Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

4
Aumento delle dimensioni degli organismi e della specializzazione cellulare Perdita di contatto diretto fra cellule ed ambiente esterno per rifornimento di materiali Sistema interno di trasporto

5
La circolazione del sangue fornisce un sistema di trasporto e di comunicazione fra le cellule, contribuendo a mantenere un ambiente interno relativamente stabile ed ottimale per l’attività cellulare. A rigore il sangue non è un liquido corporeo come le lacrime, la saliva e l’urina, ma piuttosto un tessuto connettivo specializzato formato da una soluzione acquosa (plasma) nella quale sono contenute sostanze varie: gas, elettroliti, metaboliti, cataboliti, vitamine, proteine e diverse popolazioni cellulari (parte corpuscolata)
 nella quale sono contenute sostanze varie: gas, elettroliti, metaboliti, cataboliti, vitamine, proteine e diverse popolazioni cellulari (parte corpuscolata).")
6
Il sangue costituisce circa il 6% del peso corporeo. Il plasma è circa 3 litri ed è formato al 90% da acqua, mentre della rimanente porzione il 70% sono proteine, il 20% molecole di piccole dimensioni e il 10% elettroliti La porzione corpuscolata è formata al 99% da eritrociti, l’1% sono leucociti e trombociti o piastrine. Ematocrito: volume percentuale della porzione corpuscolata rispetto al volume totale. Vale mediamente 42% nelle donne e 45% negli uomini. Per siero si intende invece il plasma privo dei fattori di coagulazione L’osmolarità del plasma risulta essere circa 280-300 mOsm/Kg.

7
Glucosioaa ac. grassi elettrolitiacqua Ossigeno Anidride carbonica Urea Acido urico Acqua in eccesso Ormoni Calore SANGUE

8
Le funzioni del sangue sono divisibili in due gruppi: 1. Funzioni di trasporto: respiratoria nutritiva depuratrice endocrina 2. Funzioni di regolazione dell’omeostasi corporea regolazione equilibrio acido-base difesa regolazione della temperatura regolazione equilibrio idrico-salino regolazione pressione osmotica regolazione pressione arteriosa emostasi

9
Cellule del sangue Fagocitosi di sostanze tossiche e microorganismi Trasporto di gas Produzione e rilascio di anticorpi Inoltre bisogna ricordare che nel sangue si trovano importanti sistemi tampone che aiutano a mantenere costante il valore del pH. Vedremo inoltre che le proteine del sangue hanno una funzione fondamentale nel mantenere una pressione osmotica che è fondamentale negli scambi transcapillari di acqua e soluti.

10
Volume del sangue (l): 7% ca del peso totale in Kg della massa corporea priva di grassi Volume del sangue (l): 7% ca del peso totale in Kg della massa corporea priva di grassi Peso specifico: 1.057-1.062 gr/ml Peso specifico: 1.057-1.062 gr/ml Concentrazione osmotica: 300mosm/l Concentrazione osmotica: 300mosm/l pH compreso fra 7.35 e 7.40 pH compreso fra 7.35 e 7.40
: 7% ca del peso totale in Kg della massa corporea priva di grassi Volume del sangue (l): 7% ca del peso totale in Kg della massa corporea priva di grassi Peso specifico: gr/ml Peso specifico: gr/ml Concentrazione osmotica: 300mosm/l Concentrazione osmotica: 300mosm/l pH compreso fra 7.35 e 7.40 pH compreso fra 7.35 e 7.40")
11
Costituente Concentrazione (mEq/l) Na + 135-145 (300mg/dl) K+K+K+K+ 3.5-5.0 (20mg/dl) Ca 2+ 2.2-2.5 (9-10mg/dl) Mg 2+ 1.5-2.0 H+H+H+H+ pH 7.35-7.45 Cl - 95-107 HCO 3 - 22-26 Lattato1.0-1.8 Solfato1.0 Fosfato2.0
 Na (300mg/dl) K+K+K+K (20mg/dl) Ca (9-10mg/dl) Mg H+H+H+H+ pH Cl HCO Lattato Solfato1.0 Fosfato2.0")
12
Proteine plasmatiche (6-8 gr /dl) 69kDa
 69kDa")
13
Riserva per la fornitura di aa Proteine plasmatiche Trasportatori di altre molecole (O 2, CO 2, lipidi, metalli, farmaci) Funzione tampone per mantenere il pH Proteine a funzione coagulante Funzione nella genesi di una pressione colloido-osmotica (25mmHg) Funzione ormonale
 Funzione tampone per mantenere il pH Proteine a funzione coagulante Funzione nella genesi di una pressione colloido-osmotica (25mmHg) Funzione ormonale")
14
PatologiaEnzima Infarto del miocardio Creatin fosfochinasi Transaminasi glutammico piruvica Transaminasi glutammico ossalacetica Affezioni epatiche Transaminasi glutammico piruvica Transaminasi glutammico ossalacetica Lattato deidrogenasi Alterazioni ossee Fosfatasi alcalina Tumori della prostata Fosfatasi acida Lesioni del pancreas Amilasi

15
Fra le proteine riveste particolare importanza l’albumina (4 gr/dl) che è sintetizzata a livello epatico e non diffonde attraverso l’epitelio vascolare. É la principale responsabile della pressione colloido-osmotica o oncotica che regola il passaggio di acqua e soluti fra letto capillare e interstizio.

