Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
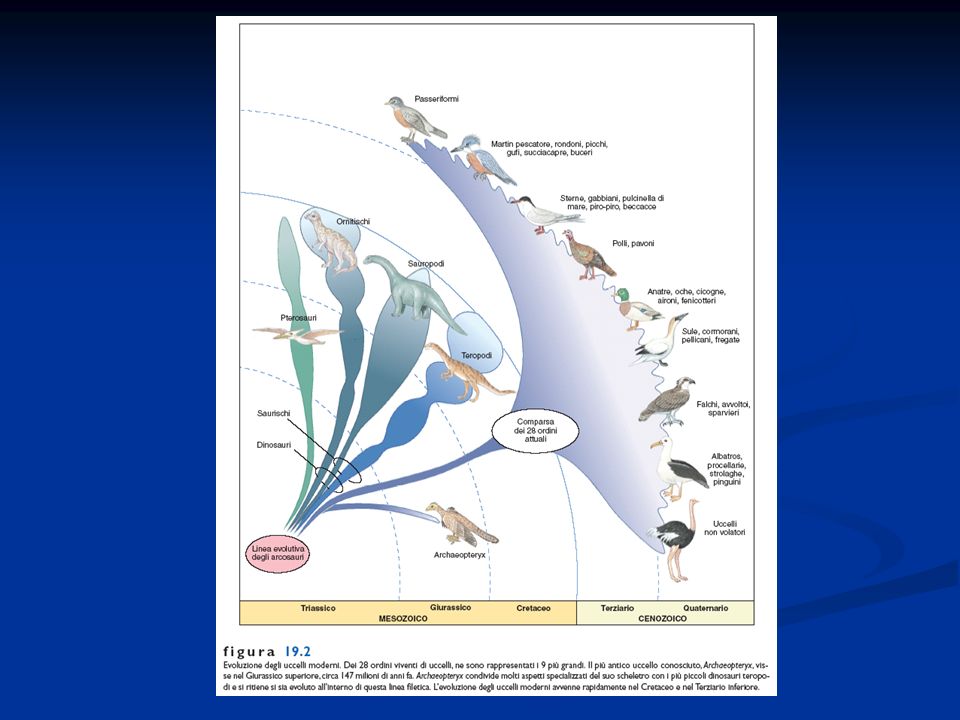
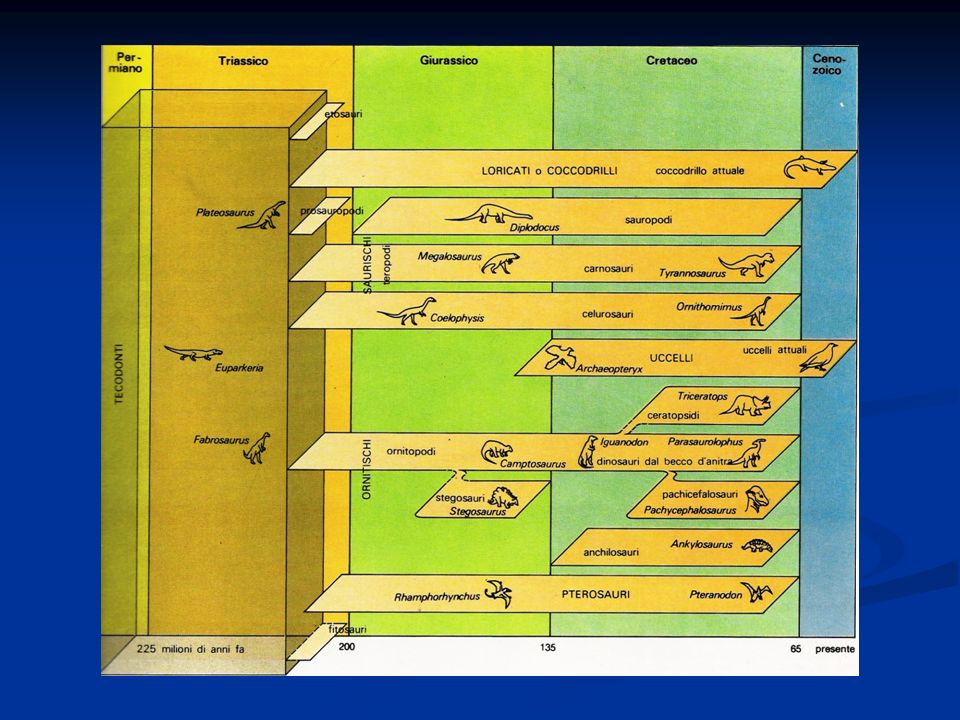
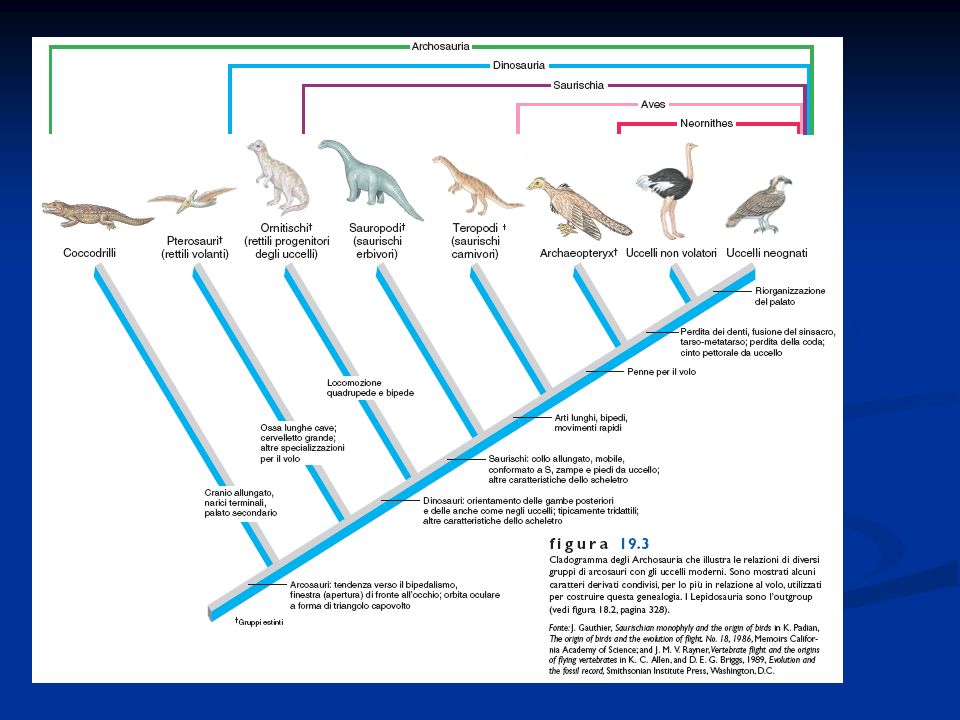
I Teropodi e l’origine e la diversificazione degli uccelli

4
La nuova specie, descritta sull'ultimo numero di Science, è stata rinvenuta nella Formazione di Ischigualasto in Argentina in uno strato datato circa 230 milioni di anni fa ed è stata battezzata Eodromaeus, che significa 'corridore dell'alba'. 'Corridore' in quanto era un predatore che, quindi, inseguiva le sue prede, 'dell'alba' dal momento visse in un periodo molto precoce dell'evoluzione di questi grandi dinosauri carnivori. Diversamente dai sui discententi, Eodromaeus non superava i due metri di lunghezza e i sette chilogrammi di peso, ma aveva alcune caratteristiche da tipico cacciatore, come le potenti zampe posteriori con cui inseguiva le prede e i denti aguzzi con cui sferrava loro il colpo di grazia

10
FIGURA 3. 39 Uccelli estinti
FIGURA 3.39 Uccelli estinti. (a) Ichthyornis, delle dimensioni di un piccione, si alimentava probabilmente di pesci e visse nel Nordamerica circa 100 milioni di anni fa. (b) Diatryma, di circa 60 milioni di anni fa, era un uccello inetto al volo che raggiungeva un’altezza di 2 metri e probabilmente utilizzava la corsa veloce per inseguire piccole prede. (c) Phororhacos, un altro predatore inetto al volo, che visse in Sudamerica circa 30 milioni di anni fa. Secondo Peterson.
 Ichthyornis, delle dimensioni di un piccione, si alimentava probabilmente di pesci e visse nel Nordamerica circa 100 milioni di anni fa. (b) Diatryma, di circa 60 milioni di anni fa, era un uccello inetto al volo che raggiungeva un’altezza di 2 metri e probabilmente utilizzava la corsa veloce per inseguire piccole prede. (c) Phororhacos, un altro predatore inetto al volo, che visse in Sudamerica circa 30 milioni di anni fa. Secondo Peterson.")
12
Xiaotingia zhengi

13
Uno straordinario ritrovamento fossile mette in crisi uno dei miti della paleontologia, quello dell'Archaeopteryx, considerato da sempre il più antico uccello finora noto. Questo organismo fu considerato fin dal momento della sua scoperta, solo due anni dopo la pubblicazione de "L'origine delle specie", un chiarissimo esempio di forma di transizione, un'istantanea sull'origine degli uccelli da progenitori dinosauri. Ma in seguito alla descrizione, sulla prestigiosa rivista Nature, di Xiaotingia zhengi, il mito dell'Archaeopteryx vacilla. Questo nuovo fossile, risalente a circa 155 milioni di anni fa (mentre Archaeopteryx è di alcuni milioni di anni più recente), presenta numerose caratteristiche in comune con Archaeopteryx ; Xiaotingia ha le dimensioni di un pollo, presenta un grosso artiglio sul