Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La maggior parte delle fermentazioni industriali sono processi aerobi e quindi, richiedono l’apporto di ossigeno. La maggior parte delle fermentazioni industriali avviene in coltura sommersa. La richiesta di O 2 è soddisfatta attraverso l’ aerazione e l’agitazione della brodocoltura. Aerazione 1. Quant’è l’O 2 necessario? 2. Qual è la [O 2 ] necessaria? Poiché l’ O 2 è poco solubile nelle soluzioni acquose, non è possibile rifornire in una volta sola la brodocoltura di tutto l’O 2 necessario 3. Come si trasferisce l’O 2 dalla fase gassosa alla fase liquida e quindi alla cellula?

2
Rifornimento di ossigeno su scala di laboratorio : su scala industriale e pilota: mediante scuotimento mediante agitazione orbitante mediante un sistema di aerazione

3
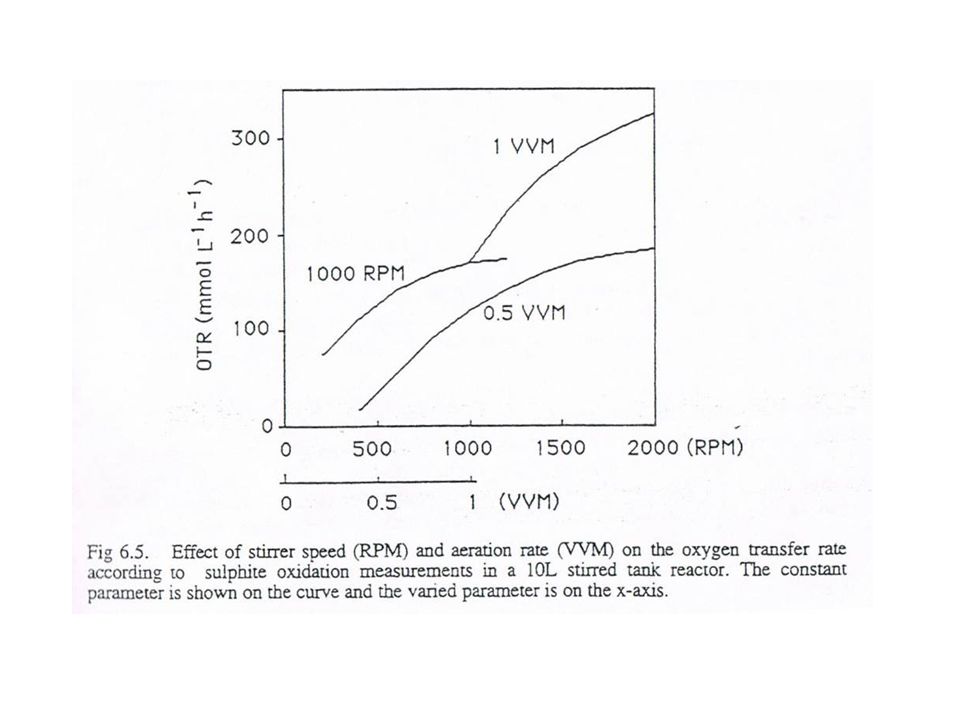
OTR max = 100 - 400 mmol O 2 l -1 h -1

4
Ruolo dell’ossigeno nel metabolismo Accettore terminale di elettroni durante il processo di ossidazione Substrato per reazione del metabolismo primario e secondario: Introduzione di O nei composti organici ad opera di mono- e diossigenasi: -catabolismo degli idrocarburi alifatici e aromatici -trasformazione degli antibiotici -reazioni di sintesi degli acidi grassi mono- e polinsaturi di membrana

5
1a. Richiesta di O 2 Alcuni esempi di fonti di C ed energia: 192 g O 2 per 180 g glucosio 1.07 g O 2 per g glucosio 64 g O 2 per 60 g acido acetico 1.07 g O 2 per g glucosio 352g O 2 per 114 g eptano 3.09 g O 2 per g eptano La stechiometria della reazione relativa all’ossidazione del substrato rappresenta una stima approssimativa dell’O 2 richiesto, poiché non tiene conto della frazione di substrato convertita in biomassa

