Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La cellula vegetale

2
Organismi autotrofi Organismi eterotrofi
= produttori della biosfera Autonomi per la nutrizione. Utilizzano composti inorganici. 6H2O + 6CO2 + ENERGIA C6H12O6 + O2 Organismi eterotrofi ‘ consumatori della biosfera Dipendono da altre forme di vita per la propria nutrizione. Utilizzano composti organici. C6H12O6 + O2 6H2O + 6CO2 + ENERGIA

3
Alcune differenze esistenti tra autotrofi ed eterotrofi
Fonte di energia Fissazione CO2 Fissazione N2 Esempio FOTOAUTOTROFI LUCE SI cianobatteri NO piante CHEMIOAUTOTROFI COMPOSTI INORGANICI solfobatteri FOTOETEROTROFI solo per produrre ATP alcuni procarioti CHEMIOETEROTROFI COMPOSTI ORGANICI funghi e animali AUTOTROFI ETEROTROFI

4
Separazione tra cell animale e vegetale è avvenuta tardi, molte caratteristiche della cellula già stabilite: - notevoli differenze con i procarioti - lieviti piante e animali hanno caratteristiche comuni

5
Ipotesi endosimbiontica
Endocitosi: quando O2 nell’atmosfera in quantità sostanziali Durante l’evoluzione trasferimento di geni del DNA mitocondri e cloroplasti a quello nucleare

6
Parete cellulare Vacuolo Cloroplasto

7
LA PARETE CELLULARE Matrice extracellulare complessa che circonda ogni cellula di una pianta, più forti e più rigide della matrice extracellulare delle cellule animali Ruolo strutturale o di “scheletro”, ruolo protettivo (involucro cellulare), importanti funzioni nel trasporto dei fluidi all’interno della pianta A seconda dello stadio di sviluppo cellulare: Primaria: sottile e semirigida, 60% acqua Secondaria: rendendo più spessa la parete primitiva o depositando nuovi strati di composizione diversa sotto i vecchi. Poca acqua, lunghe fibre di cellulosa Le pareti cellulari possono differire per composizione e organizzazione. Principio strutturale comune a materiali compositi (cemento armato): - componente dà forza elastica - materiale nel quale è immerso dà resistenza alla compressione Pareti cellulari al microscopio elettronico
, importanti funzioni nel trasporto dei fluidi all’interno della pianta. A seconda dello stadio di sviluppo cellulare: Primaria: sottile e semirigida, 60% acqua. Secondaria: rendendo più spessa la parete primitiva o depositando nuovi strati di composizione diversa sotto i vecchi. Poca acqua, lunghe fibre di cellulosa. Le pareti cellulari possono differire per composizione e organizzazione. Principio strutturale comune a materiali compositi (cemento armato): - componente dà forza elastica - materiale nel quale è immerso dà resistenza alla compressione. Pareti cellulari al microscopio elettronico.")
8
FORZA DI TENSIONE DELLA PARETE
Fibre che resistono alla tensione: presenza di lunghe fibre di un polisaccaride: CELLULOSA Cellulosa: [cellobiosio]n = catena di almeno 500 residui di glucosio legati mediante legame b-glicosidico.

9
Legami H intermolecolari tra molecole di cellulosa adiacenti fasci di catene con stessa polarità = microfibrille di cellulosa, organizzati in strati o lamelle parallele