16
Lipidi: colesterolo di cui il 30% in forma libera e il 70% in forma esterificata. Colesterolo, fosfolipidi e trigliceridi sono trasportati sotto forma di lipoproteine (chilomicroni, VLDL, LDL e HDL) Lipidi: colesterolo di cui il 30% in forma libera e il 70% in forma esterificata. Colesterolo, fosfolipidi e trigliceridi sono trasportati sotto forma di lipoproteine (chilomicroni, VLDL, LDL e HDL) Glicidi: principalmente glucosio (80-110mg/dl) Glicidi: principalmente glucosio (80-110mg/dl) Azoto: 10-25mg/dl sotto forma di urea, acido urico, creatinina, ammoniaca e aa Azoto: 10-25mg/dl sotto forma di urea, acido urico, creatinina, ammoniaca e aa
 Lipidi: colesterolo di cui il 30% in forma libera e il 70% in forma esterificata. Colesterolo, fosfolipidi e trigliceridi sono trasportati sotto forma di lipoproteine (chilomicroni, VLDL, LDL e HDL) Glicidi: principalmente glucosio (80-110mg/dl) Glicidi: principalmente glucosio (80-110mg/dl) Azoto: 10-25mg/dl sotto forma di urea, acido urico, creatinina, ammoniaca e aa Azoto: 10-25mg/dl sotto forma di urea, acido urico, creatinina, ammoniaca e aa.")
18
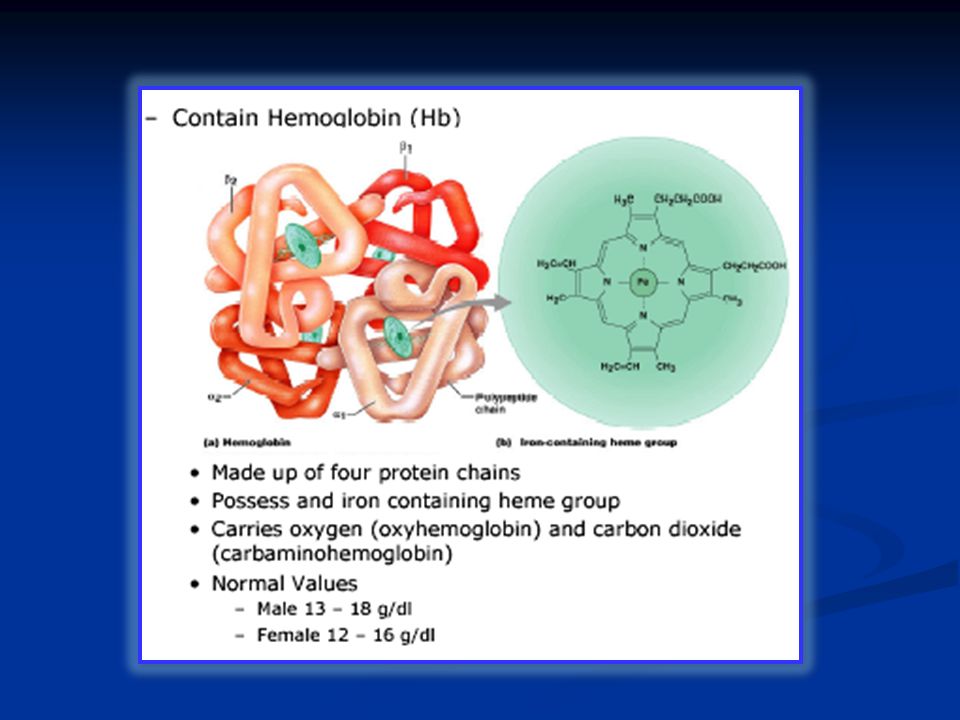
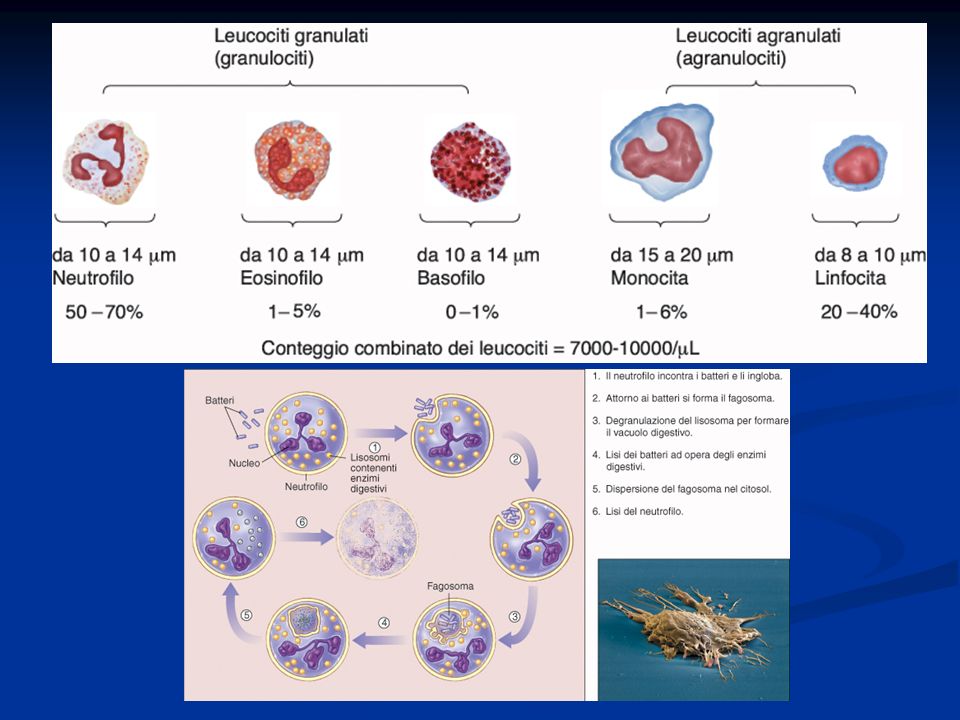
Le cellule del sangue comprendono: Eritrociti: Sono circa 5 milioni/mm 3 nell’uomo e 4.5 nella donna. Contengono circa 15 gr/dl di emoglobina. Sono circa 7μm di diametro. Hanno una vita media di 120 gg dopo i quali sono distrutti a livello splancnico. Leucociti: sono circa 4000-10.000/mm 3 e comprendono i granulociti (neutrofili 40-75%, basofili < 1%, eosinofili 1- 6%) e le cellule linfoidi con monociti (2-10%) e linfociti (20-45%) Piastrine: 150.000-400.000/mm 3 sono responsabili della coagulazione del sangue.
 e le cellule linfoidi con monociti (2-10%) e linfociti (20-45%) Piastrine: /mm 3 sono responsabili della coagulazione del sangue..")
21
L’eritropoiesi è la formazione dei globuli rossi da parte del midollo rosso delle ossa piatte (sterno, vertebre, coste, ossa della pelvi) a partire dalle cellule staminali totipotenti. Sono prodotti circa 230 miliardi di globuli rossi al giorno e altrettanti sono distrutti. Le sostanze necessarie per l’eritropoiesi sono molte, fra cui la vitamina B 12, ac. folico, ferro e proteine fra cui l’eritropoietina che ha funzione di controllo ed è prodotta in massima parte a livello renale. Controlla la quantità prodotta in condizioni patologiche (anemie, emorragia…) o parafisiologiche (alta montagna etc..)
 o parafisiologiche (alta montagna etc..).")
22
La distruzione dei globuli rossi comporta la riutilizzazione di molte parti dell’emoglobina, mentre altre sono escrete sotto forma di bilirubina.

28
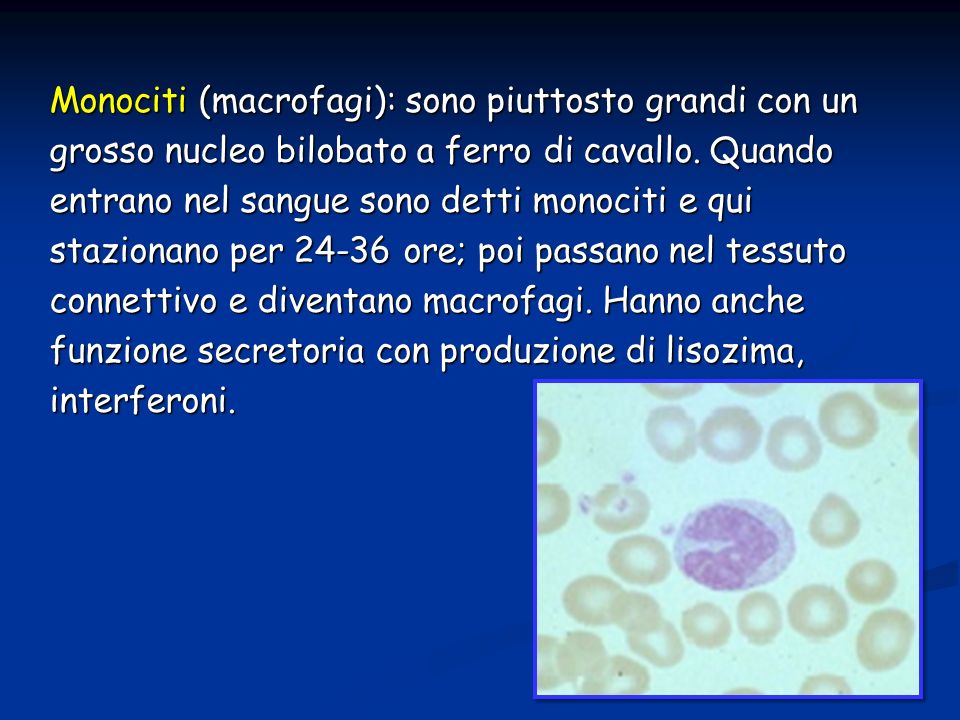
Monociti (macrofagi): sono piuttosto grandi con un grosso nucleo bilobato a ferro di cavallo. Quando entrano nel sangue sono detti monociti e qui stazionano per 24-36 ore; poi passano nel tessuto connettivo e diventano macrofagi. Hanno anche funzione secretoria con produzione di lisozima, interferoni.

29
Linfociti: comprendono generalmente il 25-30% delle cellule bianche del sangue. Sono cellule molto attive dal punto di vista immunitario e comprendono diverse classi, ciascuna con specifiche funzioni e proprietà. Gli organi linfoidi comprendono il timo, il midollo osseo, la milza, il linfonodi, le tonsille palatine, le placche di Peyer e il tessuto linfoide dei tratti respiratorio e digerente.

31
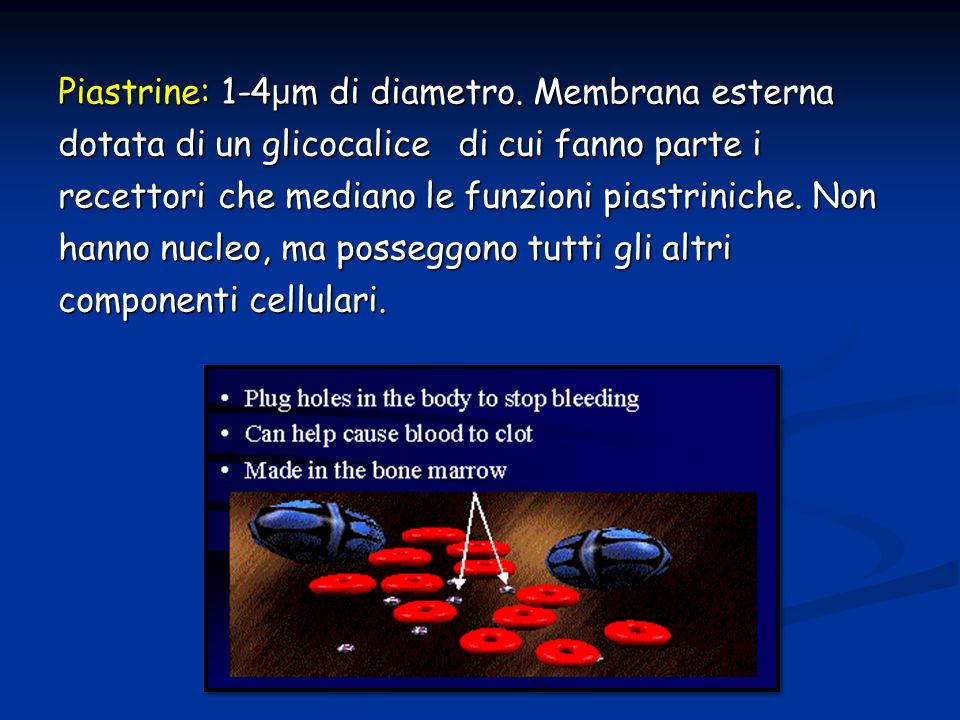
Piastrine: 1-4 μ m di diametro. Membrana esterna dotata di un glicocalice di cui fanno parte i recettori che mediano le funzioni piastriniche. Non hanno nucleo, ma posseggono tutti gli altri componenti cellulari.

