Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Micro RNA (miRNA) Piccole molecole di RNA (20-22 nt)
Prodotte da precursori di nt trascritti autonomamente (anche policistronici) o maturati da introni Si legano a mRNA che hanno sequenze complementari Presenti in tutti gli eucarioti
 o maturati da introni. Si legano a mRNA che hanno sequenze complementari. Presenti in tutti gli eucarioti.")
2
Conservazione dei miRNA

3
Strutture secondarie dei pre-miRNA
Figure 5. Predicted secondary structure of putative miRNA precursors and organization of miRNA gene clusters. (A) 70 nt of genomic sequence upsteam and 70 nt downstream of the cloned miRNAs were used to predict the secondary structure of putative miRNA precursors, using the computer program mfold (Mathews et al. 1999). The size of the putative miRNA precursors was then reduced to ∼80 nt, and this sequence was used again for secondary structure prediction by mfold. The putative precursor of miR-91 also contains the sequence of miR-17 on the opposite strand (colored in blue). This finding is similar to the C. elegans miR-56 and miR-56*, which are processed from the same precursor, although only miR-56 is thought to be the functional miRNA (Nelson et al. 2001). Chromosomal location is indicated. Red areas represent mature miRNAs. (B) The putative miRNA precursors are represented as boxes, and the mature miRNAs identified in this study as part of miRNPs are indicated in red. Previously reported miRNAs (Lagos-Quintana et al. 2001) not found in this study are shown in blue. The chromosomal location is indicated on the right. The miRNA cluster on chromosome 13 contains two miRNAs (miR-91 and miR-92) not previously identified; miRNA-91 and miR-17 (colored in blue) are presumably derived from the same precursor. The size of clusters in nucleotides (nt) is shown
 70 nt of genomic sequence upsteam and 70 nt downstream of the cloned miRNAs were used to predict the secondary structure of putative miRNA precursors, using the computer program mfold (Mathews et al. 1999). The size of the putative miRNA precursors was then reduced to ∼80 nt, and this sequence was used again for secondary structure prediction by mfold. The putative precursor of miR-91 also contains the sequence of miR-17 on the opposite strand (colored in blue). This finding is similar to the C. elegans miR-56 and miR-56*, which are processed from the same precursor, although only miR-56 is thought to be the functional miRNA (Nelson et al. 2001). Chromosomal location is indicated. Red areas represent mature miRNAs. (B) The putative miRNA precursors are represented as boxes, and the mature miRNAs identified in this study as part of miRNPs are indicated in red. Previously reported miRNAs (Lagos-Quintana et al. 2001) not found in this study are shown in blue. The chromosomal location is indicated on the right. The miRNA cluster on chromosome 13 contains two miRNAs (miR-91 and miR-92) not previously identified; miRNA-91 and miR-17 (colored in blue) are presumably derived from the same precursor. The size of clusters in nucleotides (nt) is shown.")
4
Organizzazione dei geni di miRNA
