacterial chromosome is arranged not in one supercoil but in several supercoiled domains, as shown here. In Escherichia coli over 50 supercoiled domains are thought to exist, each of which is stabilized by binding to specific proteins.")
17
StpA paralogo di H-NS The H-NS paralog StpA shares 58% amino acid identity with H-NS and the two proteins have many common properties. StpA can complement the gene expression defect of the hns single mutant (Shi and Bennet, 1994)
")
18
Antagonistic control of the Escherichia coli bgl promoter by FIS and CAP in vitro
Structure of the bgl (β‐glucoside) operon and the bgl promoter region. Top: The bgl operon encodes a transcriptional antiterminator (encoded by bglG), which acts at terminators t1 and t2, the β‐glucoside‐specific permease EIIBgl of the PTS‐family (encoded by bglF), and a phospho‐β‐glucosidase (encoded by bglB) (Mahadevan and Wright, 1987; Schnetz et al., 1987; Schnetz and Rak, 1988; Amster‐Choder and Wright, 1993). Silencer regions located upstream and downstream of the promoter (Schnetz, 1995) are marked by dotted lines. Bottom: Sequence of the bgl promoter region. The −35 and −10 motifs of the core promoter and the CAP binding site are marked by boxes. The transcriptional start site (+1) is marked by a bent arrow. −66CT indicates one of the point mutations that lead to activation of the bgl promoter by improvement of the CAP binding site (Reynolds et al., 1986). (The second known point mutation that causes activation is −54TC.) The core sequences of FIS binding sites I and II are indicated by black bars between the upper and lower strand of the sequence. Hypersensitive sites that were obtained in the footprints (Fig. 2) are marked by arrows above (upper strand) and below the sequence (lower strand). © This slide is made available for non-commercial use only. Please note that permission may be required for re-use of images in which the copyright is owned by a third party. Caramel A, Schnetz K. Molecular Microbiology Volume 36, Issue 1, pages 85-92, 2002
 operon and the bgl promoter region. Top: The bgl operon encodes a transcriptional antiterminator (encoded by bglG), which acts at terminators t1 and t2, the β‐glucoside‐specific permease EIIBgl of the PTS‐family (encoded by bglF), and a phospho‐β‐glucosidase (encoded by bglB) (Mahadevan and Wright, 1987; Schnetz et al., 1987; Schnetz and Rak, 1988; Amster‐Choder and Wright, 1993). Silencer regions located upstream and downstream of the promoter (Schnetz, 1995) are marked by dotted lines. Bottom: Sequence of the bgl promoter region. The −35 and −10 motifs of the core promoter and the CAP binding site are marked by boxes. The transcriptional start site (+1) is marked by a bent arrow. −66CT indicates one of the point mutations that lead to activation of the bgl promoter by improvement of the CAP binding site (Reynolds et al., 1986). (The second known point mutation that causes activation is −54TC.) The core sequences of FIS binding sites I and II are indicated by black bars between the upper and lower strand of the sequence. Hypersensitive sites that were obtained in the footprints (Fig. 2) are marked by arrows above (upper strand) and below the sequence (lower strand). © This slide is made available for non-commercial use only. Please note that permission may be required for re-use of images in which the copyright is owned by a third party. Caramel A, Schnetz K. Molecular Microbiology Volume 36, Issue 1, pages 85-92,")
19
Insertion of DNA activates the cryptic bgl operon in E. coli K12
293, (22 October 1981) Insertion of DNA activates the cryptic bgl operon in E. coli K12 Ann E. Reynolds, Jeffrey Felton* & Andrew Wright Spontaneous mutations which activate the cryptic bgl operon of Escherichia coli K12 are caused by insertion of DNA at a site, bglR, within the operon. Two insertion elements, IS1 and IS5, have been observed to effect this activation. Once the activating insertion has occurred the operon is inducible by glucosides in a cyclic AMP-dependent manner.
 Insertion of DNA activates the cryptic bgl operon in E. coli K12. Ann E. Reynolds, Jeffrey Felton* & Andrew Wright. Spontaneous mutations which activate the cryptic bgl operon of Escherichia coli K12 are caused by insertion of DNA at a site, bglR, within the operon. Two insertion elements, IS1 and IS5, have been observed to effect this activation. Once the activating insertion has occurred the operon is inducible by glucosides in a cyclic AMP-dependent manner.")
20
Sequence of the bgl promoter and upstream regulatory region.
Indicated are the −35, −10, and transcription start sites of the promoter; the CRP binding site (boxed); and the Fis binding sites (dotted lines), as well as the BglJ-RcsB (solid lines) binding sites.
; and the Fis binding sites (dotted lines), as well as the BglJ-RcsB (solid lines) binding sites.")
21
LeuO, a transcriptional regulator, likewise binds to the bgl upstream regulatory region and relieves repression of bgl independently of BglJ-RcsB. BglJ and RcsB (both TR of the LuxR-type family) interact and form heterodimers that presumably bind upstream of the bgl promoter Promoters (P) are indicated by pointed flags.
 interact and form heterodimers that presumably bind upstream of the bgl promoter. Promoters (P) are indicated by pointed flags.")
22
Operone bgl *