10
RESISTENZA ALLA COMPRESSIONE
Matrice in cui sono immerse le microfibrille di cellulosa Proteine strutturali, particolarmente ricche in aa idrossiprolina (collageno) Polisaccaridi: - Glicani: ossatura lineare di 1 zucchero (xilosio, glucosio, mannosio) da cui sporgono catene lineari di altri zuccheri. Formano legami crociati con microfibrille di cellulosa. Pectine: contengono residui di ac.galatturonico (carichi -), molto idratate e associate a nuvola cationica (Ca++), che contribuisce a tenere unite le componenti della parete (gelatina: pectina aggiunta a succo di frutta) - Emicellulose: gruppo eterogeneo di polisaccaridi ramificati che organizzano le microfibrille di cellulosa in una rete complessa - Lignina: polimeri fenolici, polimero addizionale più comune nelle pareti cellulari secondarie. Occupa intersizi rendendo pareti rigide e permanenti
 Polisaccaridi: - Glicani: ossatura lineare di 1 zucchero (xilosio, glucosio, mannosio) da cui sporgono catene lineari di altri zuccheri. Formano legami crociati con microfibrille di cellulosa. Pectine: contengono residui di ac.galatturonico (carichi -), molto idratate e associate a nuvola cationica (Ca++), che contribuisce a tenere unite le componenti della parete (gelatina: pectina aggiunta a succo di frutta) - Emicellulose: gruppo eterogeneo di polisaccaridi ramificati che organizzano le microfibrille di cellulosa in una rete complessa. - Lignina: polimeri fenolici, polimero addizionale più comune nelle pareti cellulari secondarie. Occupa intersizi rendendo pareti rigide e permanenti.")
11
- maggior parte delle macromolecole della matrice è sintetizzata nel ER e nell’apparato di Golgi.
La cellulosa è sintetizzata è “filata” dal sistema enzimatico della cellulosa sintasi, proteina legata alla membrana: precursori citosolici uniti a zuccheri dei nucleotidi (UDP-glucosio) cellulosa liberata sulla superficie extracellululare della membrana, catene nascenti si assemblano spontaneamente in microfibrille Catene nascenti crescono internamente alle precedenti - Orientamento microfibrille influenza il modo in cui la cellula si espande la forma finale della cellula e i fenomeni di divisione cellulare. - Schiera corticale citoplasmatica di microtubuli determina l’orientamento delle microfibrille di cellulosa appena depositate Microtuboli sono necessari per cambio di direzione dello schema di microfibrille
 cellulosa liberata sulla superficie extracellululare della membrana, catene nascenti si assemblano spontaneamente in microfibrille. Catene nascenti crescono internamente alle precedenti. - Orientamento microfibrille influenza il modo in cui la cellula si espande la forma finale della cellula e i fenomeni di divisione cellulare. - Schiera corticale citoplasmatica di microtubuli determina l’orientamento delle microfibrille di cellulosa appena depositate Microtuboli sono necessari per cambio di direzione dello schema di microfibrille.")
12
- Sintesi microfibrille è indipendente dai microtubuli della schiera corticale
- Microtubuli possono cabiare orientamento microfibrille agendo come “argini”

13
I VACUOLI - contenuti nella maggior parte delle cellule vegetali e fungine - uno o più vacuoli per cellula - organuli molto versatili - singola membrana: tonoplasto - occupano dal 5% al 90% del volume della cellula