secondo dito del piede, ma soprattutto possiede un corpo ricoperto di piume e due robusti arti superiori che lo rendevano un abile planatore. . Xiaotingia potrebbe essere anch'esso un uccello primoridiale, un secondo esempio di transizione tra i dinosauri e gli uccelli, di poco precedente ad Archaeopteryx. Ma è dall'approfondita analisi filogenetica che il mito comincia a vacillare: sulla base del confronto di numerosi tratti fenotipici di uccelli e dinosauri piumati, emerge infatti come siaArchaeopteryx che Xiaotingia condividano più tratti in comune con il secondo gruppo che con il primo. In particolare, entrambi apparterrebbero al gruppo dei Deinonychosauria, dinosauri piumati carnivori, piuttosto che a quello degli Avialae, il clade alla base della linea evolutiva che ha dato origine agli uccelli attuali. Il famoso progenitore degli uccelli viene dunque relegato a semplice dinosauro piumato ma, come conclude PZ Meyers sul suo blog Pharyngula, "non preoccupatevi, Archaeopteryx rappresenta ancora un meraviglioso esempio di forma di transizione. Questo nuovo fossile è solo un'altra forma di transizione scoperta. I creazionisti non possono quindi consolarsi: Archaeopteryx non è scomparso improvvisamente, è solo diventato parte di un più complesso disegno sull'evoluzione degli uccelli".
, presenta numerose caratteristiche in comune con Archaeopteryx ; Xiaotingia ha le dimensioni di un pollo, presenta un grosso artiglio sul secondo dito del piede, ma soprattutto possiede un corpo ricoperto di piume e due robusti arti superiori che lo rendevano un abile planatore. . Xiaotingia potrebbe essere anch esso un uccello primoridiale, un secondo esempio di transizione tra i dinosauri e gli uccelli, di poco precedente ad Archaeopteryx. Ma è dall approfondita analisi filogenetica che il mito comincia a vacillare: sulla base del confronto di numerosi tratti fenotipici di uccelli e dinosauri piumati, emerge infatti come siaArchaeopteryx che Xiaotingia condividano più tratti in comune con il secondo gruppo che con il primo. In particolare, entrambi apparterrebbero al gruppo dei Deinonychosauria, dinosauri piumati carnivori, piuttosto che a quello degli Avialae, il clade alla base della linea evolutiva che ha dato origine agli uccelli attuali. Il famoso progenitore degli uccelli viene dunque relegato a semplice dinosauro piumato ma, come conclude PZ Meyers sul suo blog Pharyngula, non preoccupatevi, Archaeopteryx rappresenta ancora un meraviglioso esempio di forma di transizione. Questo nuovo fossile è solo un altra forma di transizione scoperta. I creazionisti non possono quindi consolarsi: Archaeopteryx non è scomparso improvvisamente, è solo diventato parte di un più complesso disegno sull evoluzione degli uccelli ..")