Figure 2. miRNA genomic and functional organization. The genomic and functional organization of four miRNA clusters/families is clarified: (A) let-7/mir-98; (B) mir-141/200; (C) mir-21; and (D) mir The genomic locations for each of the miRNA members are defined. Grey lines denote intronic regions. miRNA mature sequences are colour-coded according to the sequence family to which they belong (eg in the let-7 cluster, red signifies the let-7/mir-98 sequence family). The star sequence is defined with a grey bar. The sequence families are depicted as sequence alignments compared to the most highly expressed miRNA family member shown on top, based on profiles of over 1000 human specimens (Tuschl, unpublished data). Shaded residues denote differences from the most highly expressed miRNA family member.
 let-7/mir-98; (B) mir-141/200; (C) mir-21; and (D) mir The genomic locations for each of the miRNA members are. defined. Grey lines denote intronic regions. miRNA mature sequences are colour-coded according to the sequence family to which they. belong (eg in the let-7 cluster, red signifies the let-7/mir-98 sequence family). The star sequence is defined with a grey bar. The sequence. families are depicted as sequence alignments compared to the most highly expressed miRNA family member shown on top, based on. profiles of over 1000 human specimens (Tuschl, unpublished data). Shaded residues denote differences from the most highly expressed. miRNA family member.")
5
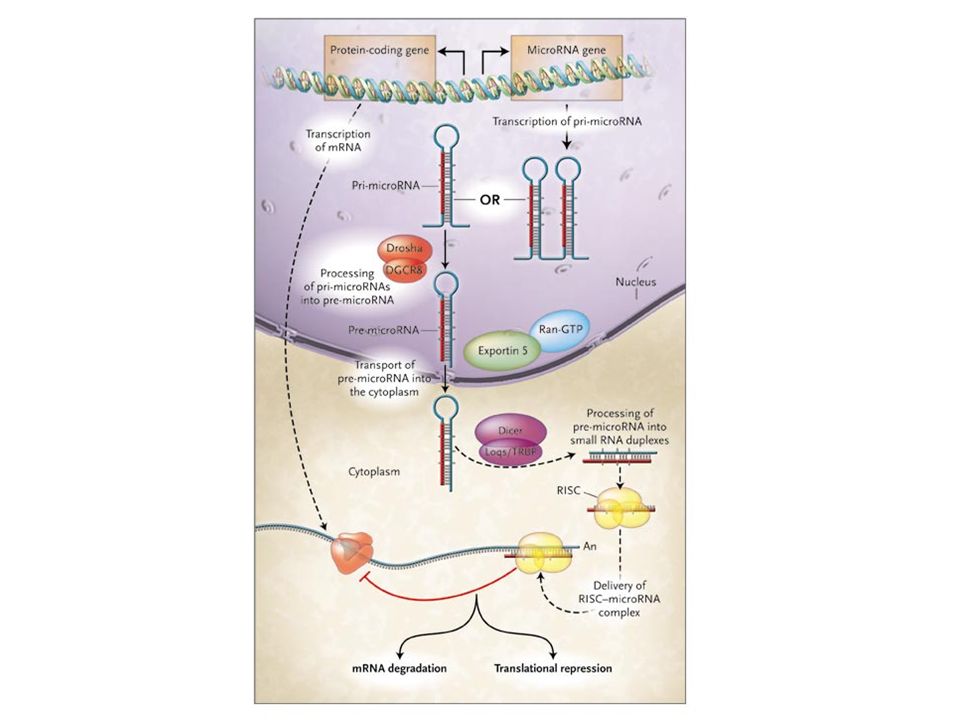
Biogenesi dei miRNA Figure 1. miRNA biogenesis pathway. miRNAs are transcribed by RNAPII to produce pri-miRNAs. Canonical miRNAs are processed by the endoribonuclease Drosha in partnership with its RBP partner DGCR8; mirtrons are instead processed by the spliceosome. The processed pre-miRNA is transported to the cytoplasm through an export complex consisting of Exportin 5. The pre-miRNA is subsequently processed in the cytoplasm by another endoribonuclease, Dicer, in partnership with its RBP partner TRBP, to form the final 21–23 nucleotide miRNA product. miR-451 is not processed by Dicer but is rather cleaved by Ago2. Mature miRNAs (indicated in red) are then incorporated into Ago 1–4, forming miRNPs, also known as miRISC. miRNPs also incorporate other proteins, such as GW182. miRNPs are thought to direct miRNA-mediated destabilization (ie through interaction with CCR4) or miRNA-mediated translational repression (ie through interaction with ribosomes) of miRNAs without perfectly complementary mRNA targets. miRISC is thought to direct Ago2-catalysed target mRNA cleavage of miRNA fully or nearly fully complementary mRNA targets.
 are then incorporated into Ago 1–4, forming miRNPs, also known as miRISC. miRNPs also incorporate other proteins, such as GW182. miRNPs are thought to direct miRNA-mediated destabilization (ie through interaction with CCR4) or miRNA-mediated translational repression (ie through interaction with ribosomes) of miRNAs without perfectly complementary mRNA targets. miRISC is thought to direct Ago2-catalysed target mRNA cleavage of miRNA fully or nearly fully complementary mRNA targets.")
6
Biogenesi dei miRNA