14
--> vacuoli con funzioni diverse sono presenti nella stessa cellula
- correlati ai lisosomi delle cellule animali, contengono varietà di enzimi idrolitici, hanno però anche funzioni molto diverse: Deposito di sostanze nutrienti e di prodotti di rifiuto (gomma, oppio, aroma dell’aglio..) Compartimento di degradazione Regolazione del pH cellulare (aumento di H+ nel citosol induce aumento del trasporto di H+ nel vacuolo) Regolazione osmotica con mantenimento del turgore (pressione osmotica che spinge verso l’esterno la parete cellulare ed avita alla pianta di avvizzire) mediante demolizione e risintesi controllate di polimeri (aumento pressione osmotica) Grosso aumento del volume cellulare senza aumento del volume del citosol (mezzo economico per aumentare dimensioni cellulari) --> vacuoli con funzioni diverse sono presenti nella stessa cellula
 Compartimento di degradazione. Regolazione del pH cellulare (aumento di H+ nel citosol induce aumento del trasporto di H+ nel vacuolo) Regolazione osmotica con mantenimento del turgore (pressione osmotica che spinge verso l’esterno la parete cellulare ed avita alla pianta di avvizzire) mediante demolizione e risintesi controllate di polimeri (aumento pressione osmotica) Grosso aumento del volume cellulare senza aumento del volume del citosol (mezzo economico per aumentare dimensioni cellulari) --> vacuoli con funzioni diverse sono presenti nella stessa cellula.")
15
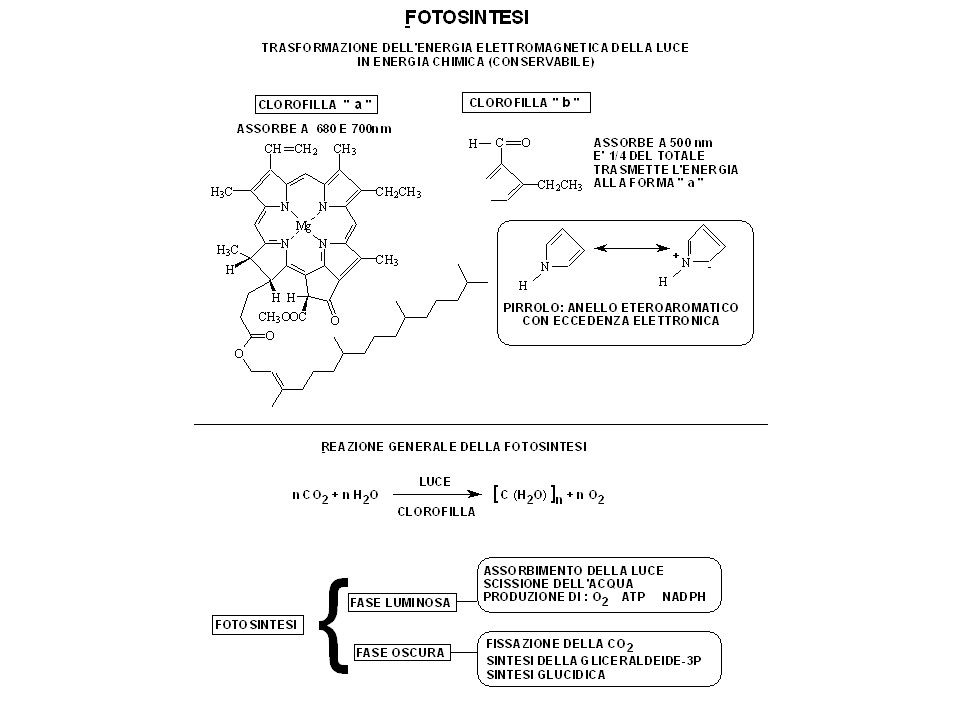
IL CLOROPLASTO -Tutti gli animali e la maggior parte dei microrganismi: continua assunzione di composti organici dal loro ambiente scheletri di carbonio per le biosintesi e l’energia metabolica per i processi cellulari - Sulla Terra primitiva: composti chimici prodotti da processi geochimici, ma esauriti dai primi organismi - Dal loro esaurimento (miliardi di anni fa) materiali organici necessari per le cellule viventi viene prodotto dagli organismi fotosintetici, tra cui molti tipi di batteri (cianobatteri sono quelli più evoluti) nH2O + nCO (CH2O)n + nO2 - Nei vegetali, sviluppatisi più tardi, la fotosintesi avviene in un organello specializzato: il cloroplasto luce
 materiali organici necessari per le cellule viventi viene prodotto dagli organismi fotosintetici, tra cui molti tipi di batteri (cianobatteri sono quelli più evoluti) nH2O + nCO2 (CH2O)n + nO2. - Nei vegetali, sviluppatisi più tardi, la fotosintesi avviene in un organello specializzato: il cloroplasto. luce.")
16
- Nei cloroplasti fotosintesi durante le ore di luce diurna.
prodotti: 1. utilizzati direttamente dalle cellule fotosintetiche per la biosintesi 2. convertiti in uno zucchero a basso P.M. (in genere saccarosio) esportato per le necessità metaboliche 3. depositati come materiali osmoticamente inerti (amido) che fungono da fonte di zuccheri per usi futuri - Prove biochimiche: cloroplasti discendono da batteri fotosintetici produttori di O2 endocitati e capaci di vivere in simbiosi con cellule eucariotiche primitive - Cloroplasti: membri della famiglia dei plastidi
 esportato per le necessità metaboliche 3. depositati come materiali osmoticamente inerti (amido) che fungono da fonte di zuccheri per usi futuri. - Prove biochimiche: cloroplasti discendono da batteri fotosintetici produttori di O2 endocitati e capaci di vivere in simbiosi con cellule eucariotiche primitive. - Cloroplasti: membri della famiglia dei plastidi.")
17
I PLASTIDI Plastidi sono presenti in tutte le cellule vegetali, hanno caratteristiche peculiari ma anche caratteristiche comuni: - copie multiple del piccolo genoma - involucro di 2 membrane concentriche Originano come proplastidi; evolvono a seconda delle necessità della cellula differenziata CROMOPLASTI: Con pigmenti colorati AMILOPLASTI: Producono amido e lo immagazzinano nello stroma (l’amido è osmoticamente inattivo) PROTEINOPLASTI: Producono proteine (in forma amorfa o cristallina) e le immagazzinano in sacche delimitate da membrane = “granuli di aleurone” ELAIOPLASTI (“OLEOSOMI”): Producono sostanze grasse CLOROPLASTI: Effettuano la fotosintesi, normalmente sono per cellula, contengono clorofilla e carotenoidi. Il numero per cellula può variare: duplicazione, degenerazione.
 PROTEINOPLASTI: Producono proteine (in forma amorfa o cristallina) e le immagazzinano in sacche delimitate da membrane = granuli di aleurone ELAIOPLASTI ( OLEOSOMI ): Producono sostanze grasse. CLOROPLASTI: Effettuano la fotosintesi, normalmente sono per cellula, contengono clorofilla e carotenoidi. Il numero per cellula può variare: duplicazione, degenerazione.")
18
STRUTTURA DEL CLOROPLASTO
Simili a mitocondri ma con un compartimento in più. - 1. membrana esterna molto permeabile 2. membrana interna molto meno permeabile con proteine trasportatrici 3.stretto spazio intermembrana - Membrana interna circonda ampio spazio = stroma (analogo della matrice mitocondriale), con enzimi, ribosomi, RNA e DNA - A differenza dei mitocondri, la membrana interna dei cloroplasti non ripiegata in creste e non contiene una catena di trasporto degli elettroni. - La catena, il sistema fotosintetico che assorbe la luce e una ATP sintasi sono contenuti in una terza membrana che forma sacchi a disco appiattito = tilacoidi
, con enzimi, ribosomi, RNA e DNA. - A differenza dei mitocondri, la membrana interna dei cloroplasti non ripiegata in creste e non contiene una catena di trasporto degli elettroni. - La catena, il sistema fotosintetico che assorbe la luce e una ATP sintasi sono contenuti in una terza membrana che forma sacchi a disco appiattito = tilacoidi.")
19
- lumi dei tilacoidi sono connessi spazio tilacoide, delimitato da membrana tilacoide
- “testa” dell’ATP sintasi sporge dalla membrana tilacoide nello stroma, mentre nel mitocondrio sporge dalla membrana mitocondriale interna nella matrice

