Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il trasporto attraverso la membrana
DT 1

3
Proteine di membrana CANALI ENZIMI RECETTORI TRASPORTATORI

4
Many Functions of Membrane Proteins
Outside Plasma membrane Inside Transporter Enzyme activity Cell surface receptor Signal transduction - transmitting a signal from outside the cell to the cell nucleus, like receiving a hormone which triggers a receptor on the inside of the cell that then signals to the nucleus that a protein must be made. Cell surface identity marker Cell adhesion Attachment to the cytoskeleton 4

5
In biochimica, un cambiamento conformazionale è un cambiamento nella forma di una macromolecola, spesso indotto da fattori ambientali. Una macromolecola è generalmente flessibile e dinamica. Può cambiare forma in risposta ai cambiamenti del suo ambiente o di altri fattori; ogni possibile forma è chiamata conformazione e una transizione tra loro è chiamata cambiamento conformazionale. I fattori che possono indurre tali modifiche includono: temperatura, pH, voltaggio, concentrazione di ioni, fosforilazione, il legame di un ligando (ione, proteina, DNA etc, etc) Molte proteine subiscono grandi cambiamenti conformazionali, come parte della loro funzionalità. Individuare e seguire questi cambiamenti è importante per comprendere il modo in cui le proteine funzionano.
 Molte proteine subiscono grandi cambiamenti conformazionali, come parte della loro funzionalità. Individuare e seguire questi cambiamenti è importante per comprendere il modo in cui le proteine funzionano.")
6
Il calcio è il minerale più abbondante nel nostro corpo ma la concentrazione di Ca2 + all'interno delle nostre cellule è estremamente bassa (~ 10-7M). Pompe di calcio e canali ionici lavorano duramente per stabilire e mantenere questo forte differenza. All'interno della cellula, il calcio è spazialmente limitato, immagazzinato in alcuni ma non in tutti gli organelli, pronto per il rilascio al momento opportuno. Proteine che legano e regolano e sono regolate dal calcio sono un grande famiglia e ampiamente distribuita. Molti membri di questa famiglia condividono una caratteristica forma manubrio, due domini globulari collegati da una regione linker flessibile. La Calmodulina, è una proteina sensibile al calcio che lega quattro + ioni Ca2, due con alta affinità e due con bassa affinità. I siti ad alta affinità possono essere riempiti a concentrazioni di calcio basse, ma quando il calcio viene rilasciato dai depositi intracellulari, anche i siti di bassa affinità vengono occupati, inducendo un grande cambiamento conformazionale della proteina. Il cambiamento espone regioni non polari di calmodulina che può legarsi alle regioni non polari di una molecola bersaglio. Nella figura si possono osservare i cambiamenti conformazionali della calmodulina.

7
materiale entri (in) & prodotti o scarti escano (out)
cellula ha bisogno che materiale entri (in) & prodotti o scarti escano (out) OUT waste ammonia salts CO2 H2O products IN food carbohydrates sugars, proteins amino acids lipids salts, O2, H2O OUT IN
 & prodotti o scarti escano (out) OUT. waste. ammonia. salts. CO2. H2O. products. IN. food. carbohydrates. sugars, proteins. amino acids. lipids. salts, O2, H2O. OUT. IN.")
8
Diffusione attraverso il bilayer fosfolipidico
Possono attraversare direttamente la membrana NON POSSONO ATTRAVERSARE LA MEMBRANA 8