26
Ruolo biologico ed evolutivo dei geni criptici
Secondo il modello di Hall et al. (1987), un gene di una popolazione microbica può trovarsi in tre stati: Allele funzionale; allele criptico; allele non-funzionale. L’allele wild type è favorevole in certi ambienti, mentre in ambienti alternativi, con risorse limitate, l’allele criptico è vantaggioso.
, un gene di una popolazione microbica può trovarsi in tre stati: Allele funzionale; allele criptico; allele non-funzionale. L’allele wild type è favorevole in certi ambienti, mentre in ambienti alternativi, con risorse limitate, l’allele criptico è vantaggioso.")
27
La distribuzione di alleli criptici/funzionali per il metabolismo di β-glucosidi varia all’interno delle Enterobacteriacae in relazione alla nicchia ecologica occupata Klebsiella spp. Gruppo I fermentano β-glucosidi aromatici e cellobiosio Escherichia coli, Shigella sonnei Gruppo II non possono fermentare β-glucosidi, ma possono mutare acquisendo indipendentemente un fenotipo Arb+, Sal+ o Cel+ Il mantenimento di alcuni sistemi criptici rappresenta un sistema di regolazione a lungo termine per funzioni raramente utilizzate In ambienti multi-risorsa, un allele criptico attivato è svantaggioso, ma selettivamente favorevole in particolari condizioni di stress (es. digiuno, risposta immunitaria dell’ospite)
")
28
A model to explain the coexistence of predator and prey in the context of β-glucoside metabolism. (1) Nematodes inhibit amoeba growth, (2) nematodes grow on amoebae, (3) amoebae repel nematodes above a certain density [29], (4) amoebae inhibit bacterial growth, (5) amoebae grow on bacteria, (6) Bgl+ bacteria can inhibit amoeba growth in the presence of β-glucosides, (7) bacteria can grow on dead amoebae, (8) nematodes inhibit bacterial growth, (9) nematodes grow on bacteria, (10) Bgl+ bacteria can inhibit nematode growth in the presence of β-glucosides, (11) bacteria can grow on dead nematodes. (Online version in colour.)
 Nematodes inhibit amoeba growth, (2) nematodes grow on amoebae, (3) amoebae repel nematodes above a certain density [29], (4) amoebae inhibit bacterial growth, (5) amoebae grow on bacteria, (6) Bgl+ bacteria can inhibit amoeba growth in the presence of β-glucosides, (7) bacteria can grow on dead amoebae, (8) nematodes inhibit bacterial growth, (9) nematodes grow on bacteria, (10) Bgl+ bacteria can inhibit nematode growth in the presence of β-glucosides, (11) bacteria can grow on dead nematodes. (Online version in colour.).")
29
Nome/Gene Funzione σ70 (rpoD) σ54 (rpoN) σ38 (rpoS) σ32 (rpoH)
Geni necessari durante la crescita esponenziale σ54 (rpoN) Geni espressi in condizioni di carenza di azoto σ38 (rpoS) Geni espressi durante la fase stazionaria e in condizioni di stress generale σ32 (rpoH) Geni espressi durante heat-shock (chaperoni per ristabilire il folding corretto delle proteine citoplasmatiche) σ28 (rpoF) Geni codificanti per proteine coinvolte nell’assemblaggio del flagello σ24 (rpoE) Geni espressi in condizioni di stress termico per ripristinare l’integrità di membrana e assicurare il folding corretto delle proteine di membrana σ19 (fecI) Geni espressi in relazione al trasporto del citrato ferrico
 Geni espressi in condizioni di carenza di azoto. σ38 (rpoS) Geni espressi durante la fase stazionaria e in condizioni di stress generale. σ32 (rpoH) Geni espressi durante heat-shock (chaperoni per ristabilire il folding corretto delle proteine citoplasmatiche) σ28 (rpoF) Geni codificanti per proteine coinvolte nell’assemblaggio del flagello. σ24 (rpoE) Geni espressi in condizioni di stress termico per ripristinare l’integrità di membrana e assicurare il folding corretto delle proteine di membrana. σ19 (fecI) Geni espressi in relazione al trasporto del citrato ferrico.")
31
Consensus E. coli 32 (rpoH)
")
32
Some sigma factors are always expressed but inactive without a coactivator
Some are only expressed when the genes they regulate are required Some are expressed but degraded Some are expressed but inactive due to anti-sigma factors