6
Conoscenza della stechiometria della crescita e della formazione del prodotto Fonte di C ed energia + Fonte di N + Fonti minerali+ O 2 + richieste specifiche* Biomassa+ prodotto + CO 2 + H 2 O + calore Nel caso di un chemioorganotrofo aerobio : * Se auxotrofo aCxHyOz + bNuHwOw + Sali + cO 2 dC H O N + eCmHnOpNr + fH 2 O +gCO 2 + H

7
1b. Richiesta di ossigeno Stima Johnson (1964) ha proposto l’equazione: A/Y x,s – B = C C = quantità di O 2 necessaria a convertire il substrato in 1 g di biomassa, cioè a produrre 1 g di biomassa ( = 1/Y x,o ) A = quantità di O 2 necessaria ad ossidare 1 g di substrato a CO 2 e H 2 O Y x,s = resa, g di biomassa prodotta da 1 g di substrato A/Y x,s = quantità di O 2 necessaria ad ossidare la quantità di substrato che dà 1 g di biomassa B = quantità di O 2 necessaria ad ossidare 1 g di biomassa a CO 2, H 2 O, NH 3 (calcolabile se è nota la composizione elementare della biomassa; B,in media = 1.33 g)
 ha proposto l’equazione: A/Y x,s – B = C C = quantità di O 2 necessaria a convertire il substrato in 1 g di biomassa, cioè a produrre 1 g di biomassa ( = 1/Y x,o ) A = quantità di O 2 necessaria ad ossidare 1 g di substrato a CO 2 e H 2 O Y x,s = resa, g di biomassa prodotta da 1 g di substrato A/Y x,s = quantità di O 2 necessaria ad ossidare la quantità di substrato che dà 1 g di biomassa B = quantità di O 2 necessaria ad ossidare 1 g di biomassa a CO 2, H 2 O, NH 3 (calcolabile se è nota la composizione elementare della biomassa; B,in media = 1.33 g).")
8
1c. Richiesta di O 2 Stima Esempio: nel caso in cui il substrato è C 6 H 12 O 6, si ha: A= 1.07 g considerando un valore di resa respiratoria pari a: Y x,s = 0.5 e, considerando: B = 1.33 g si avrà: C = 0.8 g O 2 Per ottenere valori di densità di biomassa dell’ordine delle decine di g per litro di brodocoltura, occorre una quantità di O 2 dello stesso ordine di grandezza. A causa della bassa solubilità dell’O 2 nelle soluzioni acquose, non è possibile rifornire il mezzo di tutto l’O 2 necessario in una volta sola, ma occorre rinnovare l’O 2 disciolto trasferendolo dalla fase gassosa a quella liquida. C= Y O,X

9
Y x,o Y x,s Y x,c

10
La richiesta di O 2 dipende da: -dalla fonte di C utilizzata e dal tipo di metabolismo -dal grado di riduzione della fonte di C -dalla frazione di substrato convertita in biomassa -dal grado di ossidazione della biomassa (che si può considerare mediamente costante) Nel complesso dalla stechiometria della reazione relativa alla produzione di biomassa a partire da un dato substrato. 1d. Richiesta di O 2

11
La velocità specifica di consumo di O 2 (q O2 ) dipende dalla concentrazione di O 2 secondo l’ eq. di Monod : si chiama concentrazione critica di O 2, [O 2 ] critica, quella concentrazione al di sopra della quale la velocità specifica di consumo di O 2, q O 2, non dipende più dalla concentrazione dell’ossigeno disciolto (Dissolved Oxygen Concentration - DOC). 2a. L’ossigeno come substrato limitante
. 2a. L’ossigeno come substrato limitante.")
12
Non è sufficiente basare la quantità di O 2 da fornire alla coltura solo sulla stima del fabbisogno complessivo, poiché il metabolismo e quindi la crescita dipendono dalla concentrazione dell’O 2 disciolto. Una produzione ottimale di biomassa può essere ottenuta soddisfacendo alla richiesta di O 2 e mantenendo la concentrazione dell’ O 2 disciolto al di sopra del valore critico. Se DOC < [O 2 ] critica il metabolismo risulterà disturbato. 2b. L’ossigeno come substrato limitante T [°C] [O 2 ] crit [mmoli O 2 /dm 3 ] Azotobacter sp. 300.018 Escherichia coli 270.008 Saccharomyces sp. 300.004 Penicillium chrysogenum 240.022

13
2c.Effetto della concentrazione dell’O 2 disciolto sulla velocità di crescita specifica Aerobi Anaerobi facoltativi Anaerobi stretti