16
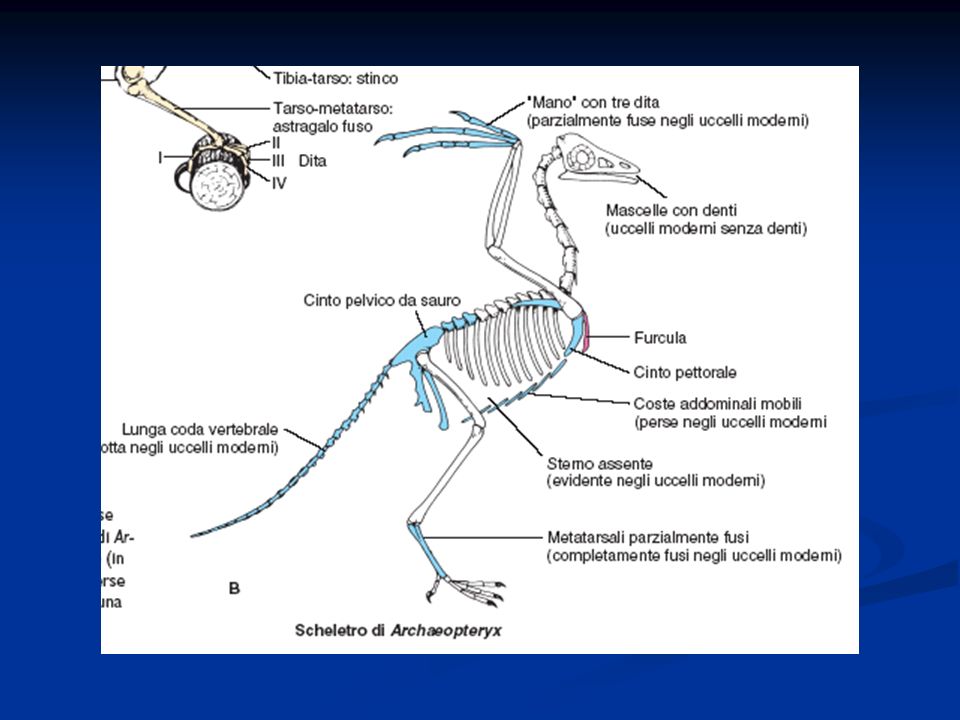
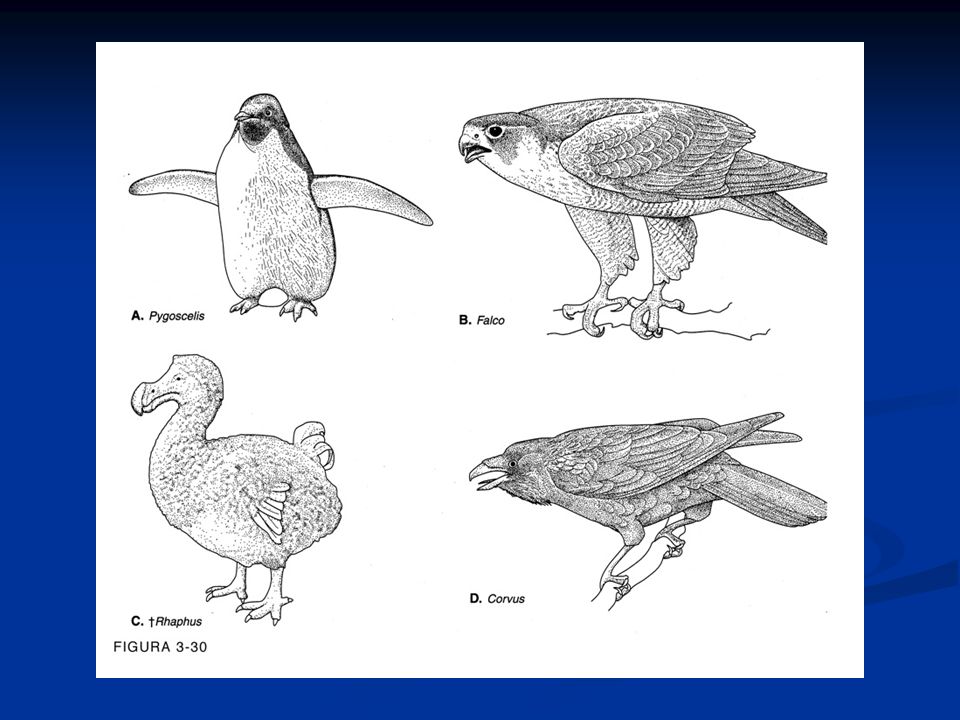
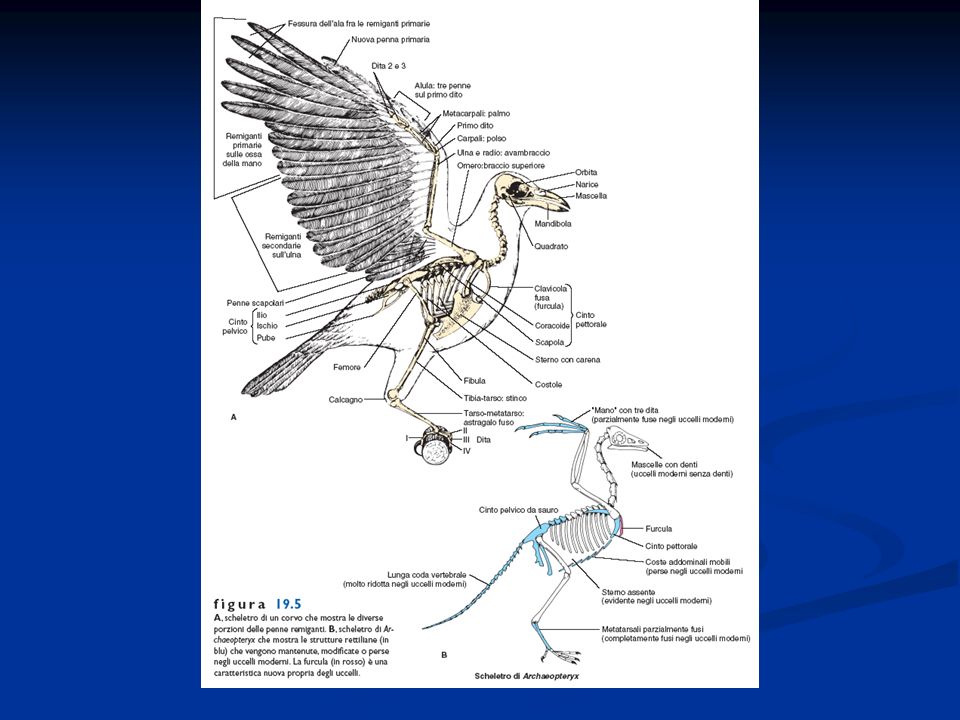
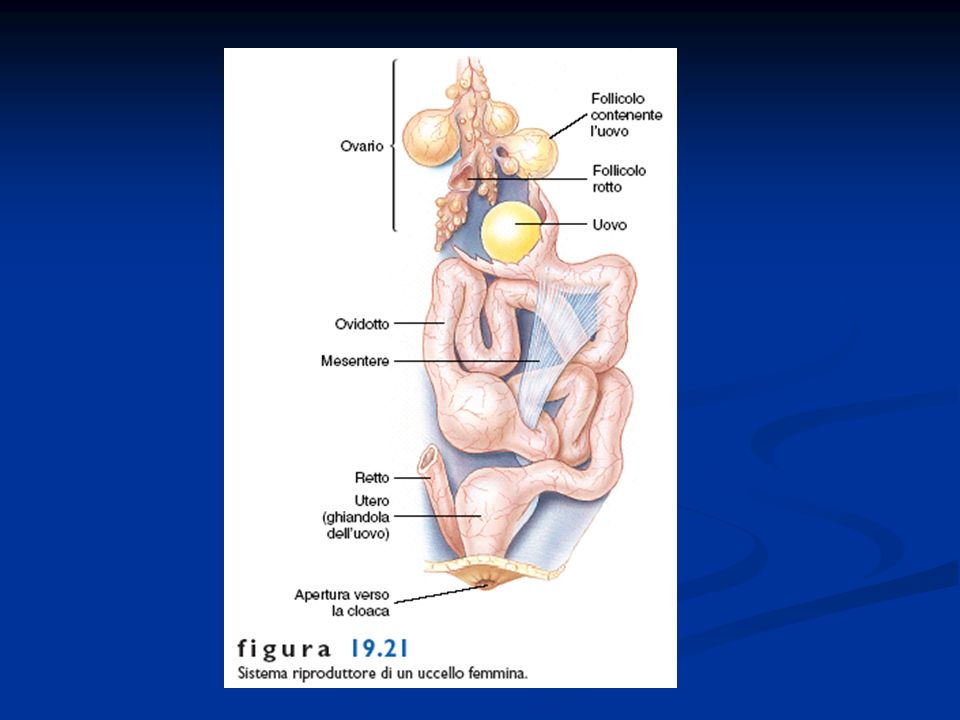
Caratteristiche generali degli Uccelli
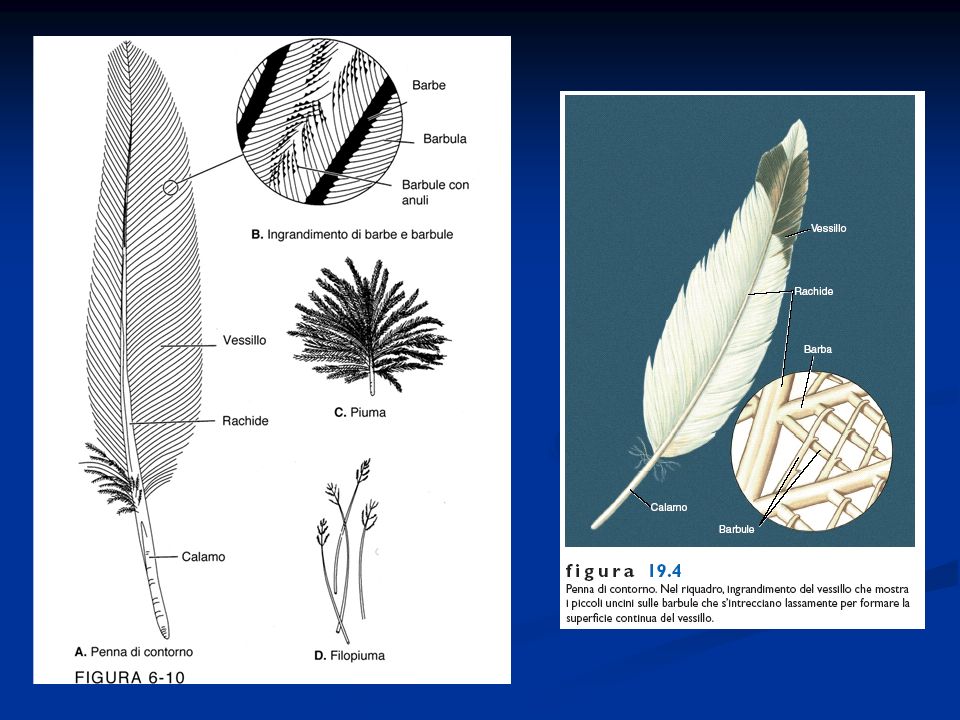
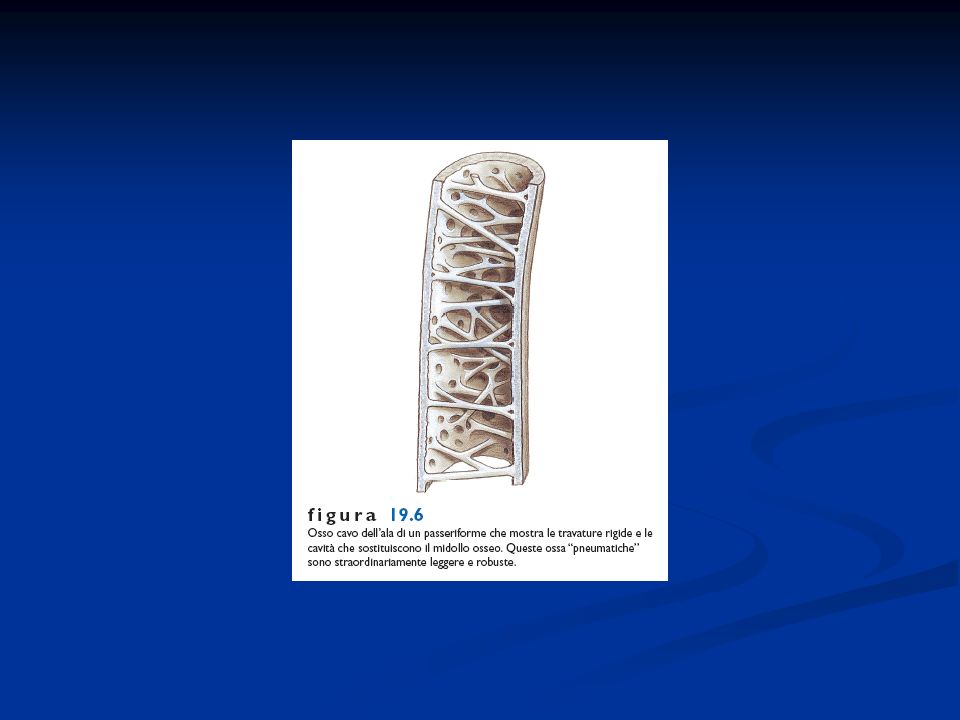
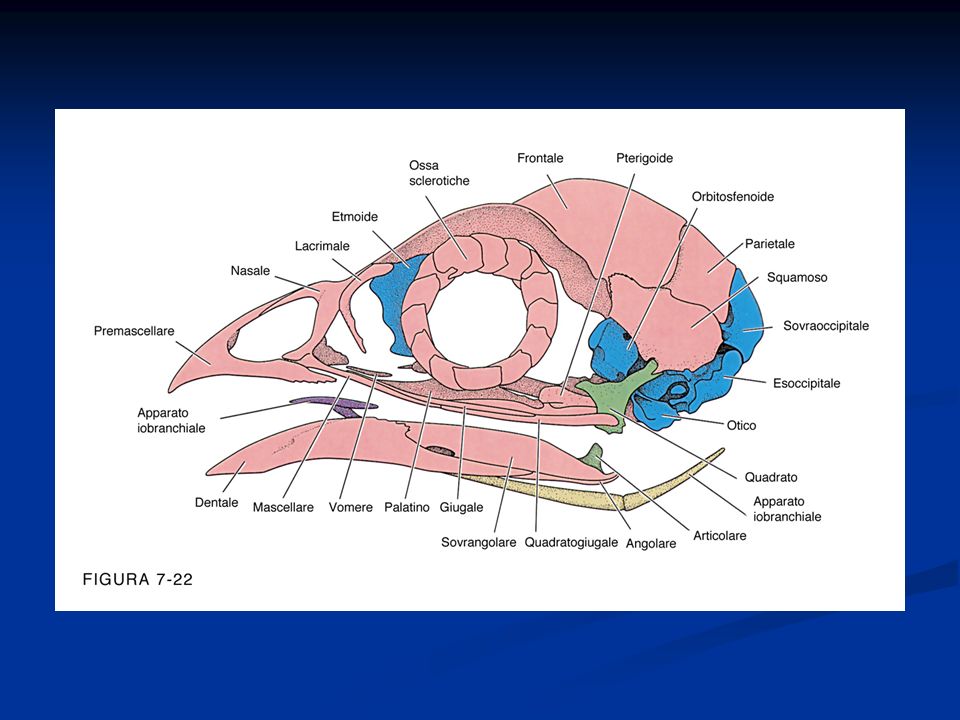
Presenza di penne disposte lungo linee del corpo dette pterilii Residui di squame a livello dell’arto posteriore Presenza di una finestra temporale inferiore. Sono diapsidi modificati per la perdita delle arcate che delimitano le finestre temporali La pelle è priva di ghiandole pluricellulari, tranne che nella regione dell’uropigio Endotermi Adattamento al volo con la conseguente modifica a livello di differenti apparati Ossa pneumatiche collegate alla presenza dei sacchi aeriferi Scompare la doppia volta cranica e la scatola cranica si allarga per la confluenza nel neurocranio di ossa dermiche. Presenza di un unico condilo nella regione occipitale. Potente muscolatura

17
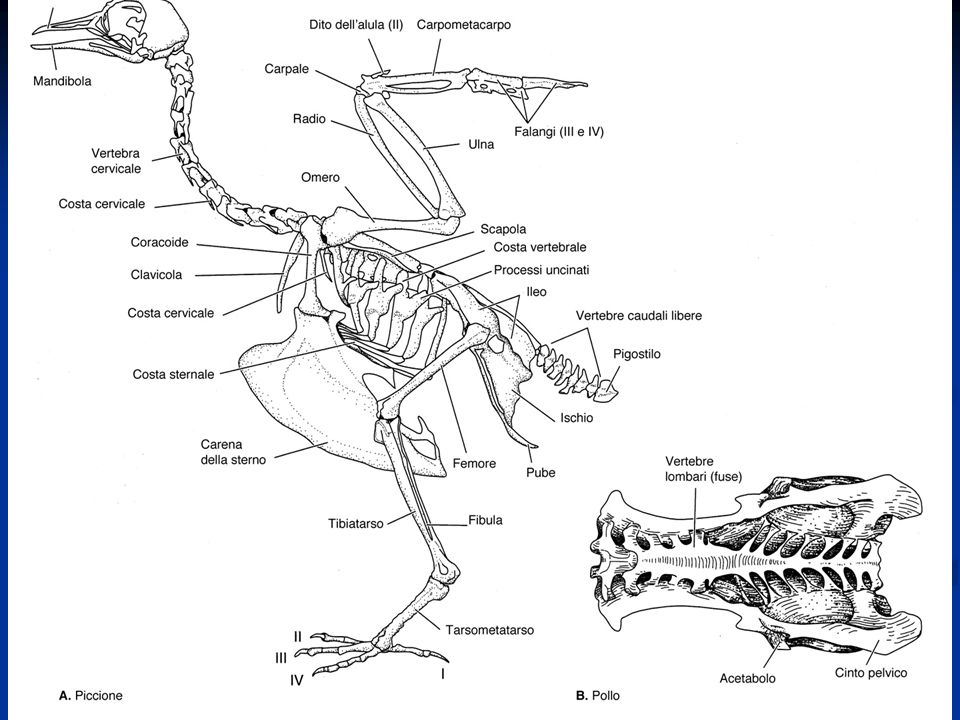
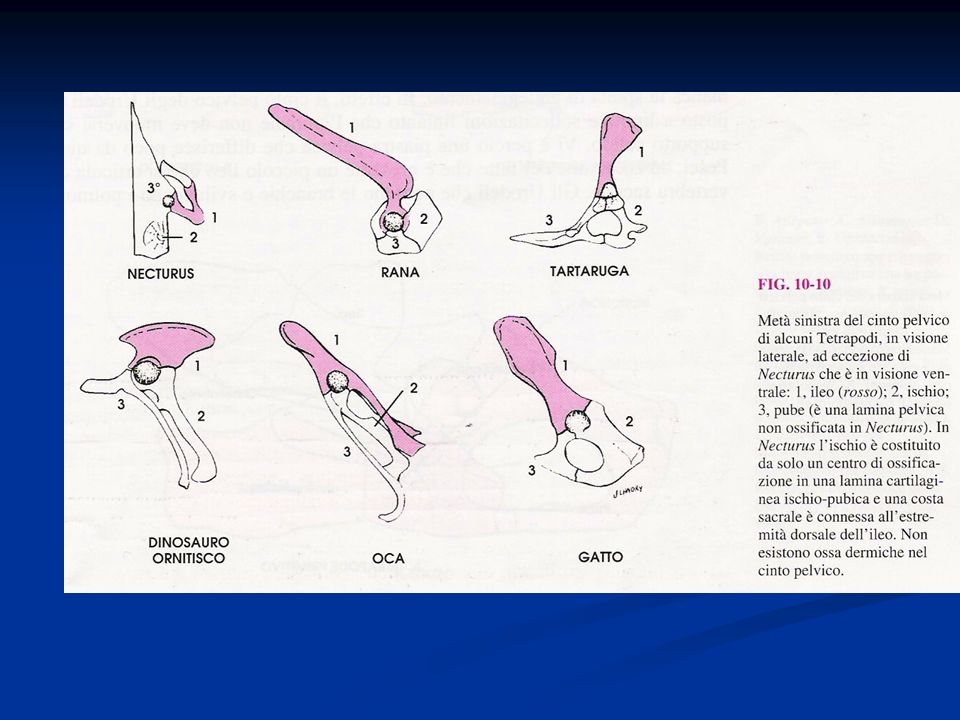
Vertebre con la tendenza alla fusione nella regione sacrale con formazione di un sinsacro
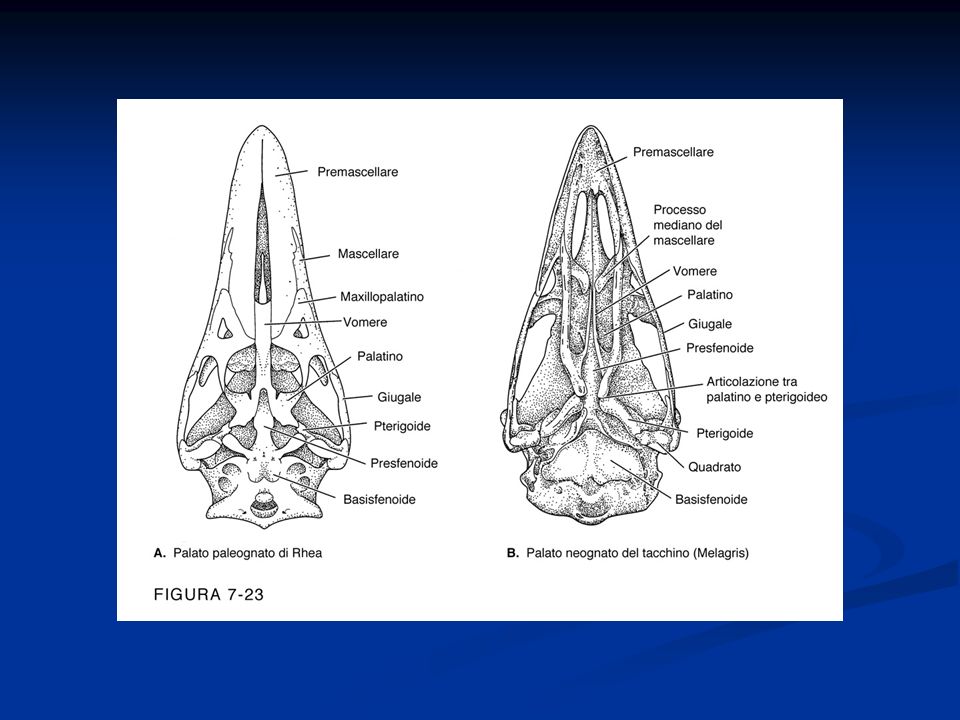
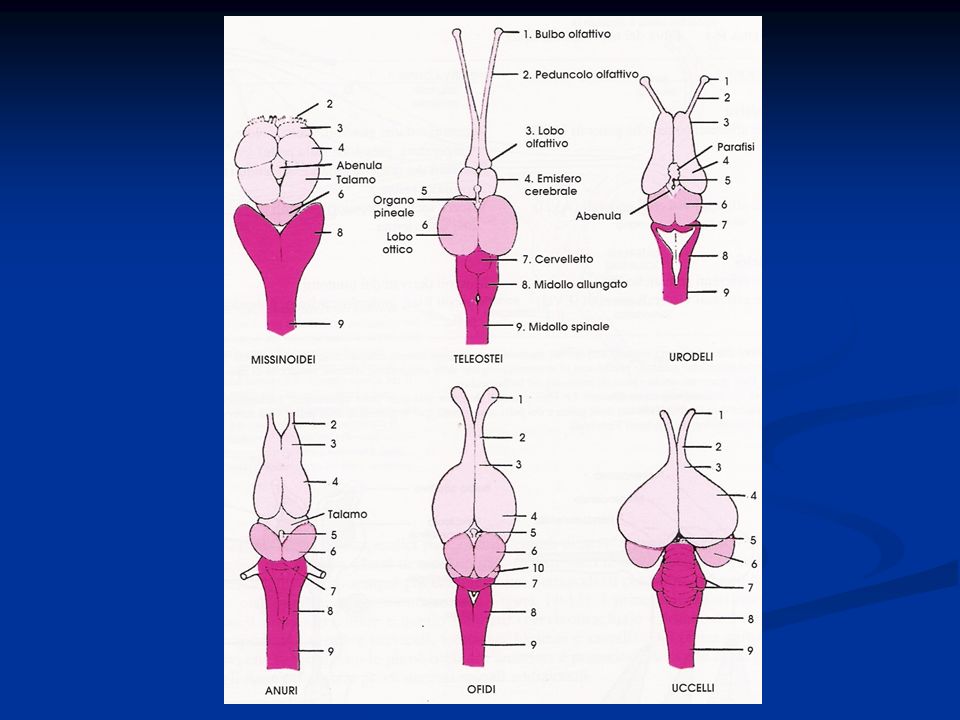