7
Regolazione mediata dai miRNA

8
Studio della funzione dei miRNA

9
Meccanismo di azione dei miRNA
Fig. 2. Proposed mechanisms of microRNA-mediated repression. The majority of microRNA (miRNA)-mediated regulation is due to mRNA degradation. However, the precise mechanism(s) underlying this process remain a topic of lively discussion. Initially, the mature, fully transcribed mRNA is thought be in a circular messenger ribonucleoprotein (mRNP) form composed of, at least, the cap-binding protein (eIF4E), the scaffold protein eIF4G, poly(A) binding protein (PABP), ribosomes and other related proteins not shown here. Once deadenylation, which is mediated by the Pan2-Pan3 and Ccr4-Caf1 deadenylase complexes, has occurred, mRNAs are decapped by the decapping enzyme (not shown) and subsequently degraded by the 5_r3_ exonuclease Xrn1. Current studies aim to elucidate the order and relative contributions of target deadenylation, capdependent translational repression and target decay. Evidence for several potential scenarios has been reported, including: (A) mRNA deadenylation leading to translational repression and mRNA decay; (B) translational repression of target mRNAs leading to deadenylation and decay; (C) decircularization of target mRNAs, which then leads to coincident deadenylation and translational repression, and then to decay; and (D) an unknown/undiscovered mechanism, which occurs coincidently with deadenylation and translational repression.
-mediated regulation is due to mRNA degradation. However, the precise mechanism(s) underlying this process remain a topic of lively discussion. Initially, the mature, fully transcribed mRNA is thought be in a circular messenger ribonucleoprotein (mRNP) form composed of, at least, the cap-binding protein (eIF4E), the scaffold protein eIF4G, poly(A) binding protein (PABP), ribosomes and other related proteins not shown here. Once deadenylation, which is mediated by the Pan2-Pan3 and Ccr4-Caf1 deadenylase complexes, has occurred, mRNAs are decapped by the decapping enzyme (not shown) and subsequently degraded by the 5_r3_ exonuclease Xrn1. Current studies aim to elucidate the order and relative contributions of target deadenylation, capdependent translational repression and target decay. Evidence for several potential scenarios has been reported, including: (A) mRNA deadenylation leading to translational repression and mRNA decay; (B) translational repression of target mRNAs leading to deadenylation and decay; (C) decircularization of target mRNAs, which then leads to coincident deadenylation and translational repression, and then to decay; and (D) an unknown/undiscovered mechanism, which occurs coincidently with deadenylation and translational repression.")
10
Regolazione dei miRNA (regolatori "sponge")
Figure 1. Sponge RNAs compete with target mRNAs for miRNA binding. Sponge RNAs (in red) contain binding sites (grey rectangles) for a miRNA of interest (grey octagons). Left: in the presence of low sponge expression, target mRNAs (in blue) are posttranscriptionally repressed by the miRNA. Right: in the presence of high sponge expression, target mRNAs are relieved of repression; higher protein output (blue ovals) results.
 contain binding sites (grey rectangles) for a miRNA of interest (grey octagons). Left: in the presence of low sponge expression, target mRNAs (in blue) are posttranscriptionally repressed by the miRNA. Right: in the presence of high sponge expression, target mRNAs are relieved of repression; higher protein output (blue ovals) results.")
12
MONDO A RNA (RNA WORLD)
")
13
Diversità degli organismi

14
CARATTERISTICHE DI UN "SISTEMA VIVENTE"
® replicazione ® evoluzione

15
DOGMA CENTRALE DNA RNA proteine

16
CAPACITA' DELL'RNA ® RNA come genoma ® RNA come catalizzatore

17
RNA come genoma

18
RNA come enzima Number of sequenced examples is a snapshot as of 2002 and is influenced by DNAsequencing strategies and database upkeep; it may provide a rough indication of relative abundance. RNAs in any group vary in size; the size provided here indicates the lower end of the length distribution for the natural examples. See and *Ribonucleoprotein enzymes. RNase P: bacterial and archaebacterial RNAs have the relevant activity in the absence of protein. Spliceosome: U2 and U6 small nuclear RNAs (snRNAs) alone show an activity related to the natural activity. Ribosome: no activity has yet been observed with protein-free, large-subunit rRNA.
 alone show an activity related to the natural activity. Ribosome: no activity has yet been observed with protein-free, large-subunit rRNA.")
19
Apparecchio di Miller (1953)
")
20
Sintesi prebiotiche Figure 2 Prebiotic clutter surrounding RNA. Each of the four components of RNA (coloured green, red, purple and blue) would have been accompanied by several closely related analogues (listed in black type), which could have assembled in almost any combination. All possible building blocks for each of the components should be regarded as sorting independently; for example, the phosphodiester linkage may have comprised either a 38,58 linkage involving a phosphate or a 28,58 linkage involving a pyrophosphate.
 would have been accompanied by several closely related analogues (listed in black type), which could have assembled in almost any combination. All possible building blocks for each of the components should be regarded as sorting independently; for example, the phosphodiester linkage may have comprised either a 38,58 linkage involving a phosphate or a 28,58 linkage involving a pyrophosphate.")
21
DIFFICOLTA' DELLE SINTESI PREBIOTICHE
® rese basse ® ribosio instabile (isomeri) ® legame glicosidico a o b ® fosfato solo al 3' e 5' ® legami fosfodiesterici solo 5'-3'
 ® legame glicosidico a o b. ® fosfato solo al 3 e 5 ® legami fosfodiesterici solo 5 -3")
22
Sintesi dei precursori del DNA