33
Nome Funzione σ70 (RpoD) σ54 (RpoN) σ38 (RpoS) σ32 (RpoH) σ28 (RpoF)
principale fattore durante la crescita in condizioni normali σ54 (RpoN) assimilazione dell’azoto σ38 (RpoS) principale fattore durante la fase stazionaria; affamamento σ32 (RpoH) risposta a heat-shock σ28 (RpoF) sintesi del flagello σ24 (RpoE) EC σ19 (FecI) trasporto del ferro
 assimilazione dell’azoto. σ38 (RpoS) principale fattore durante la fase stazionaria; affamamento. σ32 (RpoH) risposta a heat-shock. σ28 (RpoF) sintesi del flagello. σ24 (RpoE) EC. σ19 (FecI) trasporto del ferro.")
34
Anti-sigma Factors Anti-
4/15/2017 Anti- Anti-sigma factors tightly (yet reversibly) bind sigma factors and prevent their activity.
 bind sigma factors. and prevent their activity.")
35
FlgM

36
durante le fasi iniziali della biogenesi flagellare σ28 è mantenuto inattivo dalla stretta associazione con FlgM

37
l’uncino ed il corpo basale sono assemblati FlgM viene secreto attraverso il sistema di trasporto flagellare (T3S) folded form

38
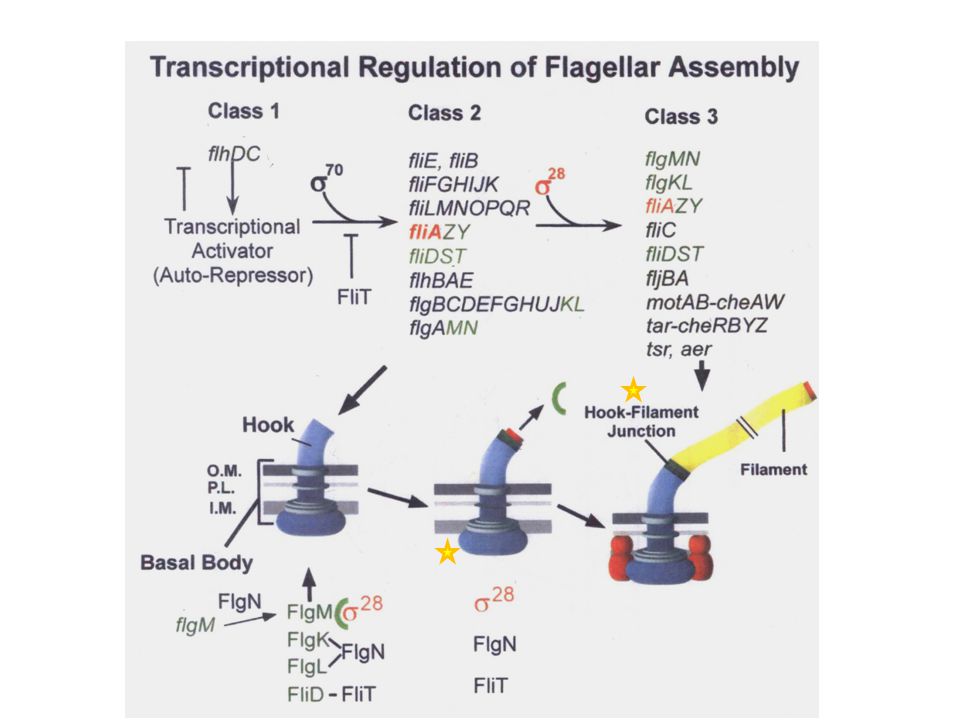
The coupling of flagellar gene regulation to flagellum assembly.
The coupling of flagellar gene regulation to flagellum assembly. The gene products of the flhDC operon activate σ70-dependent transcription from flagellar class 2 promoters, which include everything that is needed for the structure and assembly of the HBB, the ion-powered flagellar rotary motor that transverses from the cytoplasm through the membranes and cell wall to the outside of the cell. Also transcribed from a class 2 promoter is the fliA gene, which encodes the alternative σ transcription factor, σ28. Transcription of class 3 promoters by σ28-RNA polymerase produces all the gene products needed after the HBB motor is constructed. The genes expressed from class 3 promoters include everything that is needed late in the assembly process: flagellin, motor force generators, and a brain, known as the