9
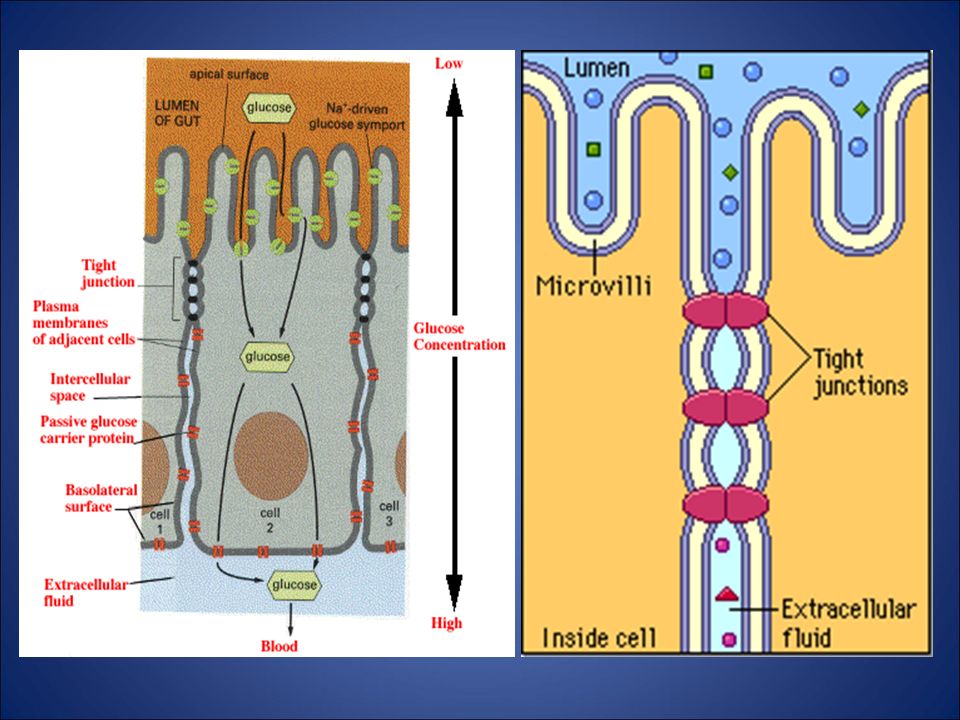
trasporto attraverso la membrana
il passaggio di molte sostanze è consentito dalla presenza di proteine vettrici che, modificando la loro conformazione, «traghettano» le molecole da un alto all’altro della membrana il trasporto contro gradienti di concentrazione richiede energia, come nel caso della pompa ionica Na-K acqua e ioni, a causa della loro polarità, passano solo attraverso canali proteici dedicati molecole voluminose come le proteine non possono attraversare la membrana

11
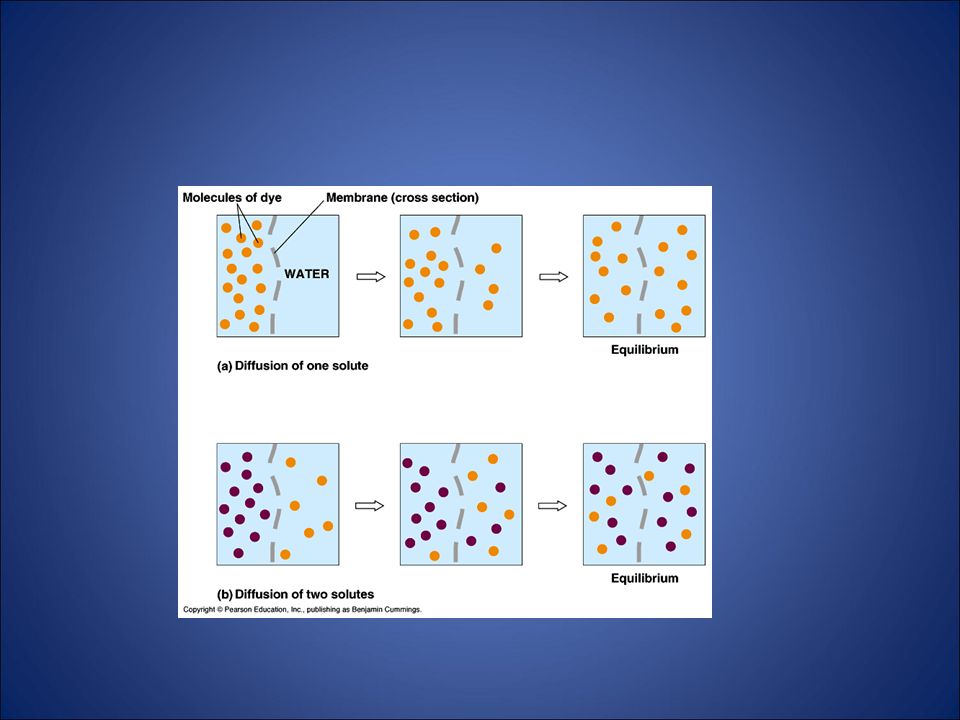
Diffusione La II Legge della Termodinamica governa i sistemi biologici
universe tends towards disorder (entropy) Movement from high concentration of that substance to low concentration of that substance. Move from HIGH to LOW concentration “trasporto passivo” Non richiede energia 11
 Movement from high concentration of that substance to low concentration of that substance. Move from HIGH to LOW concentration. trasporto passivo Non richiede energia. 11.")
13
Processi passivi: diffusione facilitata
Certi composti (troppo grandi per passare attraverso i canali di membrana) possono essere passivamente trasportati da Proteine CANALE e Proteine VETTRICI
 possono essere passivamente trasportati da Proteine CANALE e Proteine VETTRICI.")
14
I canali ionici permettono dei flussi molto rapidi e intensi
Tipo di trasportatore Velocità di turnover Pompe ioniche 2-5·102/sec Glucoso-permeasi /sec Canale del Na+ ~108/sec Batterorodopsina Segmento transmembrana

