Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

2
Neuropeptidi orexigenici
Modello temporaneo (molto) semplificato dei circuiti del SNC implicati nell’anoressia e nella cachessia del portatore di tumore - Cortisolo + Neuropeptidi orexigenici NPY - Recettore per la melanocortina, MCR Leptina e insulina - + + Neuropeptidi anorexigenici CRH Lipolisi norepinefrina, dopamina, serotonina Controllo dell’appetito Termogenesi e metabolismo basale
 semplificato dei circuiti del SNC implicati nell’anoressia e nella cachessia del portatore di tumore. - Cortisolo. + Neuropeptidi orexigenici. NPY. - Recettore per la melanocortina, MCR. Leptina. e insulina Neuropeptidi anorexigenici. CRH. Lipolisi. norepinefrina, dopamina, serotonina. Controllo dell’appetito. Termogenesi e metabolismo basale ")
3
Neuropeptidi orexigenici
Modello temporaneo (molto) semplificato dei circuiti del SNC implicati nell’anoressia e nella cachessia del portatore di tumore Tumore IL-1, TNFa, IFNg - + Neuropeptidi orexigenici NPY - - Recettore per la melanocortina, MCR Leptina e insulina - + + Neuropeptidi anorexigenici CRH + Tumore Lipolisi IL-1, TNFa, IFNg Tumore + norepinefrina, dopamina, serotonina Anoressia Termogenesi e metabolismo basale
 semplificato dei circuiti del SNC implicati nell’anoressia e nella cachessia del portatore di tumore. Tumore. IL-1, TNFa, IFNg. - + Neuropeptidi orexigenici. NPY. - - Recettore per la melanocortina, MCR. Leptina. e insulina Neuropeptidi anorexigenici. CRH. + Tumore. Lipolisi. IL-1, TNFa, IFNg. Tumore. + norepinefrina, dopamina, serotonina. Anoressia. Termogenesi e metabolismo basale ")
4
Regulation of NPY production in the hypothalamic arcuate nucleus
Regulation of NPY production in the hypothalamic arcuate nucleus. Glucocorticoids stimulate hypothalamic production of neuropeptide Y (NPY), leading to increased food intake and reduced energy expenditure. Leptin, which is produced by adipose tissue, blocks NPY production, as does insulin, which is produced by the pancreas, and increased NPY decreases leptin and insulin production.
, leading to increased food intake and reduced energy expenditure. Leptin, which is produced by adipose tissue, blocks NPY production, as does insulin, which is produced by the pancreas, and increased NPY decreases leptin and insulin production.")
5
Digiuno post-prandiale (5-6 h)
Aminoacidi ACIDI GRASSI GLUCOSIO _ catecolamine insulina