20
IL GENOMA DEL CLOROPLASTO
- Non tutte le proteine del cloroplasto, similmente a quanto avviene nel mitocondrio, sono codificate da DNA nucleare e importate nell’organello dal citosol. Alcune codificate dal DNA del cloroplasto e sintetizzate sui suoi ribosomi. - traffico citosol - cloroplasto (e mitocondrio) è unidirezionale - Il trasporto delle proteine nel cloroplasto: avviene dopo la traduzione richiede energia idrolisi di ATP per l’importazione di proteine attraverso involucro esterno (membrana doppia) utilizza peptidi segnale anfipatici che vengono rimossi dopo l’uso sfrutta il gradiente elettrochimico per il passaggio attraverso la membrana tilacoide
 è unidirezionale. - Il trasporto delle proteine nel cloroplasto: avviene dopo la traduzione. richiede energia. idrolisi di ATP per l’importazione di proteine attraverso involucro esterno (membrana doppia) utilizza peptidi segnale anfipatici che vengono rimossi dopo l’uso. sfrutta il gradiente elettrochimico per il passaggio attraverso la membrana tilacoide.")
21
- molecole di DNA del cloroplasto (e del mitocondrio) sono relativamente piccole, semplici e circolari. Geni dei cloroplasti, 4 tipi di processi: - trascrizione - traduzione - fotosintesi - biosintesi di piccole molecole (aa., acidi grassi e pigmenti) - copie multiple della molecola di DNA distribuite in molti gruppi nella matrice del mitocondrio e nello stroma del cloroplasto - struttura del genoma assomiglia a quello batterico anziché alla cromatina degli eucarioti: no istoni - Il macchinario di sintesi proteica dei cloroplasti assomiglia molto a quello batterico: Ribosomi del cloroplasto in grado di usare tRNA batterici per la sintesi proteica Sintesi proteica inizia con N-formilmetionina, come nei batteri
 - copie multiple della molecola di DNA distribuite in molti gruppi nella matrice del mitocondrio e nello stroma del cloroplasto. - struttura del genoma assomiglia a quello batterico anziché alla cromatina degli eucarioti: no istoni. - Il macchinario di sintesi proteica dei cloroplasti assomiglia molto a quello batterico: Ribosomi del cloroplasto in grado di usare tRNA batterici per la sintesi proteica. Sintesi proteica inizia con N-formilmetionina, come nei batteri.")
23
Nella fotosintesi processo di conversione energia inizia quando una molecola di clorofilla è eccitata da un quanto di luce (fotone) e un e- passa da un orbitale ad uno ad energia più alta Clorofilla eccitata è instabile tende a stato originale in 3 modi: 1. convertendo energia extra in calore (clorofilla in soluzione) 2.trasferendo energia a molecola adiacente di clorofilla = trasferimento di energia di risonanza, non viene trasferito e- 3.trasferendo e- ad alta energia a accettore di elettroni e ritornando nello stato originale prendendo e- a bassa energia da donatore di elettroni 2. e 3. sono sfruttati nella fotosintesi
 e un e- passa da un orbitale ad uno ad energia più alta. Clorofilla eccitata è instabile tende a stato originale in 3 modi: 1. convertendo energia extra in calore (clorofilla in soluzione) 2.trasferendo energia a molecola adiacente di clorofilla = trasferimento di energia di risonanza, non viene trasferito e- 3.trasferendo e- ad alta energia a accettore di elettroni e ritornando nello stato originale prendendo e- a bassa energia da donatore di elettroni. 2. e 3. sono sfruttati nella fotosintesi.")
24
IL FOTOSISTEMA Catalizzano conversione dell’energia della luce catturata dalla clorofilla in forme utili. Energia della luce catturata nelle molecole di clorofilla eccitate forme utili di energia Localizzato nella membrana tilacoide Costituito da: 1. complesso antenna: proteine di membrana e numerose molecole di clorofilla che catturano l’energia luminosa e quantità variabili di pigmenti accessori = carotenoidi 2. centro di reazione fotochimica: complesso proteina-pigmento transmembrana: coppia speciale di molecole di clorofilla = trappola irreversibile per i quanti di eccitazione. Elettrone eccitato passa subito a catena di accettori di elettroni, ambiente molto più stabile