15
Struttura di base di un canale ionico
Lato extracellulare Lato citoplasmatico bilayer lipidico Poro acquoso Filtro di selettività Residui di zuccheri

16
In soluzione acquosa tutti gli ioni sono circondati da un alone di molecole d’acqua

17
I canali ionici sono estremamente selettivi nei confronti dei vari ioni
La selettività può essere conferita da una combinazione dei seguenti fattori: presenza di cariche elettriche fisse sulla parete interna del canale di intensità e densità specifiche raggio anidro dello ione grado di idratazione dello ione

18
Basi molecolari di selettività di un canale K+ nei confronti del K+ rispetto al Na+
K+ in H2O Na+ in H2O K+ nel poro Na+ nel poro Gli ioni K+, idrati in soluzione, perdono le molecole di H2O quando passano attraverso il filtro di selettività e formano dei legami di coordinazione con quattro O di gruppi carbonilici. Gli ioni Na+, essendo più piccoli, non possono coordinarsi perfettamente con questi O e quindi attraversano il canale solo raramente.

19
Sezione frontale (sinistra) e vista dall’alto (destra) di un canale selettivo per lo ione K+. Si possono osservare tre ioni K+ che stanno attraversando il poro del canale per diffusione ristretta in singola fila. Le molecole rappresentate da atomi rossi e bianchi sono molecole di H2O, alcune delle quali sono ancora legate agli ioni K+ a costituire l’alone di solvatazione.

20
Passaggio in singola file di ioni K+ attraverso il poro del canale

21
Anche nel caso dei canali ionici i flussi sono saturanti
Concentr. di Na+ o K+ (mM) Conduttanza di singolo canale (pS)
 Conduttanza di singolo canale (pS)")
22
Canali Ionici: classificazione in base alle modalità di apertura
Sempre aperti extracell. intracell. Voltaggio-dipendenti: rispondono a variazioni di Vm Chemio-dipendenti: rispondono a un messaggero extracellulare neurotrasmettitore Chemio-dipendenti: rispondono a un secondo messaggero (intracellulare) cAMP, cGMP, Ca2+, IP3, proteine G
 cAMP, cGMP, Ca2+, IP3, proteine G.")
23
Canale chemio-dipendente Canale voltaggio-dipendente
Canali ionici chemio- e voltaggio-dipendenti Canale chemio-dipendente Canale voltaggio-dipendente

24
Struttura di base di un canale voltaggio-dipendente
Lato citoplasmatico Lato extracellulare Sensore del voltaggio porta Filtro di selettività