35
Evoluzione del sistema circolatorio Un sistema circolatorio può essere aperto o chiuso. In un sistema aperto il sangue è pompato dal cuore attraverso i vasi ma poi lascia i vasi sanguigni ed entra in cavità (emocele) in un fluido detto emolinfa. Da questo liquido ossigeno e nutrienti entrano in contatto con le cellule. L’emolinfa scorre lentamente perché non vi è pressione sanguigna dopo che essa ha lasciato i vasi. L’animale deve muovere i muscoli per far muovere l’emolinfa in questi spazi. Gli Artropodi e la maggior parte dei molluschi (eccetto i cefalopodi) hanno un sistema aperto (sistema lacunare).
 in un fluido detto emolinfa. Da questo liquido ossigeno e nutrienti entrano in contatto con le cellule. L’emolinfa scorre lentamente perché non vi è pressione sanguigna dopo che essa ha lasciato i vasi. L’animale deve muovere i muscoli per far muovere l’emolinfa in questi spazi. Gli Artropodi e la maggior parte dei molluschi (eccetto i cefalopodi) hanno un sistema aperto (sistema lacunare)..")
36
In un sistema chiuso il sangue rimane all’interno dei vasi, la pressione è elevata e il sangue è quindi pompato più velocemente. Opportune valvole evitano il reflusso del sangue nei vasi. Questo tipo di sistema circolatorio si trova nei Vertebrati e in diversi tipi di Invertebrati come anellidi, calamari e ottopodi. Il sangue usualmente contiene cellule e plasma. Le cellule del sangue dei vertebrati contengono emoglobina.

37
Vediamo uno schema del sistema circolatorio nei diversi gruppi di vertebrati. In tutti i vertebrati il cuore è formato da camere atriali e camere ventricolari

38
Le branchie contengono numerosi capillari per gli scambi gassosi. Quando esce dalle branchie la pressione del sangue è molto bassa. Da quì il sangue va alle diverse parti del corpo.

39
Il sangue dai polmoni va all’atrio sx. Il sangue refluo dal corpo (sistemico) va all’atrio dx. Entrambi gli atri si svuotano nel ventricolo. Il vantaggio di questo sistema è che il sangue ha una pressione elevata nei vasi che portano sia ai polmoni che al corpo.

40
Nella maggior parte dei rettili il ventricolo è parzialmente diviso. Questo riduce il mixing fra sangue ossigenato e non ossigenato. La parziale divisione è rappresentata dalla linea tratteggiata.

41
Coccodrilli, uccelli e mammiferi hanno quattro camere e due pompe separate di cui una per la circolazione polmonare e una per la circolazione sistemica. La pressione rimane sempre elevata.

42
Generalità sul sistema cardiovascolare umano

43
Il sistema cardiovascolare è costituito da una pompa, il cuore, da un sistema di condotti di distribuzione e di raccolta e una vasta rete di vasi. Vedremo che il cuore è formato da due pompe connesse fra loro da condotti cilindrici (vasi) in cui scorre il sangue: la parte destra spinge il sangue ai polmoni per l’ossigenazione (circolo polmonare) la parte sinistra spinge il sangue ai vari distretti corporei (circolo sistemico) Opportune valvole cardiache assicurano l’unidirezionalità. Vedremo inoltre che la gittata cardiaca è intermittente, mentre il flusso periferico è continuo. Il cuore pompa 7200 l in 24 h
 in cui scorre il sangue: la parte destra spinge il sangue ai polmoni per l’ossigenazione (circolo polmonare) la parte sinistra spinge il sangue ai vari distretti corporei (circolo sistemico) Opportune valvole cardiache assicurano l’unidirezionalità. Vedremo inoltre che la gittata cardiaca è intermittente, mentre il flusso periferico è continuo. Il cuore pompa 7200 l in 24 h.")
45
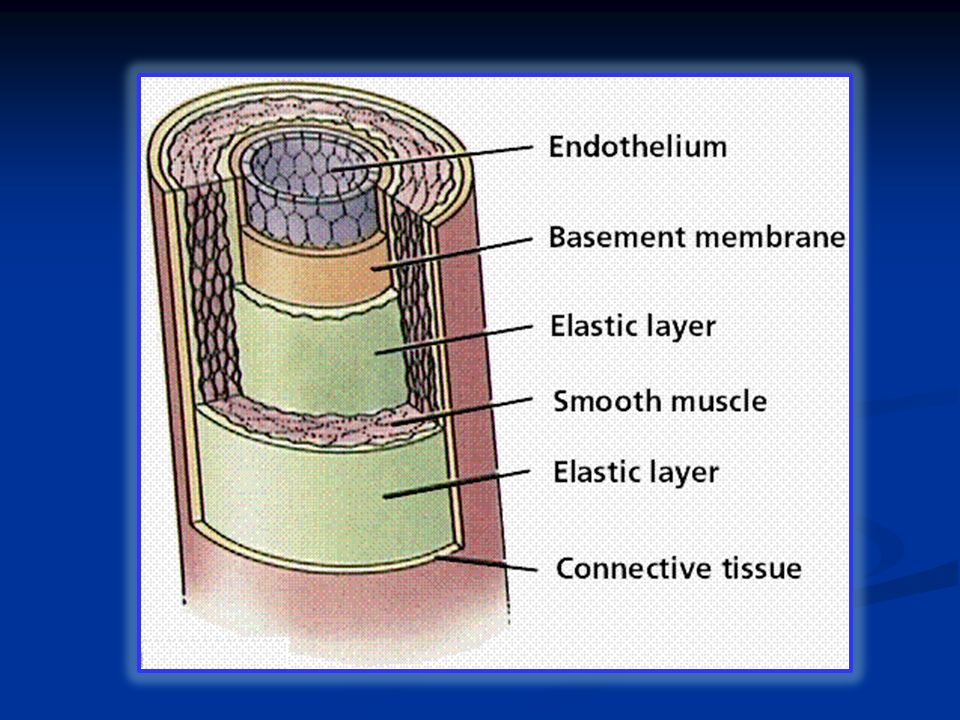
Il sangue scorre nelle arterie il cui diametro diminuisce progressivamente verso la periferia insieme ad una variazione della struttura. Dalle arteriole e dai capillari il sangue torna al cuore attraverso il sistema venoso. Anche le vene modificano diametro e struttura e assicurano il ritorno venoso grazie alle vene cave superiore ed inferiore. I condotti del sistema circolatorio sono distribuiti in serie (aorta, arterie, arteriole, capillari) e in parallelo (arteriole totali e capillari totali)
 e in parallelo (arteriole totali e capillari totali).")
46
Q=ΔP·R Circolazione in serie: il flusso Q è costante ma varia il gradiente pressorio Circolazione in parallelo: varia la distribuzione del flusso ai vari distretti Circolazione in parallelo Circolazione in serie

48
Quali sono le funzioni del sistema circolatorio? Trasporto di sostanze nutritizie ai tessuti Eliminazione delle sostanze di rifiuto Trasporto di ormoni ai tessuti bersaglio In generale, mantenimento di un ambiente ideale per la sopravvivenza ed il buon funzionamento cellulare

49
In generale si può dire che la funzione principale del sistema circolatorio è quella di fornire un flusso di sangue continuo e costante attraverso i tessuti

50
Vediamo quali sono le funzioni dei vasi sanguigni. Arterie: trasporto di sangue a regimi di alta pressione Arteriole: valvole di controllo di resistenza del sistema arterioso Capillari: scambio di fluidi, nutrienti, elettroliti ed ormoni Venule: raccolta di sangue refluo Vene: trasporto di sangue al cuore a regimi di bassa pressione

51
Ventricolo sx Aorta Grandi arterie Piccole arterie Arteriole Capillari Venule Piccole vene Grandi vene Vene cave Atrio dx Circolo polmonare Vene polmonari Atrio sx Ventricolo dx Arterie polmonari

52
VasiNumero Area sezione trasversa (cm 2 ) Volume di sangue (%) SISTEMICI Aorta12.8 Arterie40-110.0004011 Arteriole 2.8 10 6 55 Capillari 2.7 10 9 250005 Venule 10 7 785 Vene 660.000-10 6 63167 Cave23.1 Arterie e arteriole 1-1.5 10 6 1373 Capillari 2.7 10 9 13574 Vene-venule 2 10 7 2105 CUOREAtri-ventricoli 2 + 2 5 POLMONARI
 Volume di sangue (%) SISTEMICI Aorta12.8 Arterie Arteriole Capillari Venule Vene Cave23.1 Arterie e arteriole Capillari Vene-venule CUOREAtri-ventricoli POLMONARI")
53
Aorta Arteria Vena Cava Arteriola Arteriolaterminale Venula Capillare

56
Notare la relazione inversa fra velocità e area della sezione traversa e la caduta di pressione a livello delle arteriole

57
Valori pressori medi (mmHg) Velocità media (cm/s) Aorta8531 Grandi arterie -----20 Arteriole38----- Capillari250.5 Venosa5----- Vene cave -----13
 Velocità media (cm/s) Aorta8531 Grandi arterie Arteriole Capillari250.5 Venosa Vene cave")
58
Pressione sanguigna nelle diverse parti del sistema circolatorio Pressione sanguigna nelle diverse parti del sistema circolatorio

59
Il flusso del sangue in ogni tessuto è determinato dalle necessità del tessuto stesso. La gittata cardiaca è controllata principalmente dalla somma di tutti i flussi sanguigni locali. In genere la pressione arteriosa è controllata in modo indipendente sia dal controllo del flusso locale sia dal controllo della gittata cardiaca.