25
clorofilla carotene A seconda della pianta, quantità variabili di pigmenti accessori, i carotenoidi, che aiutano a raccogliere la luce con altre l, si trovano in ciascun complesso antenna

26
IL FOTOSISTEMA

27
REAZIONI ALLA LUCE Fotosintesi piante e cianobatteri produce ATP e NADPH mediante fotofosforilazione non ciclica: 2 fotosistemi in serie per aumentare energia di un e- Ogni e- che entra nel PS I è stato eccitato ad un livello E molto alto e può passare al centro Fe-S della ferredossina (Fd) e quindi al NADPH
 e quindi al NADPH.")
28
SCHEMA “Z”

29
I sistemi delle reazioni alla luce sono inseriti nella membrana tilacoide

30
In fotofosforilazione non ciclica poco più di una molecola di ATP per ogni coppia di e- che passa da H2O a NADP+ per produrre NADPH ma 1,5 molecole di ATP per NADPH sono necessarie per la fissazione del carbonio per produrre ATP extra: cloroplasti di alcune piante attuano fotofosforilazione ciclica fotosistema I trasferisce e- ad alta energia a complesso dei citocromi b6-f invece che a NADP+ reduttasi, così e- passano di nuovo a PSI a bassa energia e altri H+ sono pompati nello spazio tilacoide risultato netto: H+ pompato nel lume del tilacoide aumenta il gradiente elettrochimico sintesi ATP FOTOFOSFORILAZIONE CICLICA COINVOLGE SOLO IL FOTOSISTEMA I E PRODUCE ATP SENZA LA FORMAZIONE DI NADPH E O2