Arto posteriore si presenta modificato per la presenza di una tibia-tarso e di un tarso-metatarso (articolazione intratarsale anziché tra zeugopodio ed autopodio) Riduzione del numero delle dita Presenza della carena Palato differentemente organizzato, per cui gli uccelli sono distinti Paleognati e Neognati. Palato secondario assente Circolazione doppia e completa Sistema digerente modificato per la presenza di un esofago dotato di un gozzo o ingluvie e di uno stomaco modificato per la presenza di un ventriglio Sono uricotelici (eliminano acido urico) senza vescica Sistema respiratorio caratterizzato dalla presenza di sacchi aeriferi che costituiscono una sorta di ghiandole sudoripare Il mesencefalo resta sviluppato per lo sviluppo della vista, ma le attività associative a livello del telencefalo diventano prominenti. Il cervelletto è notevolmente aumentato
 Riduzione del numero delle dita. Presenza della carena. Palato differentemente organizzato, per cui gli uccelli sono distinti Paleognati e Neognati. Palato secondario assente. Circolazione doppia e completa. Sistema digerente modificato per la presenza di un esofago dotato di un gozzo o ingluvie e di uno stomaco modificato per la presenza di un ventriglio. Sono uricotelici (eliminano acido urico) senza vescica. Sistema respiratorio caratterizzato dalla presenza di sacchi aeriferi che costituiscono una sorta di ghiandole sudoripare. Il mesencefalo resta sviluppato per lo sviluppo della vista, ma le attività associative a livello del telencefalo diventano prominenti. Il cervelletto è notevolmente aumentato.")