23
Cofattori di enzimi proteici

24
Origine dei cofattori

25
FUNZIONI DELL'RNA NELLA CELLULA MODERNA
® traduzione rRNA, mRNA, tRNA ® maturazione rRNA snoRNA, RNasi MRP ® splicing snRNA, introni gruppi I e II ® maturazione tRNA RNasi P ® sintesi DNA primers, telomerasi ® traslocaz. proteine srpRNA

26
PERCHE' L'RNA? ® l'RNA deve essere venuto prima del DNA
l'RNA deve essere venuto prima delle proteine ® molti coenzimi hanno un ribonucleotide nella struttura l'RNA può agire come catalizzatore

27
EVOLUZIONE SINTESI PROTEICA
1) tRNA 2) tRNA sintetasi 3) protoribosoma 4) codice genetico 5) mRNA 6) proteine
 tRNA. 2) tRNA sintetasi. 3) protoribosoma. 4) codice genetico. 5) mRNA. 6) proteine.")
28
PASSAGGIO RNA DNA 1) sintesi dei precursori del DNA
2) trascrizione inversa 3) replicazione 4) trascrizione
 trascrizione inversa. 3) replicazione. 4) trascrizione.")
29
Scala temporale dell'evoluzione
Figure 1 Timeline of events pertaining to the early history of life on Earth, with approximate dates in billions of years before the present.

30
Evoluzione prebiotica

32
(SYSTEMATIC EVOLUTION OF LIGANDS BY EXPONENTIAL ENRICHMENT)
SELEX (SYSTEMATIC EVOLUTION OF LIGANDS BY EXPONENTIAL ENRICHMENT) es. affinity column
 es. affinity column.")
33
Primo esperimento SELEX
FIG. 1. The sequences and reasonable secondary structures of two RNAs that bind with identical affinities to bacteriophage T4 DNA polymerase are shown. The green nucleotides in the major variant are the positions of difference between the two RNAs.

34
Selezione di un RNA con capacità di legame

35
Specificità del legame

36
Selezione di una RNA polimerasi
Figure 4 Successive phases in the in vitro evolution of an RNA polymerase ribozyme. a, Class I ligase ribozyme catalyses template-directed joining of the 38 end of an RNA primer (open line) to the 58 end of the ribozyme44. b, Class I ligase also catalyses addition of three nucleoside 58-triphosphates (NTPs) to the 38 end of the primer, directed by an internal template46. c, Class I-derived polymerase catalyses addition of up to 14 NTPs on an external RNA template5
 to the 58 end of the ribozyme44. b, Class I ligase also catalyses addition of three nucleoside 58-triphosphates (NTPs) to the 38 end of the primer, directed by an internal template46. c, Class I-derived polymerase catalyses addition of up to 14 NTPs on an external RNA template5.")
38
Figure 5 A model for the mechanism of RNAi
Figure 5 A model for the mechanism of RNAi. Silencing triggers in the form of doublestranded RNA may be presented in the cell as synthetic RNAs, replicating viruses or may be transcribed from nuclear genes. These are recognized and processed into small interfering RNAs by Dicer. The duplex siRNAs are passed to RISC (RNA-induced silencing complex), and the complex becomes activated by unwinding of the duplex. Activated RISC complexes can regulate gene expression at many levels. Almost certainly, such complexes act by promoting RNA degradation and translational inhibition. However, similar complexes probably also target chromatin remodelling. Amplification of the silencing signal in plants may be accomplished by siRNAs priming RNA-directed RNA polymerase (RdRP)-dependent synthesis of new dsRNA. This could be accomplished by RISC-mediated delivery of an RdRP or by incorporation of the siRNA into a distinct, RdRP-containing complex.
, and the complex becomes activated by unwinding of the duplex. Activated RISC complexes can regulate gene expression at many levels. Almost certainly, such complexes act by promoting RNA degradation and translational inhibition. However, similar complexes probably also target chromatin remodelling. Amplification of the silencing signal in plants may be accomplished by siRNAs priming RNA-directed RNA polymerase (RdRP)-dependent synthesis of new dsRNA. This could be accomplished by RISC-mediated delivery of an RdRP or by incorporation of the siRNA into a distinct, RdRP-containing complex.")
40
Scheme for the function of different snmRNAs by targeting bacterial or eukaryal mRNAs or pre-mRNAs leading to regulation of gene expression. Binding of a snoRNA to the splice-site of a eukaryal pre-mRNA or involvement in mRNA editing and modification has recently been proposed as one of several alternatives for a functional role, but remains to be experimentally verified.

Presentazioni simili













>")




 viene copiata in una sequenza complementare di RNA dall’enzima.>")