6
Digiuno prolungato (1-7 giorni)
Proteine GLUCOSIO Aminoacidi CORPI CHETONICI + glucagone ACIDI GRASSI

7
Digiuno protratto N ureico nelle urine (da 12 a 2 g/24 h) Tessuti +
GLUCOSIO Aminoacidi CORPI CHETONICI + glucagone

8
Controllo dell’ossidazione degli acidi grassi
* *insulina glucagone Chetogenesi in absence of oxalacetate Keton bodies

9
Nel soggetto digiunante
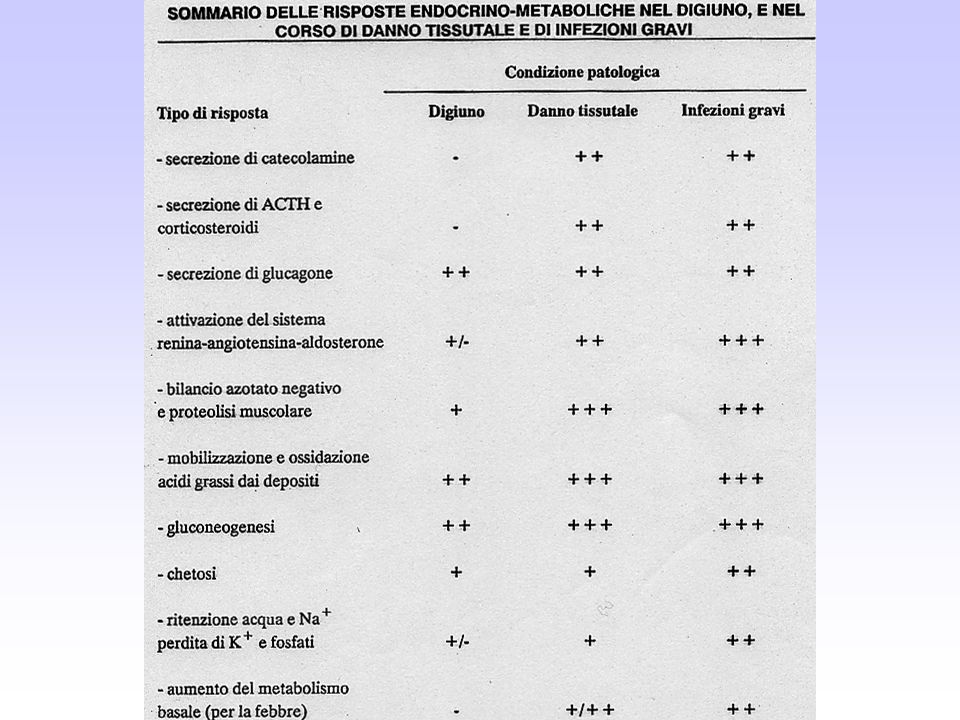
CARATTERISTICHE ENDOCRINO-METABOLICHE DEL SOGGETTO DIGIUNANTE E DEL PORTATORE DI TUMORE Nel soggetto digiunante Aumento della lipolisi Blocco della sintesi de novo degli acidi grassi Inibizione della lipogenesi Inibizione della lipasi lipoproteica (iperlipemia) Aumento della proteolisi muscolare Aumento dell’azoto ureico nelle urine Riduzione della sintesi proteica nei muscoli scheletrici Aumento della sintesi epatica delle proteine della fase acuta Aumento della gluconeogenesi dagli aminoacidi, dal glicerolo, dall’acido lattico (prodotto dal tumore e dai tessuti dell’ospite) Resistenza all’insulina Iperinsulinemia Intolleranza al glucosio Aumento del metabolismo basale Insulinemia fortemente aumentata Cortisolo nel siero aumentato (digiuno prolungato) Riduzione del metabolismo basale Insulinemia ridotta Cortisolo nel siero non modificato
 Aumento della proteolisi muscolare. Aumento dell’azoto ureico nelle urine. Riduzione della sintesi proteica nei muscoli scheletrici. Aumento della sintesi epatica delle proteine della fase acuta. Aumento della gluconeogenesi dagli aminoacidi, dal glicerolo, dall’acido lattico (prodotto dal tumore e dai tessuti dell’ospite) Resistenza all’insulina. Iperinsulinemia. Intolleranza al glucosio. Aumento del metabolismo basale. Insulinemia fortemente aumentata. Cortisolo nel siero aumentato. (digiuno prolungato) Riduzione del metabolismo basale. Insulinemia ridotta. Cortisolo nel siero non modificato.")
10
- - Effetto del tumore sulla sintesi e sull’utilizzazione
(TNFa) ACIDI GRASSI TRIGLICERIDI - (TNFa) Tumore Effetto del tumore sulla sintesi e sull’utilizzazione degli acidi grassi dopo il pasto
 ACIDI GRASSI. TRIGLICERIDI. - (TNFa) Tumore. Effetto del tumore sulla sintesi e sull’utilizzazione. degli acidi grassi dopo il pasto.")
11
Caratteristiche metaboliche del portatore di tumore
N ureico nelle urine Sintesi proteica PROTEINE Corpi chetonici tessuti, tumore lattato PIF TNFa Aminoacidi Aminoacidi TNF Corpi chetonici Sintesi delle proteine della fase acuta Caratteristiche metaboliche del portatore di tumore