chemosensory system, which allows the flagellum to direct movement of the bacterium up a chemical gradient. The anti-σ28 factor FlgM couples class 3 promoter transcription to HBB completion. Upon HBB completion, the secretion specificity of the flagellar type III secretion system changes from hook/rod-type substrates to late substrates. FlgM, a late secretion substrate, is secreted, and σ28 is free, timed to HBB completion, to transcribe the late flagellar genes and complete flagellum assembly and the chemosensory network. Chevance F F V et al. J. Bacteriol. 2006;188:

40
Some sigma factors are always expressed but inactive without a coactivator
Some are only expressed when the genes they regulate are required Some are expressed but degraded Some are expressed but inactive due to anti-sigma factors

41
Nome Funzione σ70 (RpoD) σ54 (RpoN) σ38 (RpoS) σ32 (RpoH) σ28 (RpoF)
principale fattore durante la crescita in condizioni normali σ54 (RpoN) assimilazione dell’azoto σ38 (RpoS) principale fattore durante la fase stazionaria; affamamento σ32 (RpoH) risposta a heat-shock σ28 (RpoF) sintesi del flagello σ24 (RpoE) EC σ19 (FecI) trasporto del ferro
 assimilazione dell’azoto. σ38 (RpoS) principale fattore durante la fase stazionaria; affamamento. σ32 (RpoH) risposta a heat-shock. σ28 (RpoF) sintesi del flagello. σ24 (RpoE) EC. σ19 (FecI) trasporto del ferro.")
42
Alcuni sRNA legandosì al 5' dell'mRNA impediscono la formazione della struttura secondaria e permettono la traduzione di RpoS (σs). ll sito di legame al ribosoma (RBS) dell'mRNA rpoS è appaiato alla regione 5‘ non tradotta dell'mRNA.
 dell mRNA rpoS è appaiato alla regione 5‘ non tradotta dell mRNA.")
43
Most RpoS expression is determined at the translational level
At least three different small RNAs, DsrA, RprA and OxyS, regulate RpoS translation the abundance and, therefore, the activity of each is in turn regulated by different environmental cues.