18
Gli uccelli hanno avuto origine da un taxon di rettili arcosauri saurischi, di modeste dimensioni, carnivori, con un bacino organizzato in un modo simile a quello degli uccelli, vita bipede, arto idoneo a catturare la preda, riduzione dei denti Le forme di uccelli attuali si sono sviluppate a partire dal Cretaceo e la loro fioritura si è verificata nell’era Cenozoica. Sono definiti anche rettili glorificati (Huxley) Gli uccelli sono divisi un due taxa Archeorniti con l’Archeopterix e Neorniti suddivisi in Odontognati (Ichthyornis, Hesperornis), Paleognati o Ratiti (struzzi, emù, casuari, nandù ovvero uccelli corridori) e Neognati , uccelli buoni volatori con sterno carenato
 Gli uccelli sono divisi un due taxa Archeorniti con l’Archeopterix e Neorniti suddivisi in Odontognati (Ichthyornis, Hesperornis), Paleognati o Ratiti (struzzi, emù, casuari, nandù ovvero uccelli corridori) e Neognati , uccelli buoni volatori con sterno carenato.")
23
Cosa serve agli uccelli per volare
Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato alla classe Aves, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il pianocorporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra la classe Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione. Cosa serve agli uccelli per volare? Penne robuste, uno scheletro particolare adatto a sostenere i rigori del volo, sacchi aeriferi in grado di alleggerire il corpo e al tempo stesso sostenere la loro respirazione iperefficiente. Guardando l'anatomia di questi vertebrati volanti si ha in effetti l'illusione che poco non sia stato accuratamente progettato per permettere loro di volare. Ma per sfatare l'illusione è sufficiente guardare ai fossili degli uccelli e dei loro antenati, i dinosauri teropodi, per rendersi conto che il passaggio alla forma attuali è avvenuta senza una vera soluzione di continuità; tanto che non sembra possibile stabilire dove finiscano i dinosauri e dove inizino gli uccelli. Non solo. Guardando al ramo evolutivo che ha portato Agli uccelli, ci si rende facilmente conto di come molte delle loro caratteristiche fisiche, che sembrano 'progettate' per il volo, siano comparse in momenti diversi in differenti antenati non volanti e per rispondere a pressioni selettive ambientali, non legate quindi al volo. Ne è un notevole esempio l'evoluzione di penne e piume, individuate negli anni novanta del secolo scorso in numerosi fossili di dinosauro risalenti al Cretaceo e provenienti prevalente da depositi fossiliferi cinesi. In questi animali, molti dei quali erano di grossa taglia, le penne non potevano certo essere utilizzate per il volo e si sono probabilmente evolute per motivi di isolamento