60
Ricordiamo che si intende per arterie i vasi che si allontanano dal cuore. Arterie sistemiche: trasportano sangue ossigenato ai distretti corporei Arterie polmonari: trasportano sangue non ossigenato ai polmoni Si intende per vene i vasi che arrivano al cuore. Vene sistemiche: trasportano sangue non ossigenato refluo dai distretti corporei Vene polmonari: trasportano sangue ossigenato dai polmoni all’atrio sx.

62
Il cuore: configurazione e rapporti Il cuore è un organo muscolare sacciforme, diviso in due parti, destra e sinistra, completamente separate fra di loro. Ogni metà, come vedremo, è formata da due camere, una superiore detta atrio ed una inferiore detta ventricolo. Il cuore è situato nella cavità toracica in uno spazio compreso fra i due polmoni, detto mediastino, ed è appoggiato sul centro frenico del diaframma. È spostato anteriormente e verso sinistra.

63
Il mediastino è la zona mediana della cavità toracica, posta tra i due polmoni e delimitata in basso dal diaframma. Il mediastino anteriore contiene il cuore ed i vasi sanguigni che da questo si dipartono, il mediastino posteriore contiene la trachea, i bronchi, l‘esofago, il dotto toracico e l‘aorta discendente. Il mediastino è la zona mediana della cavità toracica, posta tra i due polmoni e delimitata in basso dal diaframma. Il mediastino anteriore contiene il cuore ed i vasi sanguigni che da questo si dipartono, il mediastino posteriore contiene la trachea, i bronchi, l‘esofago, il dotto toracico e l‘aorta discendente.

64
Il cuore ha forma conica ed è leggermente appiattito in senso antero- posteriore. La punta o apice volge in basso, in avanti e a sinistra, mentre la base è in alto e guarda a destra e indietro. Pesa circa 200-300 gr e contiene 500ml di sangue. Il cuore è completamente avvolto dal pericardio, una membrana sierosa che forma un sacco a doppia parete, bagnata internamente da un velo di liquido pericardico Il cuore è completamente avvolto dal pericardio, una membrana sierosa che forma un sacco a doppia parete, bagnata internamente da un velo di liquido pericardico.

65
Prende rapporto tramite il pericardio con la faccia mediastinica dei polmoni lateralmente; con l’esofago, i bronchi extrapolmonari, l’aorta toracica, il dotto toracico e la vena azigos posteriormente; anteriormente è a diretto contatto con la regione costo-sternale della parete toracica; inferiormente appoggia sul diaframma e con l’interposizione di questo contrae rapporto con stomaco e fegato.

67
Conformazione interna del cuore L’atrio destro comunica con il sottostante ventricolo destro e lo stesso vale a sinistra. Non esiste comunicazione fra cuore destro e cuore sinistro. Essi infatti sono separati da un setto che comprende una porzione inter-atriale ed una porzione inter- ventricolare. Gli atri comunicano con i sottostanti ventricoli attraverso le valvole atrio-ventricolari che vedremo più avanti. La valvola destra presenta tre cuspidi ed è detta tricuspide, mentre la sinistra ne presenta due ed è detta bicuspide o mitrale

68
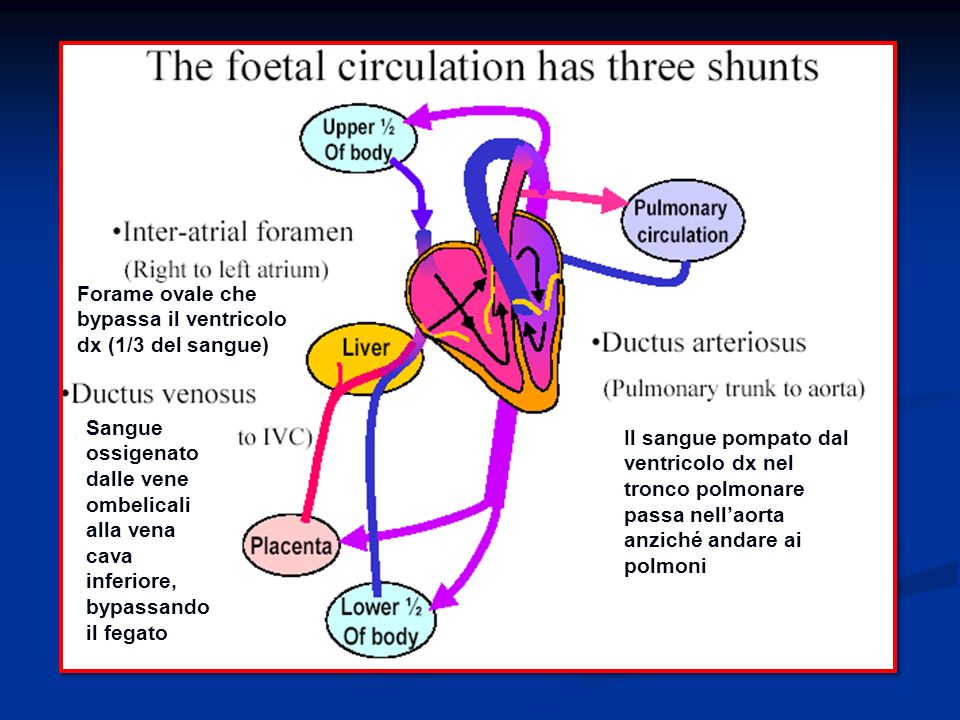
Circolazione fetale

69
Attraverso la vena cava inferiore il sangue penetra nell'atrio destro del cuore. L'atrio destro fetale, in corrispondenza dello sbocco della vena cava inferiore, presenta una formazione chiamata “valvola di Eustachio”. Per mezzo di essa la corrente sanguigna proveniente dalla vena cava inferiore viene avviata verso il setto interatriale, nel quale è presente il forame ovale o di Botallo che mette in comunicazione i due atri.

71
Sangue ossigenato dalle vene ombelicali alla vena cava inferiore, bypassando il fegato Forame ovale che bypassa il ventricolo dx (1/3 del sangue) Il sangue pompato dal ventricolo dx nel tronco polmonare passa nell’aorta anziché andare ai polmoni
 Il sangue pompato dal ventricolo dx nel tronco polmonare passa nell’aorta anziché andare ai polmoni")
72
La superficie interna degli atri è piuttosto liscia mentre la superficie interna dei ventricoli presenta numerosi rilievi intrecciati ed anastomizzati detti trabecole carnee. Alcune trabecole si estroflettono nelle cavità ventricolari in forma di tozzi coni, i muscoli papillari che vanno a fissarsi con le corde tendinee alla faccia inferiore e sui margini liberi delle cuspidi valvolari.