12
Breve storia della scoperta del ruolo delle citochine infiammatorie
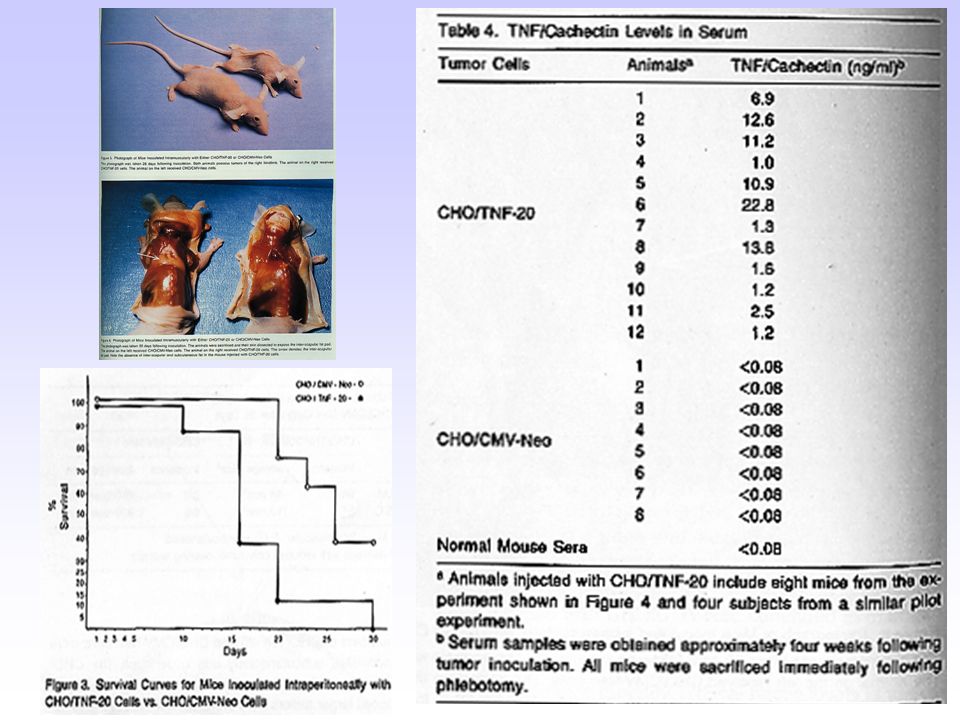
nella cachessia neoplastica Cerami e coll. negli anni ‘70 osservano in conigli affetti da tripanosomiasi una notevole riduzione della massa corporea proteica e lipidica associata a una marcata iperlipidemia come risultato di una ridotta attività lipoprotein- lipasica; Kawakami e Cerami nel 1981 dimostrano che il trattamento con endotossine batteriche induce nel topo la comparsa di un fattore serico capace di sopprimere l’attività lipoprotein-lipasica; Beutler e Cerami nel 1985 dimostrano che il fattore inibitorio dell’attività lipoprotein- lipasica corrisponde a una molecola polipeptidica, di origine macrofagica, a struttura ben definita che viene denominata cachettina; Beutler e Cerami nel 1986 dimostrano che la sequenza aminoacidica della cachettina è omologa a quella del fattore di necrosi tumorale (TNF); Successivamente l’analisi genetica ha confermato che la cachettina e il TNF sono molecole identiche.
; Successivamente l’analisi genetica ha confermato che la cachettina e il TNF sono molecole identiche.")
14
Catabolic mediators in cancer.
Both tumor-derived and humoural (cytokines) factors are involved in mediating anorexia and metabolic changes, characteristic of the cachectic state.
 factors are involved in mediating anorexia and metabolic changes, characteristic of the cachectic state.")
15
PIRUVATO CARBOSSILASI
ATP GLUCAGONE + ESOCHINASI INSULINA + GLUCAGONE_--_ GLUCOSIO 6-FOSFATASI GLUCOCHINASI ADP ATP GLUCAGONE + FRUTTOSIO 1,6-DIFOSFATASI INSULINA + GLUCAGONE_--_ FOSFOFRUTTOCHINASI ADP ADP ADP GLICEROCHINASI ATP ATP GLICEROLO ADP INSULINA + GLUCAGONE_--_ PIRUVATO CHINASI ATP GDP LATTATO GLUCAGONE + FOSFOENOLPIRUVATO CARBOSSICHINASI GTP ATP PIRUVATO CARBOSSILASI ADP PROPIONATO