termico o in seguito all’azione della selezione sessuale. Al di là degli esempi aneddotici, la validità generale di un modello evolutivo degli uccelli basato su un lento accumulo di prerequisiti fisici, seguita da un vertiginoso aumento di specie una volta che il piano corporeo di base si era formato, è stata dimostrata con metodi matematici e statistici da Stephen L. Brusatte e colleghi, delle università britanniche e statunitensi di Edimburgo, Oxford e Swarthmore, e del museo di storia naturale di New York. Il lavoro, pubblicato dalla rivista Current Biology, ha esaminato 853 caratteri fisici misurati sui resti di 150 taxa del sottordine dei Theropoda, includendo, oltre agli uccelli e ai loro diretti antenati, anche taxa più vicini a Tyrannosaurus e Velociraptor ed altri rami evolutivi strettamente imparentati, ma distinti, rispetto a quello degli uccelli. Le analisi, che non prende in considerazione informazioni di tipo genetico ma un grande numero di caratteri anatomici, indicano chiaramente che la linea evolutiva che ha portato agli attuali uccelli presenta una velocità di evoluzione molto maggiore rispetto alla media degli altri teropodi. Quindi, se si volesse stabilire un punto di separazione fra Aves e i loro antenati dinosauri, bisognerebbe scegliere il momento in cui l'evoluzione di questo gruppo ha subito una brusca e imprevedibile accelerazione.