25
Nei canali-recettori l’apertura è indotta dall’interazione con un ligando specifico (qui simboleggiato a forma di T), mentre i canali voltaggio-dipendenti si aprono quando il potenziale di membrana cambia; nel caso indicato si ha apertura passando da potenziali negativi a potenziali positivi (depolarizzazione). Canali ionici associati a recettori il legame di un liegando apre un canale ionico nella membrana

26
Il canale voltaggio-dipendente del sodio è
formato da un singolo lungo peptide: Il canale è costituito da 4 domini simili che si raggruppano a formare un poro che normalmente risulta chiuso: I canali per il sodio si aprono sempre per un breve tempo, meno di 1msec, reagendo ad una variazione di potenziale o alla presenza di "qualcosa" che ne modifica la struttura: Se la depolarizzazione permane, sapendo che il valore soglia è di circa -40mV, una porzione globulare della proteina-canale (ossia delle proteine che fungono da canale per il passaggio degli ioni) oscilla verso l'alto ed apre il poro, che allora si dice essere attivato. Quando la membrana si ripolarizza, il poro si chiude e, successivamente, la porzione globulare si sposta, tornando alla posizione iniziale: La variazione di carica all'interno ed all'esterno della membrana prende il nome di potenziale d'azione.
 oscilla verso l alto ed apre il poro, che allora si dice essere attivato. Quando la membrana si ripolarizza, il poro si chiude e, successivamente, la porzione globulare si sposta, tornando alla posizione iniziale: La variazione di carica all interno ed all esterno della membrana prende il nome di potenziale d azione.")
27
cystic fibrosis transmembrane conductance regulator (CFTR) gene.
CFTR controls chloride ion movement in and out in the lungs, liver, pancreas, digestive tract, reproductive tract, and skin Delta F508, the most common CF-causing mutation, occurs in the DNA sequence that codes for the first nucleotide-binding domain (NBD1).
.")
28
acido γ-amminobutirrico (GABA) è un γ-amminoacido principale neurotrasmettitore inibitorio nei mammiferi
 è un γ-amminoacido principale neurotrasmettitore inibitorio nei mammiferi")
30
Problemi clinici correlati ai sistemi di trasporto
Fibrosi cistica – canali del cloro epiteliali Arritmia cardiaca – canali del sodio e del potassio cardiaci Diabete mellito non-insulina-dipendente – trasportatore del glucosio nel tessuto adiposo

31
Le Proteine vettrici Aggancio della molecola da trasportare induce un cambio di conformazione della proteina vettrice rilascio della molecola nel citoplasma

32
Cinetica di saturazione
Se la concentrazione di glucoso da un lato della membrana sale rapidamente, tutte le proteine vettrici possono risultare occupate: la velocità di passaggio del soluto diventa costante

33
Processi di trasporto attivo
Consentono di spostare sostanze contro gradiente (di concentrazione, elettrico, ecc.) Per questo richiedono molta energia = ATP
 Per questo richiedono molta energia. = ATP.")
34
Trasporto attivo Molti modelli & mecccanismi ATP ATP antiport symport
Plants: nitrate & phosphate pumps in roots. Why? Nitrate for amino acids Phosphate for DNA & membranes Not coincidentally these are the main constituents of fertilizer Supplying these nutrients to plants Replenishing the soil since plants are depleting it antiport symport 34

35
ATPasi di trasporto Le ATPasi di trasporto, dette anche pompe ioniche, sfruttano l'idrolisi di ATP per trasportare ioni o protoni contro il gradiente di concentrazione. Si suddividono in tre classi, a seconda del meccanismo utilizzato e del substrato trasportato: ATPasi di tipo P; ATPasi di tipo F e ATPasi di tipo V. ATPasi di tipo P Le ATPasi di tipo P sono deputate principalmente al trasporto di ioni. Sono principalmente composte da un'unica catena polipeptidica che esplica sia la funzione idrolitica che quella trasportatrice. Vengono dette di tipo P per la presenza di un sito di fosforilazione a livello dell’aspartato, che nelle due diverse forme apre e chiude il canale per lo ione (Na+/K+, H+/K+, Ca++). ATPasi di tipo P sono presenti sia sulla membrana plasmatica che sulla membrana RE
. ATPasi di tipo P sono presenti sia sulla membrana plasmatica che sulla membrana RE.")
36
trasporto attivo Pompa di scambio Na-K