31
REAZIONI AL BUIO ATP e NADPH: fonte di energia e potere riducente per la conversione di CO2 (C inorganico) in carboidrati (C organico) Iniziano nello stroma del cloroplasto e continuano nel citosol della cellula vegetale Reazione centrale della fissazione del carbonio Reazione catalizzata da ribulosio bisfosfato carbossilasi: grosso enzima. Reazione avviene nello stroma, procede lentamente, sono perciò necessarie molte molecole di enzima Tale enzima costituisce spesso più del 50% delle pt totali del cloroplasto Equazione netta: 3 CO2 + 9 ATP + 6 NADPH + acqua gliceraldeide 3-fosfato + 8Pi + 9ADP + NADP+

32
Ciclo di Calvin: guadagno netto di 3C
- 3 ATP e 2 NADPH utilizzati per ogni molecola di CO2

33
maggior parte GAP che rimane nel cloroplasto viene trasformata in AMIDO = riserva di carboidrati
saccarosio = forma principale nella quale lo zucchero è trasportato fra le cellule vegetali fogliefasci vascolari

34
PERO’ CO2 RIBULOSIO 1,5-DIFOSFATO (C 5) 2 ( 3-FOSFOGLICERATO) 2 (C3) RIBULOSIO 1,5-DIFOSFATO CARBOSSILASI PM: circa (NB: H2O 18). Composto da 16 subunità: 8 prodotte dalla cellula e 8 dal cloroplasto. Probabilmente è uno degli enzimi più antichi del Pianeta. E’ il più abbondante sul pianeta. E’ lento e fissa anche l’ossigeno FOTORESPIRAZIONE: utilizza O2 e produce CO2 senza produzione di depositi utili di energia FOTORESPIRAZIONE si forma fosfoglicolato e si perde circa 1/3 della CO2 precedentemente fissata.
. Composto da 16 subunità: 8 prodotte dalla cellula e 8 dal cloroplasto. Probabilmente è uno degli enzimi più antichi del Pianeta. E’ il più abbondante sul pianeta. E’ lento e fissa anche l’ossigeno FOTORESPIRAZIONE: utilizza O2 e produce CO2 senza produzione di depositi utili di energia. FOTORESPIRAZIONE si forma fosfoglicolato e si perde circa 1/3 della CO2 precedentemente fissata.")
35
Piante C3: effettuano il ciclo C3
Fotorespirazione problema serio per piante in condizioni calde e secche, in cui chiudono gli stomi (pori che scambiano gas nelle foglie) per evitare eccessiva perdita di acqua caduta livelli CO2 che favorisce fotorespirazione Per evitare la FOTORESPIRAZIONE le piante C3 si sono evolute in C4 e successivamente in piante CAM. Piante C3: effettuano il ciclo C3 Piante C4: effettuano il ciclo C4 e il ciclo C3 di giorno Piante CAM: effettuano il ciclo C4 di notte e il ciclo C3 di giorno a stomi chiusi Cactus spinoso (Opuntia sp.) è una pianta CAM
 per evitare eccessiva perdita di acqua caduta livelli CO2 che favorisce fotorespirazione. Per evitare la FOTORESPIRAZIONE le piante C3 si sono evolute in C4 e successivamente in piante CAM. Piante C3: effettuano il ciclo C3. Piante C4: effettuano il ciclo C4 e il ciclo C3 di giorno. Piante CAM: effettuano il ciclo C4 di notte e il ciclo C3 di giorno a stomi chiusi. Cactus spinoso (Opuntia sp.) è una pianta CAM.")
36
Piante C4: ciclo di Calvin confinato nelle cellule della guaina del fascio, relativamente isolate dalla CO2 e dall’O2 dell’atmosfera. Utilizzano il ciclo di Hatch-Slack per raccogliere CO2 nelle cellule del mesofillo e per concentrarla nelle cellule della guaina del fascio, che circondano i fasci vascolari della foglia. Tale disposizione concentrica = anatomia Kranz, essenziale per l’efficienza fotosintetica delle piante C4. Piante C3: ciclo di Calvin nelle cellule del mesofillo