75
Il cuore è un organo cavo sacciforme la cui parete è formata da tre tonache sovrapposte. La più esterna detta EPICARDIO è data dal foglietto viscerale del sacco pericardico, mentre la più interna, l’ENDOCARDIO, è una lamina endoteliale che riveste la superficie di atri (endocardio più spesso) e ventricoli (endocardio più sottile) e delle lamine valvolari. La tonaca intermedia o MIOCARDIO è formata da tessuto muscolare cardiaco i cui fasci prendono attacco allo scheletro fibroso del cuore
 e ventricoli (endocardio più sottile) e delle lamine valvolari. La tonaca intermedia o MIOCARDIO è formata da tessuto muscolare cardiaco i cui fasci prendono attacco allo scheletro fibroso del cuore.")
76
Endocardio atriale Endocardio ventricolare

78
Il miocardio è formato da tessuto muscolare striato cardiaco, le cui fibre formano un reticolo tridimensionale, in modo che le fibre siano orientate in tutte le direzioni garantendo un perfetto svuotamento delle camere cardiache. Il miocardio atriale è piuttosto sottile e formato da due strati: - lo strato interno forma un sacco incompleto, proprio di ciascun atrio - lo strato più esterno è formato da fasci ad andamento trasversale in comune ad entrambi gli atri

79
La muscolatura dei ventricoli forma due sacchi conoidi affiancati (fasci propri) con la base in corrispondenza delle valvole AV, rivestiti all’interno e all’esterno da fasci muscolari comuni. I fasci esterni si portano con andamento obliquo dagli osti AV fino all’apice del cuore, ruotano di 180˚ e si portano all’interno dei sacchi propri, distribuendosi sotto l’endocardio. Trabecole carnee e muscoli papillari sono formati da queste fibre. Il miocardio del ventricolo sx è molto più sviluppato di quello del ventricolo dx

81
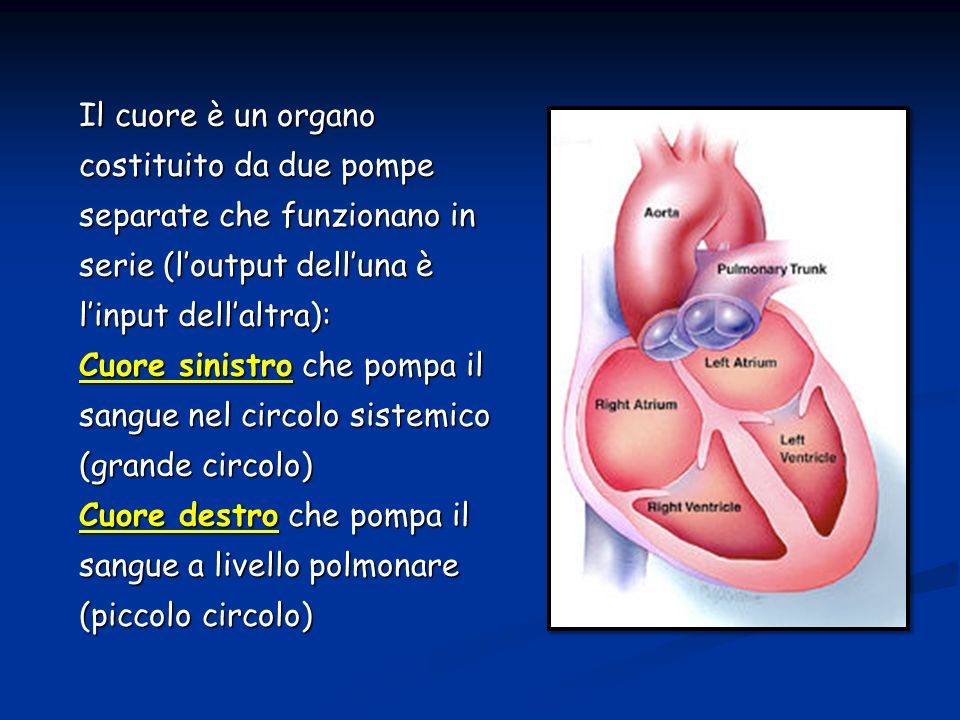
Il cuore è un organo costituito da due pompe separate che funzionano in serie (l’output dell’una è l’input dell’altra): Cuore sinistro che pompa il sangue nel circolo sistemico (grande circolo) Cuore destro che pompa il sangue a livello polmonare (piccolo circolo)
: Cuore sinistro che pompa il sangue nel circolo sistemico (grande circolo) Cuore destro che pompa il sangue a livello polmonare (piccolo circolo)")
82
A sua volta ciascuna parte del cuore è costituita da una camera superiore detta ATRIO e da una camera inferiore detta VENTRICOLO. L’atrio costituisce una debole pompa che contribuisce a far passare il sangue nel ventricolo, mentre il ventricolo è la pompa principale che imprime la forza necessaria a pompare il sangue in circolo.

83
Il cuore è formato da tre tipi di muscolo cardiaco: -Muscolo atriale Muscolo ventricolare -Muscolo ventricolare -Fibre eccitatorie conduttive specializzate con scarsa contrattitlità Il muscolo atriale e quello ventricolare hanno caratteristiche morfologiche simili a quelle del muscolo scheletrico, ma con una contrazione che dura più a lungo. Le fibre conduttive si contraggono poco, perché hanno pochi elementi contrattili, ma godono invece di ritmicità e vari gradi di velocità di conduzione.

84
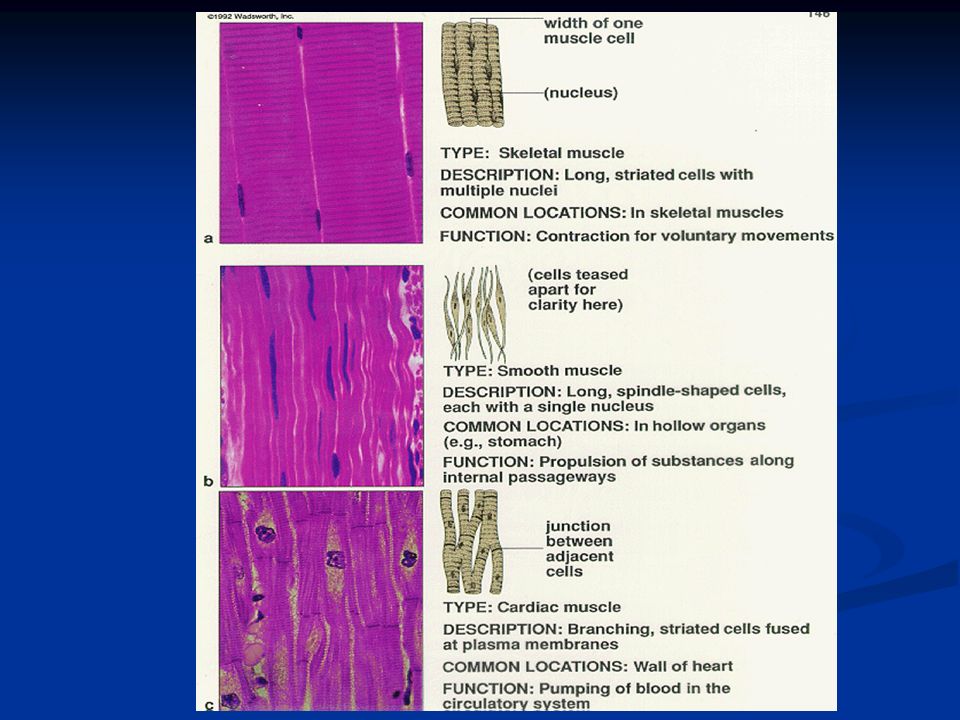
Il muscolo cardiaco è striato come il muscolo scheletrico, contiene similmente actina e miosina che si interdigitano come succede nel muscolo scheletrico. Le fibre si ramificano più volte e si interdigitano fra loro. Dal punto di vista contrattile il muscolo cardiaco è assai diverso da quello scheletrico

87
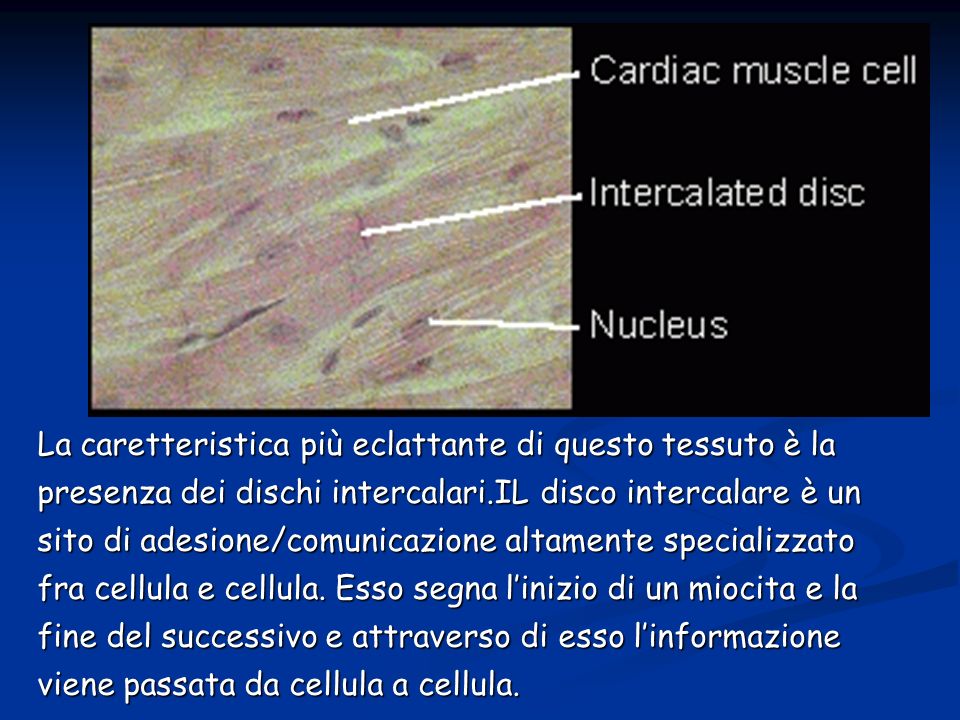
La caretteristica più eclattante di questo tessuto è la presenza dei dischi intercalari.IL disco intercalare è un sito di adesione/comunicazione altamente specializzato fra cellula e cellula. Esso segna l’inizio di un miocita e la fine del successivo e attraverso di esso l’informazione viene passata da cellula a cellula.