37
Conformational changes of Na,K-ATPase upon ATP binding

38
Pompa di scambio Na-K Fino al 40% dell’energia di una cellula a riposo
Canali per micron2: 4 K 100 Na 1000 pompe Na-K ATP ADP

39
La presenza di pompe ioniche provoca...
Pompa sodio-potassio Canale per il potassio Canale per il sodio proteina

40
distribuzione asim-metrica di ioni
i canali di membrana sono attraversati dagli ioni passivamente, a seconda dei loro gradienti distribuzione asim-metrica di ioni Cl- Na+ K+ le pompe ioniche determinano e mantengono gradienti di concentrazione ai due lati della membrana proteina

41
Potenziale transmembrana
+ + + + + + + + + + - - + + - - - - - - + + - - - La asimmetria nella distribuzione di ioni carichi elettricamente è all’origine di una differenza di potenziale fra i due lati della membrana che si trova normalmente in tutte le cellule - -70 mV -

42
ATPasi di tipo F e di tipo V
Le ATPasi di tipo V e di tipo F sono deputate al trasporto di PROTONI. Questa classe di ATPasi è strutturalmente più complessa, essendo formata da più subunità proteiche che si assemblano sulla membrana di molti compartimenti ed organelli cellulari (KD500). A differenza delle ATPasi di tipo P, quelle di tipo V e di tipo F non trasferiscono su un proprio aminoacido un residuo di fosofato.
. A differenza delle ATPasi di tipo P, quelle di tipo V e di tipo F non trasferiscono su un proprio aminoacido un residuo di fosofato.")
43
Le ATPasi di tipo F si trovano nei mitocondri dove sono responsabili della movimentazione di protoni per la sintesi energetica della cellula

44
Le ATPasi di tipo V sono presenti sui lisosomi, dove con il trasporto di protoni favorisce l'instaurazione di un ambiente idrolizzante acido.

45
Transport summary simple diffusion facilitated diffusion
ATP active transport 45

46
The Special Case of Water
Acquaporina 46

47
Il movimento effettivo di acqua attraverso una membrana cellulare è il risultato di due processi: diffusione e il flusso di massa. Come si ricorderà una membrana è lo spessore di un doppio strato di fosfolipidi. La dimensione di una molecola d'acqua le consenta di passare attraverso il doppio strato. Questo sarebbe un movimento di diffusione per legge di Fick. La membrana possiede anche proteine integrali che partecipano al trasporto di acqua chiamate aquaporin. Le aquaporine sono come tubi pieni d'acqua che attraversano la membrana. Attraverso questo canale viene realizzato un flusso di massa submicroscopico soggetto all'equazione di Poiseuille. Questi due percorsi sono riportati di seguito.

48
Osmosis is diffusion of water
Water is very important to life Diffusion of water from high concentration of water to low concentration of water across a semi-permeable membrane 48