16
MF Cori cycle with sources of gluconeogenic substrates. Tumours produce factors such as lipid-mobilizing factor (LMF), which induces breakdown of adipose tissue into fatty acids, and proteolysis-inducing factor (PIF), which induces protein degradation (amino acids) in skeletal muscle. Tumour necrosis factor (TNF)-a also contributes to these processes. These are important gluconeogenic substrates that can be used in acute-phase protein (APP) synthesis by the liver. Tumours convert glucose to lactate, which is transferred to the liver, where it is converted back into glucose. This cycle uses a large amount of energy, and might contribute to cachexia.
, which induces breakdown of adipose tissue into fatty acids, and proteolysis-inducing factor (PIF), which induces protein degradation (amino acids) in skeletal muscle. Tumour necrosis factor (TNF)-a also contributes to these processes. These are important gluconeogenic substrates that can be used in acute-phase protein (APP) synthesis by the liver. Tumours convert glucose to lactate, which is transferred to the liver, where it is converted back into glucose. This cycle uses a large amount of energy, and might contribute to cachexia.")
18
Synthesis and degradation of proteins in skeletal muscle
Synthesis and degradation of proteins in skeletal muscle. Protein levels in muscle are determined by the amount of dietary intake of protein and levels of protein synthesis. Decreases in plasma insulin concentrations or insulin sensitivity of skeletal muscle can activate three main proteolytic pathways that underlie protein catabolism in skeletal muscle. These are the lysosomal system, which proteolyses extracellular proteins and cell-surface receptors; the cytosolic calcium-activated system, which involves calpains I and II and is involved in tissue injury, necrosis and autolysis, and the ATP-ubiquitin-dependent proteolytic pathway. This proteolysis leads to hepatic production of acute-phase protein (APP), which can limit the availability of certain amino acids for protein synthesis in skeletal muscle. Protein deamination also leads to nitrogen excretion, producing a negative nitrogen balance, and glucose production, which increases muscle activity.
, which can limit the availability of certain amino acids for protein synthesis in skeletal muscle. Protein deamination also leads to nitrogen excretion, producing a negative nitrogen balance, and glucose production, which increases muscle activity.")
19
Ubiquitin-proteasome pathway Ubiquitin (Ub)
")
20
The Ubiquitin–Proteasome Pathway of Proteolysis.

22
Indicazioni sulla partecipazione del sistema ubiquitinico nella proteolisi dei muscoli scheletrici nella cachessia neoplastica Aumento dell’mRNA dell’ubiquitina del muscolo retto-addominale di portatore di cancro gastrico Aumento delle subunità alfa e beta del proteosoma nei muscoli del portatore di tumore Aumentata espressione dell’enzima E2 che coniuga l’ubiquitina alle proteine Aumentata espressione dell’enzima E3 legante la proteina

23
Muscle breakdown. Signaling pathways that regulate protein homeostasis in skeletal muscle. Cytokines such as TNF-a together with IFN-g activate the transcription factor NF-kB. This leads to decreased expression of MyoD, a transcription factor that may be important for replenishing wasted muscle. Activated NF-kB also acts as a repressor of proteasome subunit expression and hence suppresses protein degradation, an activity that is antagonized by glucocorticoids. (The proteasome is a multisubunit complex involved in the breakdown of ubiquitinated proteins.) Tumor factors such as PIF increase production of proteasome subunits through the intermediary 15-HETE. It is not known whether this is a direct or indirect effect (dashed arrows). Eicosapentaenoic acid (EPA) inhibits 15-HETE production in response to PIF and prevents muscle wasting in cancer patients.
 Tumor factors such as PIF increase production of proteasome subunits through the intermediary 15-HETE. It is not known whether this is a direct or indirect effect (dashed arrows). Eicosapentaenoic acid (EPA) inhibits 15-HETE production in response to PIF and prevents muscle wasting in cancer patients..")
24
Interactions between pro-inflammatory cytokines and PIF