44
The stress response sigma factor RpoS has ~500 genes under its control
Default situation: «off», the 5’ folds back to block the chromosome entry

45
Hfq consists of six identical subunits arranged in a ring shape structure
Hfq is an RNA-binding protein encoded in approximately half of the (in 2002) sequenced bacterial genomes and is one of the most abundant proteins in E. coli.
 sequenced bacterial genomes and is one of the most abundant proteins in E. coli.")
46
The typical action of an Hfq-associated sRNA.
The sRNA binds to the Hfq protein and then finds its target mRNA

47
regolazione mediante fattori sigma alternativi

48
sRNA (small RNA), possono svolgere la loro azione regolativa appaiandosi a regioni specifiche di un RNA bersaglio perfettamente o parzialmente complementare (sRNA antisenso) Gli RNA antisenso sono molecole piccole (da poche decine a poche centinaia di nucleotidi), diffusibili
, diffusibili.")
49
pori grandi pori piccoli
OmpF pori grandi in alternativa OmpC pori piccoli livelli di osmolarità livelli di osmolarità

50
MicF OmpC

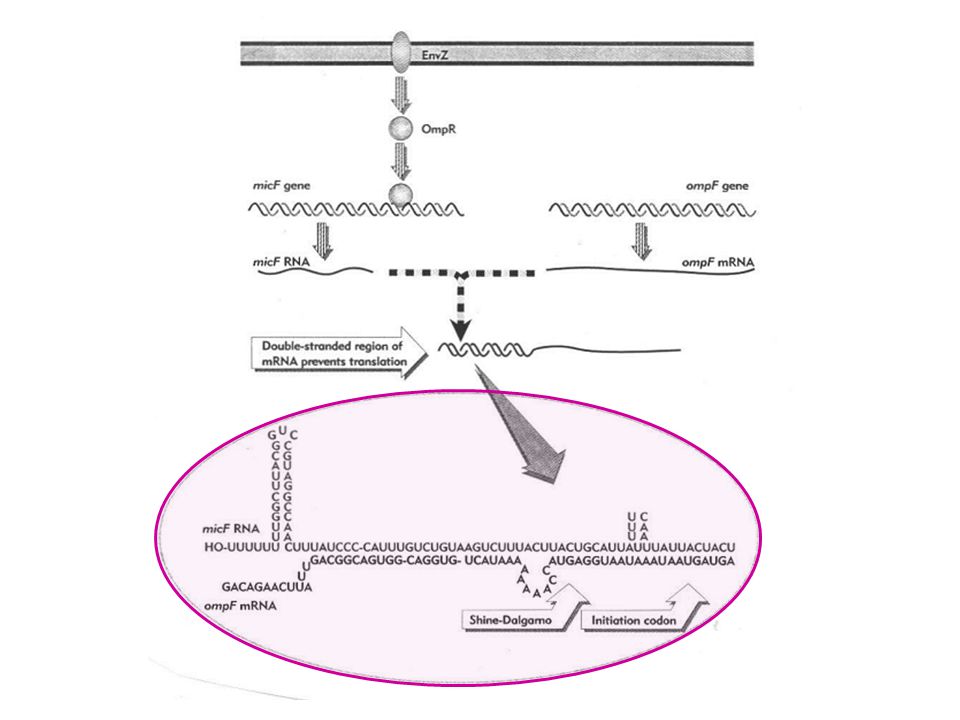
51
Senses changes in extracellular osmolarity
EnvZ/OmpR System of E. coli Senses changes in extracellular osmolarity

Presentazioni simili
















 viene copiata in una sequenza complementare di RNA dall’enzima.>")




 Piccole molecole di RNA (20-22 nt)>")