37
Ciclo di Hatch-Slack Nelle cellule del mesofillo l’enzima fosfoenolpiruvato carbossilasi (PEP carbossilasi) combina CO2 con una molecola a 3C, originando composto a 4C, il malato, che diffonde nella c. della guaina del fascio attraverso i plasmodesmi Nella cellule della guaina il malato viene demolito per rilasciare CO2 e piruvato, riportato nel mesofillo
 combina CO2 con una molecola a 3C, originando composto a 4C, il malato, che diffonde nella c. della guaina del fascio attraverso i plasmodesmi. Nella cellule della guaina il malato viene demolito per rilasciare CO2 e piruvato, riportato nel mesofillo.")
38
Il pompaggio della CO2 costa energia.
Ma negli ambienti caldi e secchi questo costo può essere inferiore all’energia persa dalla fotorespirazione nelle piante C3 piante C4 hanno vantaggio potenziale

39
SI NO Fonte di energia Fissazione CO2 Fissazione N2 Esempio LUCE
FOTOAUTOTROFI LUCE SI cianobatteri NO piante CHEMIOAUTOTROFI COMPOSTI INORGANICI solfobatteri FOTOETEROTROFI solo per produrre ATP alcuni procarioti CHEMIOETEROTROFI COMPOSTI ORGANICI funghi e animali AUTOTROFI ETEROTROFI

40
FISSAZIONE DELL’AZOTO
Tra tutti gli elementi minerali, Azoto limita maggiormente la crescita di piante e la resa dei raccolti PARADOSSO: piante soffrono carenza azoto ma atmosfera circa 80% azoto Azoto atmosferico = N2 non è direttamente utilizzabile dalle piante Deve essere convertito in NH3, con reazione che richiede grande dispendio energetico perché il triplo legame della molecola di N2 conferisce notevole stabilità

41
Batteri azotofissatori:
1. procarioti che convertono N2 nella sua forma utile mediante il processo di fissazione dell’azoto 2. possono vivere liberi nel terreno ma rendono disponibile l’azoto fissato solo dopo la loro morte 3. vivono in simbiosi con le radici di certe piante: Rhizobium vive in noduli situati a livello radici delle leguminose (soia, piselli, fagioli, trifoglio, erba medica) cede alla pianta sino al 90% dell’azoto fissato e secerne alcuni aa nel terreno per rendere azoto immediatamente disponibile a altri organismi 4. Rhizobium assume la forma di batteroide contenuto in vacuoli di cellule radicali 5. la simbiosi è di tipo mutualistico 6. cianobatteri si associano con la componente fungina dei licheni o con le felci 7. attinomiceti ( batteri filamentosi) con le radici dell’ontano I grandi noduli rotondi si sviluppano in corrispondenza di un esteso sistema radicale e contengono batteri azotofissatori
 cede alla pianta sino al 90% dell’azoto fissato e secerne alcuni aa nel terreno per rendere azoto immediatamente disponibile a altri organismi. 4. Rhizobium assume la forma di batteroide contenuto in vacuoli di cellule radicali. 5. la simbiosi è di tipo mutualistico. 6. cianobatteri si associano con la componente fungina dei licheni o con le felci. 7. attinomiceti ( batteri filamentosi) con le radici dell’ontano. I grandi noduli rotondi si sviluppano in corrispondenza di un esteso sistema radicale e contengono batteri azotofissatori.")
42
Azoto viene fissato ad opera di nitrogenasi
La reazione richiede grandi quantità di energia: per ogni molecola di NH3 16 di ATP (a seconda della specie batterica fornite da fotosintesi o da processi respiratori) Nitrogenasi è fortemente inibita da O2 Ambiente interno dei noduli radicali in cui si trova Rhizobium è privo di ossigeno Grosso ostacolo nell’ottenimento di piante ricombinanti ottenute per inserimento nel genoma del gene per la nitrogenasi: esclusione dell’ossigeno
 Nitrogenasi è fortemente inibita da O2. Ambiente interno dei noduli radicali in cui si trova Rhizobium è privo di ossigeno. Grosso ostacolo nell’ottenimento di piante ricombinanti ottenute per inserimento nel genoma del gene per la nitrogenasi: esclusione dell’ossigeno.")
43
NH3 fissato è potenzialmente tossico per le piante
Processo di nitrificazione converte ammoniaca in nitrati = NO3 - I nitrati passano alla pianta che poi li riduce a nitriti e nuovamente ad ammoniaca

Presentazioni simili



















>")