25
The Importance of Myosin in Cachexia
The Importance of Myosin in Cachexia. Soluble factors released from tumors or immune effector cells and implicated in cachexia can lead to a specific decrease in the levels of the myosin heavy chain, a muscle contractile protein. The data provide support for the existence of two pathways. In Panel A, the combination of tumor necrosis factor a (TNF-a) and interferon- g (IFN-g) results in the suppression of the nuclear transcription factor MyoD and, hence, a decrease in the transcription of the myosin heavy chain; a deficit in the cellular pool of myosin heavy chain results in cachexia. Cytokines such as interleukin-6 increase the production of ubiquitin and E3 ubiquitin ligase proteins. In Panel B, stimulation of the ubiquitin ligase–dependent proteasome pathway leads to increased and preferential ubiquination of the myosin heavy chain, causing the dissociation of myosin from the contractile apparatus and its subsequent degradation into peptides by the proteasomes. The loss of functional contractile units, probably combined with the selective loss of other specific proteins, leads to muscle atrophy and wasting.
 and interferon- g (IFN-g) results in the suppression of the nuclear transcription factor MyoD and, hence, a decrease in the transcription of the myosin heavy chain; a deficit in the cellular pool of myosin heavy chain results in cachexia. Cytokines such as interleukin-6 increase the production of ubiquitin and E3 ubiquitin ligase proteins. In Panel B, stimulation of the ubiquitin ligase–dependent proteasome pathway leads to increased and preferential ubiquination of the myosin heavy chain, causing the dissociation of myosin from the contractile apparatus and its subsequent degradation into peptides by the proteasomes. The loss of functional contractile units, probably combined with the selective loss of other specific proteins, leads to muscle atrophy and wasting.")
26
Proton transport by UCP1 across the inner mitochondrial membrane Normally, proton transport across the inner mitochondrial membrane is coupled to phosphorylation of ADP to generate ATP. An increase in membrane proton permeability that is not coupled to an energy-consuming system constitutes a proton leak. This leak decreases the coupling of respiration to ADP phosphorylation, and increases substrate oxidation and the dissipation of oxidation energy as heat. This process protects against hypothermia and regulates energy balance. Both animal and plant mitochondria contain a group of mitochondrial carrier proteins known as the uncoupling proteins (UCP). UCP1 is highly expressed by brown adipose tissue (BAT) — the main site for thermogenesis. The inner membrane of BAT mitochondria have a high permeability to protons, due to the abundance of UCP1, so ATP production is uncoupled, leading to heat production. Two mechanisms have been proposed by which UCP1 is able to transport protons. a | In the first model, UCP1 transports protons (H+) and fatty acids (RCOO-). The fatty acid provides a free carboxyl group that makes proton transport possible. The proton is then liberated from the fatty acid after it has crossed the membrane. b | In the second model, the protonated form of the fatty acid diffuses freely across the membrane, and UCP1 transports the anionic form of the fatty acid (RCOO-) back across to the other side.
. UCP1 is highly expressed by brown adipose tissue (BAT) — the main site for thermogenesis. The inner membrane of BAT mitochondria have a high permeability to protons, due to the abundance of UCP1, so ATP production is uncoupled, leading to heat production. Two mechanisms have been proposed by which UCP1 is able to transport protons. a | In the first model, UCP1 transports protons (H+) and fatty acids (RCOO-). The fatty acid provides a free carboxyl group that makes proton transport possible. The proton is then liberated from the fatty acid after it has crossed the membrane. b | In the second model, the protonated form of the fatty acid diffuses freely across the membrane, and UCP1 transports the anionic form of the fatty acid (RCOO-) back across to the other side..")
Presentazioni simili















 A.Cristofoli (laureando.>")

>")

>")
>")