50
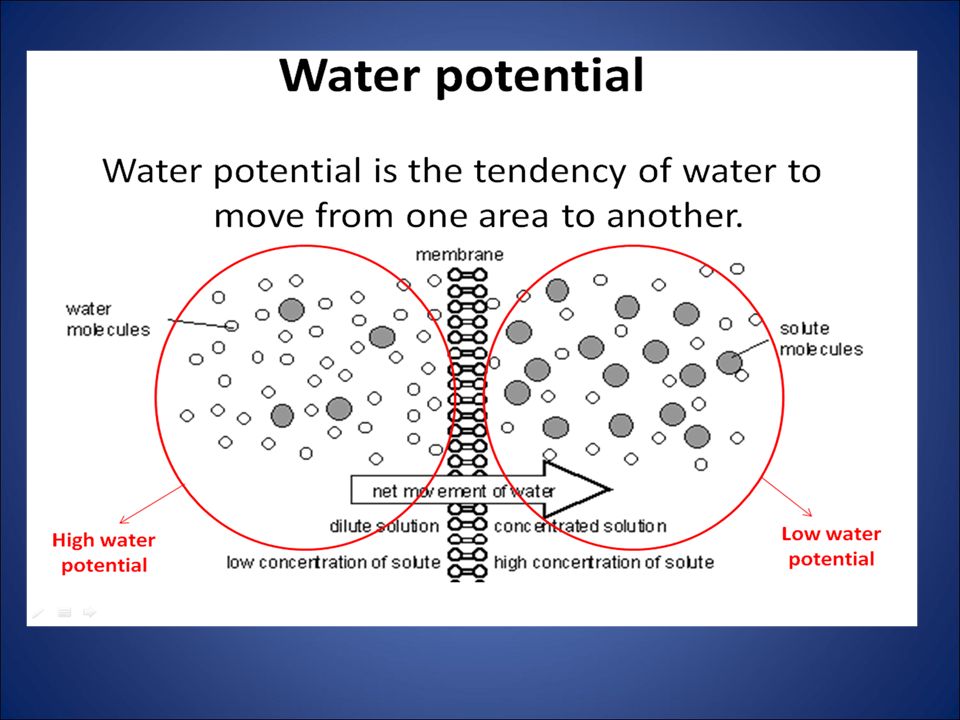
Concentration of water
Direction of osmosis is determined by comparing total solute concentrations Hypertonic - more solute, less water Hypotonic - less solute, more water Isotonic - equal solute, equal water hypotonic hypertonic water net movement of water 50

52
Aquaporins 1991 | 2003 Water moves rapidly into & out of cells
evidence that there were water channels Peter Agre John Hopkins Roderick MacKinnon Rockefeller 52

53
Aquaporins An you can see aquaporins have been well studied. In a. you can see the pore…it is just the size of an 18 molecular weight molecule and water as it goes through in particular orientations brought about with these R groups. Two things make this work, size restriction and electrostatic repulsion (in case a Na+ or other ions bounce in)
")
54
What is an Aquaporin? Transmembrane channel found in all life forms
They facilitate the efficient permeation of water in a bidirectional nature across the cell membrane They have high specificty for water and a select few solutes but are able to exclude proton transfer

55
Aquaporins in Different Life Forms
Human AQP – 10 different genes Bacterial AQP – 2 different genes Plant AQP – over 30 differernt genes Why all the variation in humans and even more so in plants?

56
General Homology of Aquaporins
Despite species diversity, all aquaporins (AQP) exhibit a remarkable similarity in their general composition This suggests a common evolutionary ancestor amoung the aquaporin family
 exhibit a remarkable similarity in their general composition. This suggests a common evolutionary ancestor amoung the aquaporin family.")
57
AQP Conserved Structural Elements
NPA motif Asn-Pro-Ala Restriction Pore H2, H5, LE1, LE2

58
Bacterial Aquaporins – A Model System
There are two classes of bacterial AQP AQPZ – an explicit water channel very similar to human AQP1 which is also an expilict water channel GlpF – channel capable of conduction glycerol aquaglycoporins What makes bacteria a model system? What constitutes the differences between these two channels?

59
Bacterial GlpF NPA motif is consistant within all AQP Restriction Pore
vital to the exclusion of protons transport Restriction Pore Consists of a tetrad of residues from the H2, H5, LE1, and LE2 regions constituents vital to glycerol conduction

60
Role of the NPA Motif binds water molecule in center of pore
similar orientation of hydroxyl dipoles in glycerol initiates bipolar arrangement of molecules above & below bound region favorable for the conduction of solute unfavorable for protons

61
NPA Positioning and Location
The NPA motif is held in place with its two Asparagine residues pointed toward the center of the channel by four strong hydrogens bonds