88
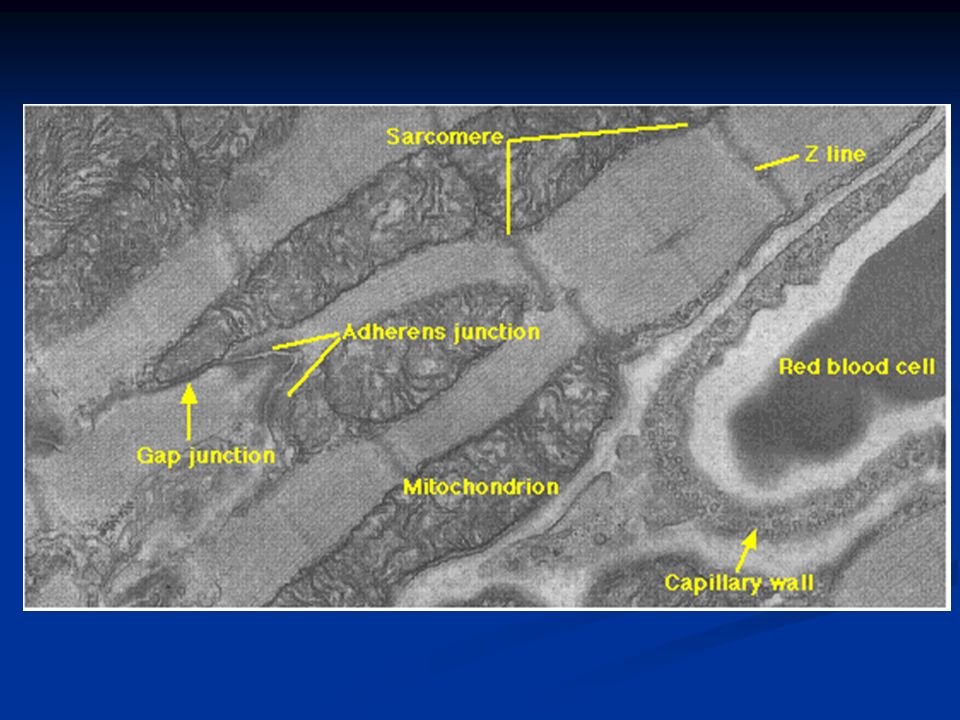
I dischi intercalari sono zone molto frastagliate caratterizzate dalla presenza di gap junctions e desmosomi. Le prime garantiscono una veloce progressione del segnale elettrico, mentre i secondi sono zone di forte adesione meccanica attraverso le quali la forza generata da una cellula viene trasmessa alle altre

89
I dischi intercalari sono vie a bassa resistenza (1/400 della resistenza attraverso la membrana esterna). Gli ioni si muovono quindi facilmente lungo l’asse longitudinale e lo spike si propaga facilmente da una fibra a quella successiva. Il muscolo cardiaco può essere considerato un sincizio per questa facilità di comunicazione. Va però notato che il cuore è in realtà formato da:

90
sincizio atriale sincizio ventricolare separati fra loro da tessuto fibroso valvolare. Questo allo scopo di sfalsare temporalmente la contrazione degli atri dalla contrazione dei ventricoli

93
La contrazione si propaga da atri a ventricoli con un ritardo di circa 1/6 di secondo, in modo da garantire una completa contrazione ventricolare, DOPO una completa contrazione atriale. Il segnale elettrico, come vedremo, è veicolato dal fascio atrio-ventricolare (A-V).
..")
94
Nonostante le forti similitudine fra muscolo scheletrico e muscolo cardiaco, esistono anche fondamentali differenze che ne determinano le proprietà contrattili. Vediamole i. Il muscolo scheletrico è capace di sostenere una contrazione prolungata in regime tetanico; il muscolo cardiaco non può assolutamente giungere in regime tetanico e la sua contrazione è solo transitoria, per poi rilassarsi nuovamente.

95
ii. La disposizione delle fibre nel miocardio è interconnessa con continue ramificazioni e presenza di abbondanti fibre di collagene, mentre il muscolo scheletrico presenta una configurazione in fibre parallele e minor contenuto in collagene iii. Il meccanismo di eccitazione è unico ed intrinseco al cuore stesso; le fibre vengono incendiate e reclutate tutte insieme con l'ordine prestabilito per la contrazione

96
iv. Il muscolo scheletrico è molto più reattivo: la velocità con cui si sviluppa forza a seguito di una stimolazione è di un ordine di grandezza superiore al miocardio. v. A completa differenza del muscolo scheletrico, le cellule muscolari cardiache sono mononucleate e dotate di molti mitocondri. Il muscolo deve contrarsi ripetitivamente per l'intera vita e non può contrarre un debito significativo di ossigeno.

97
Cellula miocardica Anche le cellule miocardiche sono formate da sarcomeri con filamenti spessi di miosina e filamenti sottili di actina. Vale anche in questo caso la teoria dello slittamento dei filamenti. La relazione lunghezza/tensione è simile allo striato con una lunghezza di riposo di 2-2.4μm. Vedremo però che mentre lo striato lavora in condizioni di lunghezza ideale per sviluppare la tensione massima, la fibrocellula cardiaca lavora ad una lunghezza inferiore a l 0. Le cellule sono circa 5-15μm di diametro e 30-50μm di lunghezza.

98
Nelle cellule cardiache il sistema dei tubuli T è molto meno sviluppato e dà luogo a diadi (1 cisterna più il tubulo) invece che a triadi. Inoltre questo sistema si apre in corrispondenza della linea Z e non fra banda A e banda I come nello scheletrico. Le fibre muscolari cardiache sono essenzialmente aerobiche perché il muscolo cardiaco non ha mai fasi di riposo e quindi sono ricche di mitocondri e mioglobina.

99
Diagramma lunghezza-tensione per un singolo sarcomero Tensione sviluppata (%) 100 50 1234 Lunghezza sarcomero (μm) A BC D N.B. Contrazione di tipo isometrico. Il sarcomero è fissato ad una certa l e si misura la max tensione sviluppata N.B. Contrazione di tipo isometrico. Il sarcomero è fissato ad una certa l e si misura la max tensione sviluppata