14
2d. La solubilità dell’ossigeno dipende da: la pressione parziale dell’O 2 la temperatura la presenza di altri soluti nel mezzo di coltura (la presenza di sali in soluzione fa diminuire la solubilità di O 2 ) T [°C]C O 2 [mg l -1 ]H [mg l -1 atm -1 ] 258.1038.8 356.9933.4 - Se si usa aria come fonte di O 2 - Se si usa ossigeno puro come fonte di O 2
 T [°C]C O 2 [mg l -1 ]H [mg l -1 atm -1 ] Se si usa aria come fonte di O 2 - Se si usa ossigeno puro come fonte di O 2.")
15
2e. A causa della bassa solubilità dell’O 2 in acqua, durante la crescita, in assenza di trasferimento, l’ossigeno sarebbe consumato in brevissimo tempo ! Se conosco C = 1/Y x,o, posso calcolare r O2, essendo noti X e , in quanto: r O2 = q O2 X = X/ Y x,o Esempio: Se : C = 0.8 g O 2 g -1 biomassa, si avrà : Y x,o = 1.25 e, inoltre se: = 0.5 h -1, X = 10 g l -1 Si avrà: r O2 = 4 g l -1 h -1 Poiché la [O 2 ] a 25 °C è pari a 8.1 mg l -1, si avrà: 0.008 g l -1 /4 g l -1 h -1 = 0.002 h = 7.2 s In assenza di trasferimento tutto l’O 2 presente sarebbe consumato in 7.2 s !

16
3a. Trasferimento Una serie di resistenze caratterizzate da costanti di trasferimento K determinano la velocità complessiva di trasferimento dalla bolla d’aria alla cellula (OTR, Oxygen Transfer Rate). Il passaggio più lento e pertanto limitante la velocità di trasferimento complessiva è il primo, poiché la resistenza al trasferimento attraverso la soluzione può essere ridotta dall’agitazione e perché l’uptake di O 2 da parte della cellula è molto rapido Il passaggio di O 2 dall’aria alla cellula microbica (trasporto multistep) si può considerare che avvenga in 3 step principali: -trasferimento dalla bolla d’aria alla soluzione -trasferimento dell’O 2 disciolto in soluzione attraverso il mezzo fino alla cellula -uptake dell’O 2 disciolto da parte della cellula
. Il passaggio più lento e pertanto limitante la velocità di trasferimento complessiva è il primo, poiché la resistenza al trasferimento attraverso la soluzione può essere ridotta dall’agitazione e perché l’uptake di O 2 da parte della cellula è molto rapido Il passaggio di O 2 dall’aria alla cellula microbica (trasporto multistep) si può considerare che avvenga in 3 step principali: -trasferimento dalla bolla d’aria alla soluzione -trasferimento dell’O 2 disciolto in soluzione attraverso il mezzo fino alla cellula -uptake dell’O 2 disciolto da parte della cellula.")
17
3b. Trasferimento Secondo il modello del film liquido stazionario si può ipotizzare che all’interfaccia gas/liquido esista un film liquido stazionario. La diffusione attraverso il film liquido stazionario determina la velocità di trasferimento complessiva dell’ossigeno dalla bolla d’aria alla cellula Si assume che il liquido sia saturato con il gas, cioè che all’interfaccia gas-liquido C = C*

18
In accordo con la legge di Fick, la portata di O 2 attraverso il film liquido stazionario è: A = area interfacciale del film (m 2 ) D f = coefficiente di diffusione di O 2 (m 2 h -1 ) ΔC = gradiente di concentrazione attraverso il film (moli m -3 ) δ= spessore del film (m) δ non è noto e si combina con D f nel termine K L (coefficiente di trasferimento di massa), per cui: per unità di volume di coltura (V L ) e ponendo: 3c. Trasferimento ovvero : si ha: (mol h -1 ) (mol m -3 h -1 ) (m 2 m -3 )
 (mol m -3 h -1 ) (m 2 m -3 ).")
19
3d. Capacità massima di trasferimento dell’ossigeno Il fermentatore deve essere capace di fronteggiare la richiesta di ossigeno da parte della brodocoltura, cioè la velocità massima di trasferimento di ossigeno deve essere uguale o superiore alla velocità di consumo dell’ossigeno da parte del microrganismo. Poiché: OTR è massima se C = 0, per cui:

20
3e. Coefficiente di trasferimento volumetrico: K L a [h -1 ] - Rappresenta la misura della capacità di aerazione del fermentatore, tanto maggiore quanto maggiore è il K L a - Se il K L a è piccolo, la concentrazione di O 2 disciolto scende sotto il livello critico e il fabbisogno del microrganismo non sarà soddisfatto. Quindi il K L a di un fermentatore dovrebbe essere tale da garantire, nel corso della fermentazione, una concentrazione di O 2 disciolto superiore (o uguale) a quella critica. -La conoscenza del K L a di un fermentatore è essenziale per stabilire l’efficienza di aerazione in determinate condizioni operative
 a quella critica. -La conoscenza del K L a di un fermentatore è essenziale per stabilire l’efficienza di aerazione in determinate condizioni operative.")
21
3f. Fattori che influenzano il K L a Temperatura (un aumento di T aumenta K L, ma riduce la solubilità dell’ossigeno) Velocità del flusso d’aria (aumentando il flusso d’aria aumenta «a», ma effetto modesto sul K L a) Proprietà reologiche della brodocoltura (un’elevata viscosità del mezzo dovuta a materie prime complesse e, soprattutto, alla presenza del micelio, aumenta sia che la coalescenza delle bolle riducendo «a» e di conseguenza K L a significativamente) Presenza di agenti antischiuma (aumentano la coalescenza delle bolle riducendo «a» e di conseguenza K L a) Grado di agitazione
 Velocità del flusso d’aria (aumentando il flusso d’aria aumenta «a», ma effetto modesto sul K L a) Proprietà reologiche della brodocoltura (un’elevata viscosità del mezzo dovuta a materie prime complesse e, soprattutto, alla presenza del micelio, aumenta sia che la coalescenza delle bolle riducendo «a» e di conseguenza K L a significativamente) Presenza di agenti antischiuma (aumentano la coalescenza delle bolle riducendo «a» e di conseguenza K L a) Grado di agitazione.")
22
3g. Influenza dell’agitazione sul K L a L’agitazione ha un effetto positivo sul K L a, perché: -disperde l’aria in bolle di piccole dimensioni con aumento quindi di «a» -ostacola la fuoriuscita di bolle dal liquido -impedisce/riduce la coalescenza tra bolle di gas - riduce lo spessore del film liquido all’interfaccia gas-liquido

25
900 rpm750 rpm Acqua Brodo + antischiuma 500 rpm200 rpm Il sistema di agitazione/aerazione e il mezzo di coltura

26
Misurazione dell’ossigeno disciolto MISURAZIONE DIRETTA determinando la sua concentrazione in soluzione (mediante metodi chimici) determinando la tensione dell’ossigeno disciolto (DOT: Dissolved Oxygen Tension) Sensore di Clark MISURAZIONE INDIRETTA
 determinando la tensione dell’ossigeno disciolto (DOT: Dissolved Oxygen Tension) Sensore di Clark MISURAZIONE INDIRETTA")
27
Il sensore di Clark è un dispositivo elettrochimico che permette di misurare per via amperometrica la concentrazione di ossigeno disciolto nelle acque o nei fluidi biologici. Ideato da Leland Clark, venne brevettato nel 1956. Quando è immerso nell'analita, l'O 2 diffonde attraverso la membrana e giunto al catodo viene ridotto, le reazioni sono: O 2 + 4e − + 2H 2 O → 4OH − CATODO 2Ag + 2Cl − → 2AgCl + 2e − ANODO REAZIONE GLOBALE O 2 + 4e − + 2H 2 O + 4Ag + 4Cl − → 4OH − + 4AgCl + 4e − La misura della corrente prodotta dalla reazione di ossidoriduzione è direttamente proporzionale alla pressione parziale di ossigeno che diffonde attraverso la membrana. L'elettrodo viene tarato con soluzioni a concentrazione nota di ossigeno disciolto. Sensore di Clark Sensore di Clark: elettrodi di Pt (A) e Ag/AgCl (B) soluzione di KCl (C) membrana (D) anello di gomma (E) potenziostato (F) galvanometro (G)
 e Ag/AgCl (B) soluzione di KCl (C) membrana (D) anello di gomma (E) potenziostato (F) galvanometro (G).")
Presentazioni simili









1 VELOCITA DI REAZIONE ED EQUILIBRI.>")