62
Role of the Restriction Pore
Suprise! It restricts the entry of molecules primarily based on size A secondary mechanism entails exclusion based on the charges of potential transfer molecules Both of these mechanisms are dictated by the amino acid composition of the pore

63
Restriction Pore, con't Atoms: H2 – Trp48 H5 – Gly191 LE1 – Phe200
LE2 - Arg206 Water and glycerol bound in the pore Increased hydrophobicity Phe200/Trp48 Smaller Pore Size Gly191

64
Glycerol & Water – How does it Happen?
glycerol and water are not transported to fulfill any stoiciometric requirements within the cell the glycerol moves down the channel by competing with water for hydrogen bonds

65
Glycerol is Much Larger Than Water
The z-axis of aquaporins is very narrow which also helps to limit “proton hopping” along with the bipolar orientation of dipole moments This narrowness makes glycerol conduction difficult GlpF contains a periplasmic vestibule located between the restriction pore and NPA motif allows glycerol molecules to orient themselves for entry into the pore this same vestibule is not present in AQPZ

66
Glycerol & Water con't There are loops which constitue the vestibule formed in GlpF arising from its asymmetric nature these sames loops are much shorter in AQPZ leading to no vestibule and consequently no glycerol conduction the vestibule in GlpF has a -4kcal/mol energy well in the proton motive force (PMF)
")
67
Human Aquaporins

69
La segnalazione intercellulare
L’evoluzione degli organismi multicellulari dipende dalla capacità delle cellule di comunicare una con l’altra. La comunicazione intercellulare regola lo sviluppo e l’organizzazione dei tessuti, controlla la crescita e la divisione cellulare, la loro sopravvivenza e coordina le diverse attività cellulari. La comunicazione intercellulare negli animali superiori è complessa, suggerendo che molti geni in questi organismi siano coinvolti nel suo controllo e coordinazione. I segnali che le cellule si inviano devono essere riconosciuti

70
Il recettore è una proteina che ha il compito di riconoscere una sostanza esogena (farmaco) o endogena e provocare, dopo il riconoscimento, una risposta biologica all'interno della cellula. I recettori si trovano sulla membrana o nel citosol della cellula, quindi all'interno della cellula stessa.

71
Esistono recettori specifici per ogni molecola segnale.
Ogni cellula, a seconda del suo stato di differenziamento, possiede uno specifico set di recettori per rispondere ai segnali che sono necessari per le sue funzioni. Le molecole segnale idrofiliche hanno recettori sulla membrana plasmatica, mentre quelli lipofiliche hanno recettori intracellulari

72
L’interazione tra le molecole segnale ed i loro recettori impone cambiamenti conformazionali ai recettori che subiscono transizioni allosteriche. L’interazione molecola segnale-recettore scatena una cascata di segnali intracellulari. Segnali intracellulari

73
Molecola segnale – recettore Cambiamento allosterico
Attivazione della segnalazione (ad esempio fosforilazione) Risposte intracellulari (messaggeri secondari) 1) Modificazione del metabolismo 2) Attivazione della trascrizione genica
 Risposte intracellulari (messaggeri secondari) 1) Modificazione del metabolismo. 2) Attivazione della trascrizione genica.")
74
a) aggiunta di un gruppo fosfato b) addizione di un gruppo GTP
La fosforilazione: a) aggiunta di un gruppo fosfato b) addizione di un gruppo GTP Gli enzimi chinasi Le proteine G
 aggiunta di un gruppo fosfato. b) addizione di un gruppo GTP. Gli enzimi chinasi. Le proteine G.")
75
La fosforilazione può avvenire sul gruppo -OH degli aminoacidi serina,
treonina e tirosina

76
I recettori di membrana per molecole idrofiliche
ed i recettori intracellulari per molecole liposolubili

77
I recettori intracellulari degli ormoni steroidei: legano l’ormone
nel citoplasma, subiscono una transizione allosterica e si spostano nel nucleo dove mediano la trascrizione genica.

Presentazioni simili











>")