100
Equivalente della lunghezza del sarcomero Equivalente dello sviluppo di tensione

101
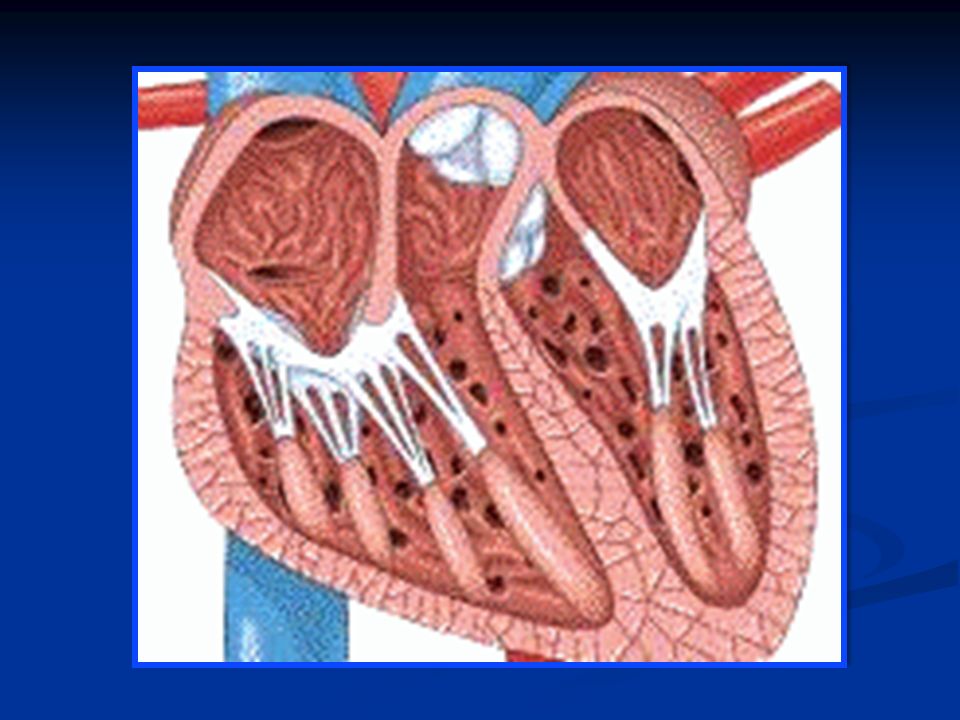
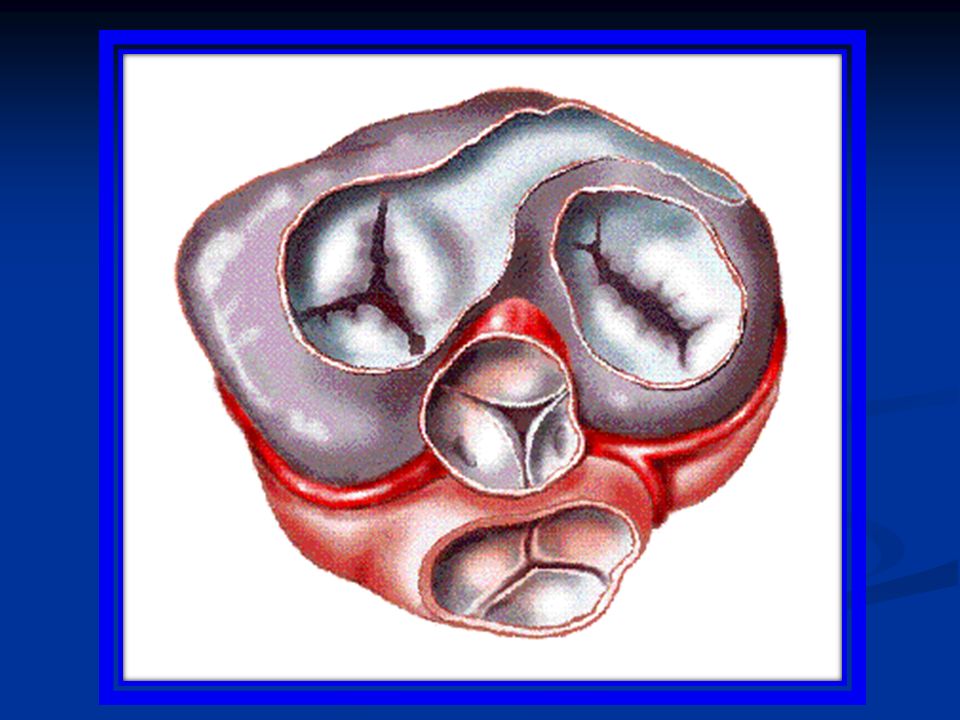
Funzioni delle valvole cardiache Nel cuore si distinguono due tipi di valvole: Valvole atrio-ventricolari: sono situate fra la camera atriale e la camera ventricolare ed impediscono il reflusso di sangue dal ventricolo all’atrio durante la sistole Valvole semilunari (aortica e polmonare): sono situate fra ventricolo e vaso arterioso di uscita (aorta e arteria polmonare rispettivamente) ed impediscono il reflusso di sangue dall’arteria al ventricolo durante la diastole.
: sono situate fra ventricolo e vaso arterioso di uscita (aorta e arteria polmonare rispettivamente) ed impediscono il reflusso di sangue dall’arteria al ventricolo durante la diastole.")
102
Valvola AV Valvola aortica Valvola polmonare Valvola AV

105
È importante ricordare che tutte queste valvole si aprono e si chiudono PASSIVAMENTE in presenza di un gradiente pressorio. Per ragioni puramente anatomiche le valvole AV non hanno bisogno di reflusso di sangue per chiudersi, mentre le semilunari (più spesse) richiedono un reflusso minimo per alcuni millisecondi
 richiedono un reflusso minimo per alcuni millisecondi.")
106
Come sono fatte queste valvole? Le valvole AV sono dette Valvola tricuspide fra atrio e ventricolo destro Valvola bicuspide o mitrale fra atrio e ventricolo sinistro. Sono formate rispettivamente da tre e due lembi valvolari che richiudono l’apertura della valvola, ciascun lembo con una superficie molto estesa in modo che la valvola sia abbondantemente chiusa. Le cuspidi della valvola sono collegate ai muscoli papillari attraverso le corde tendinee, che si contraggono durante la contrazione ventricolare.

107
Le corde tendinee non servono a far richiudere la valvola (che si richiude per gradiente pressorio), ma piuttosto servono ad impedire che i lembi si ripieghino eccessivamente verso l’atrio. In caso di danneggiamento delle corde (come nella mucopolisaccaridosi) o dei muscoli papillari intervengono danni anche letali.
 o dei muscoli papillari intervengono danni anche letali..")
109
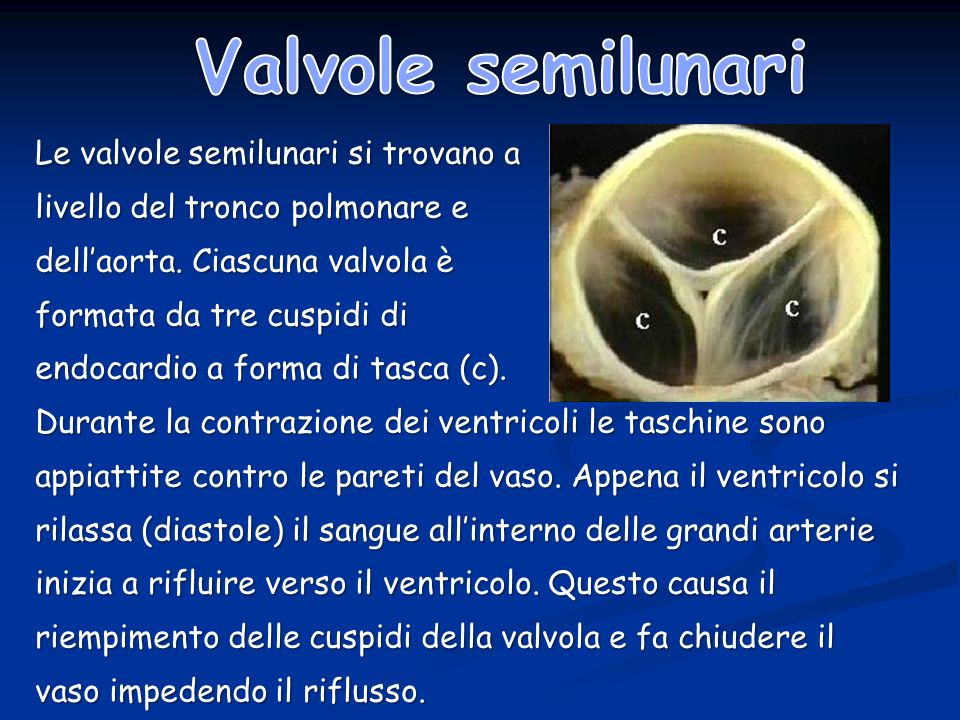
Le valvole semilunari si trovano a livello del tronco polmonare e dell’aorta. Ciascuna valvola è formata da tre cuspidi di endocardio a forma di tasca (c). Durante la contrazione dei ventricoli le taschine sono appiattite contro le pareti del vaso. Appena il ventricolo si rilassa (diastole) il sangue all’interno delle grandi arterie inizia a rifluire verso il ventricolo. Questo causa il riempimento delle cuspidi della valvola e fa chiudere il vaso impedendo il riflusso.
. Durante la contrazione dei ventricoli le taschine sono appiattite contro le pareti del vaso. Appena il ventricolo si rilassa (diastole) il sangue all’interno delle grandi arterie inizia a rifluire verso il ventricolo. Questo causa il riempimento delle cuspidi della valvola e fa chiudere il vaso impedendo il riflusso..")
110
S emilunar Valve

112
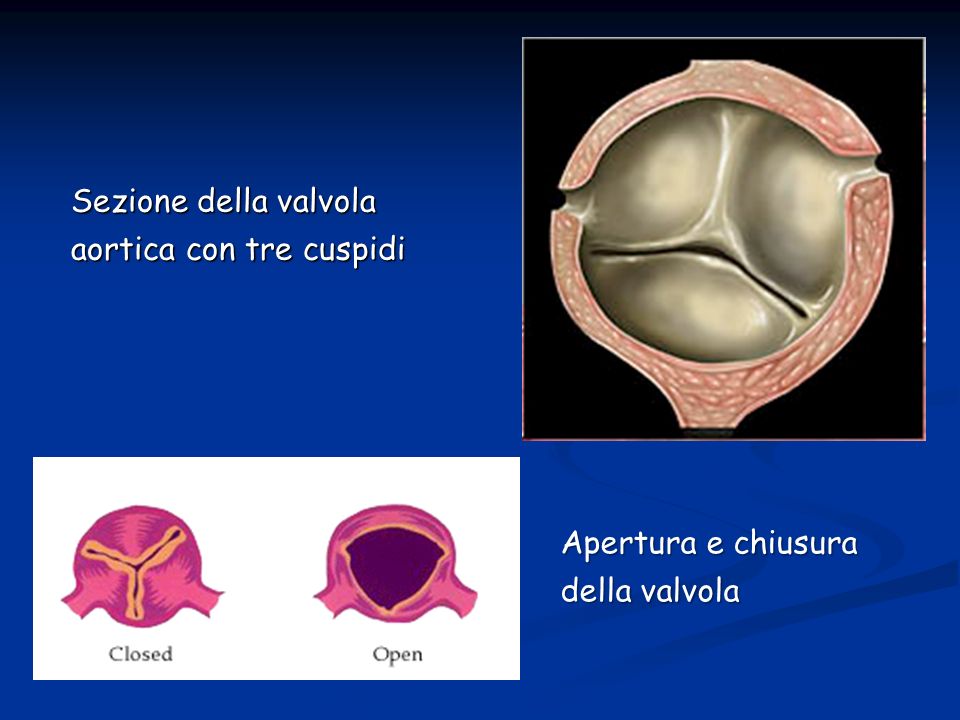
Sezione della valvola aortica con tre cuspidi Apertura e chiusura della valvola

113
Quindi le valvole semilunari differiscono dalle valvole AV perché: Le valvole semilunari si chiudono molto più bruscamente delle AV a causa dei valori di pressione molto più elevati nei vasi arteriosi La velocità di eiezione del sangue attraverso le valvole semilunari è molto maggiore che nelle AV a causa della minore apertura Le valvole semilunari sono più facilmente soggette ad usura che le AV

114
Valvola artificiale

115
Insieme dei meccanismi attraverso i quali l’attività elettrica del cuore causa la contrazione delle fibre muscolari.

116
Aumento della concentrazione del Ca 2+ intracellulare Legame del Ca 2+ con la troponina Liberazione della tropomiosina Sliding dei filamenti STRIATO CARDIACO Ingresso dai canali tipo L della membrana plasmatica e dei tubuli T e liberazione di calcio dal reticolo (CICR) Liberazione dal reticolo sarcoplasmatico soltanto
 Liberazione dal reticolo sarcoplasmatico soltanto")
117
Come accade per il muscolo striato, il potenziale si propaga all’interno della fibra muscolare cardiaca attraverso il sistema dei tubuli T. Questo, a sua volta, eccita la membrana del reticolo sarcoplasmatico, causando il rilascio di ioni Ca 2+ nel sarcoplasma con un meccanismo CICR mediato da RyR1 nello scheletrico e RyR2 nel cardiaco. Questi ioni determinano lo slittamento dei filamenti, come succede nel muscolo striato.

118
Le cisterne terminali del reticolo sarcoplasmatico (SR) si avvicinano al sarcolemma. I canali del calcio tipo L del sarcolemma e i canali del calcio sensibili al calcio(RyR2) di SR sono separati da circa 12nm RyR2 (isoforma del m. cardiaco) L-type Ca channels
 di SR sono separati da circa 12nm RyR2 (isoforma del m. cardiaco) L-type Ca channels.")
119
MUSCOLO STRIATO

120
MUSCOLO CARDIACO

121
La differenza fondamentale dal muscolo striato è che la maggior parte del calcio necessario alla contrazione deriva da diffusione attraverso i tubuli T durante lo spike, e non dal reticolo sarcoplasmico. Questo perché il reticolo sarcoplasmico del muscolo cardiaco è molto meno sviluppato di quello scheletrico (volume circa 25 volte inferiore) e il calcio storato e liberato è insufficiente ad assicurare una corretta contrazione.
 e il calcio storato e liberato è insufficiente ad assicurare una corretta contrazione..")
122
La forza di contrazione del muscolo cardiaco dipende soprattutto dalla concentrazione del calcio extracellulare.

123
NOTA BENE

124
Il fosfolambano regola la pompa

125
Va notato che nel muscolo cardiaco non esiste una triade come nello striato (fra banda A e banda I), ma piuttosto una diade a livello di stria Z e il sistema è molto meno sviluppato.
, ma piuttosto una diade a livello di stria Z e il sistema è molto meno sviluppato.")
126
Alla fine del potenziale d’azione il calcio è prontamente sequestrato nel reticolo sarcoplasmico grazie ad una Ca-ATPasi e scambiato con uno scambiatore Na-Ca all’esterno della cellula. Il muscolo cardiaco inizia a contrarsi pochi ms dopo l’inizio dello spike e la contrazione finisce pochi ms dopo la fine dello spike, per una durata di circa 200ms nel muscolo atriale e 300ms nel muscolo ventricolare.

127
All’aumentare della frequenza cardiaca, il fattore pesantemente alterato è proprio il rilassamento perché non si riesce a smaltire il calcio intracellulare, tanto che ad alte frequenze di battito, il cuore non ha il tempo di rilassarsi completamente.

128
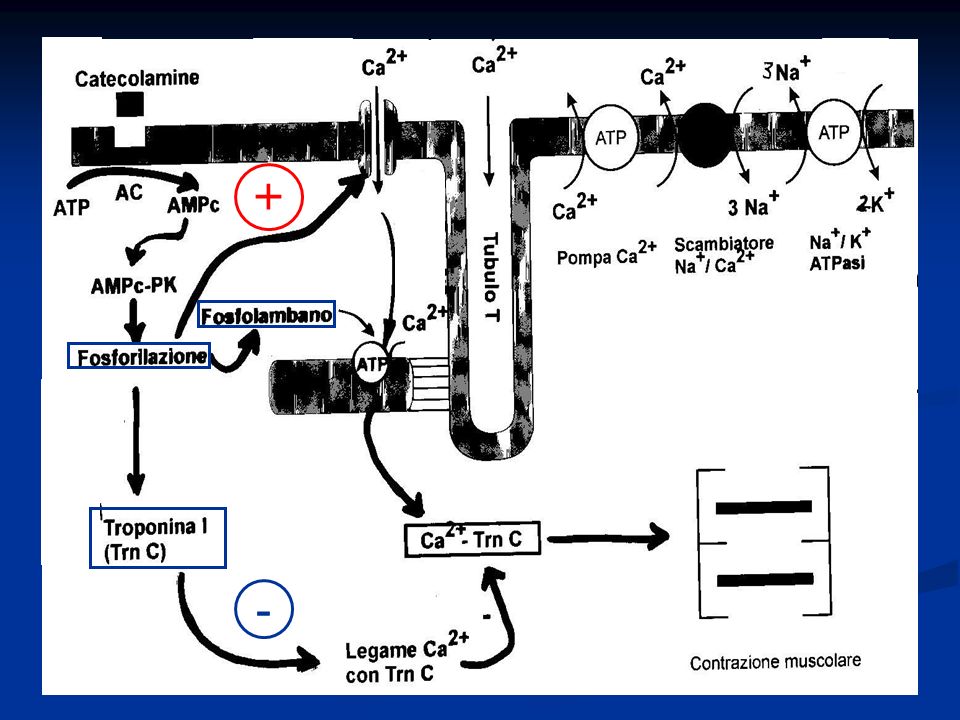
Il calcio citosolico può essere aumentato in vario modo, causando un aumento della forza di contrazione (effetto inotropo positivo): Le catecolamine si legano al recettore β- adrenergico e, attraverso un sistema di secondi messaggeri, permettono l’aumento del calcio per fosforilazione dei canali DHP-sensibili (L-type) I glicosidi cardiaci (digitale) agiscono sulla Na-K ATPasi bloccandola, con conseguente aumento del Na citosolico. Lo scambiatore Na-Ca funziona meno bene ed il calcio tende ad accumularsi con conseguente effetto INOTROPO positivo

129
Va notato che ad un effetto inotropo positivo delle catecolammine fa seguito anche un effetto lusitropo positivo (aumento della velocità di rilassamento) che permette una più veloce rimozione del calcio intracellulare. Questo è possibile grazie all’aumento di attività della Ca-ATPasi conseguente alla fosforilazione della proteina regolatrice fosfolambano e anche grazie alla maggior dissociazione di calcio dalla troponina che si verifica quando la troponina è fosforilata.

131
- - +

132
Va ricordato che le fosforilazioni operate dalla proteina-chinasi AMPc-dipendente (PKA) servono per incrementare sia la velocità della contrazione (attraverso la fosforilazione dei canali del Ca 2+ del sarcolemma che causano un aumentato ingresso di calcio nella cellula), sia la velocità del rilasciamento (attraverso la fosforilazione del fosfolambano che aumenta la captazione del Ca 2+ dal reticolo sarcoplasmatico e attraverso la fosforilazione della TrnI che inibisce la fissazione del Ca 2+ alla TrnC)
 servono per incrementare sia la velocità della contrazione (attraverso la fosforilazione dei canali del Ca 2+ del sarcolemma che causano un aumentato ingresso di calcio nella cellula), sia la velocità del rilasciamento (attraverso la fosforilazione del fosfolambano che aumenta la captazione del Ca 2+ dal reticolo sarcoplasmatico e attraverso la fosforilazione della TrnI che inibisce la fissazione del Ca 2+ alla TrnC)")
Presentazioni simili


































, specializzato per la distribuzione di:>")
, specializzato per la distribuzione di: gas.>")