26
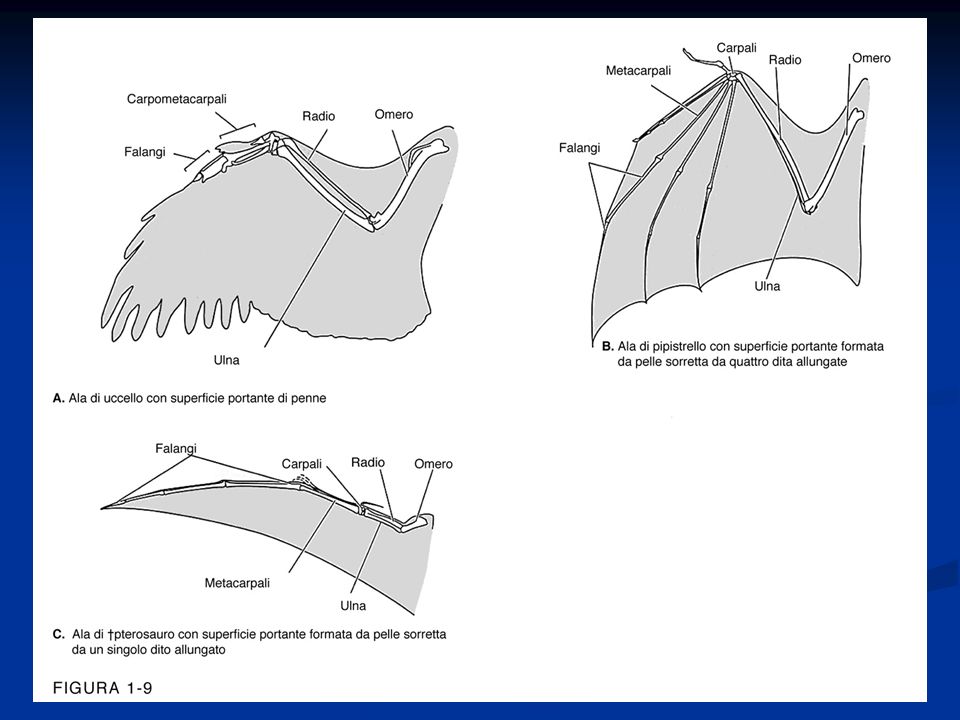
Aspetti degli arti negli Uccelli
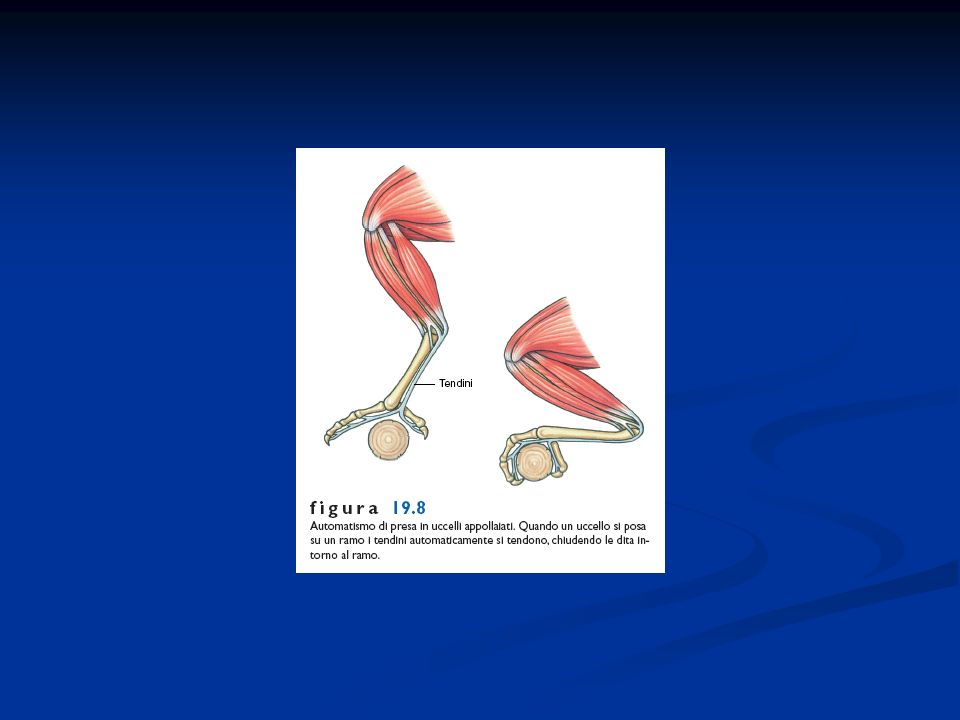
Negli uccelli, come in molti rettili estinti, con la comparsa dell’andatura bipede, si è affermata la disposizione parasagittale dell’arto posteriore. Ciò avviene mediante la rotazione di questo in avanti per cui viene a disporsi sotto il tronco in un piano parasagittale, ciò consente di scaricare parte del peso del corpo sulle strutture scheletriche e di usare più vantaggiosamente i muscoli degli arti per la spinta durante il movimento Negli uccelli l’arto anteriore si modifica in ala, dove è possibile rilevare la presenza di un basipodio modificato

27
Uno studio pubblicato su Evolution (2013) ipotizza che lo sviluppo degli uccelli sia iniziato da un repentino cambio di proporzioni negli arti dei dinosauri. Secondo le conoscenze attuali, gli uccelli hanno avuto origine circa 150 milioni di anni fa da un piccolo gruppo di dinosauri teropodi carnivori chiamati maniraptora. Diverse recenti ricerche hanno mostrato che molti maniraptori avevano un aspetto simile a uccelli, con piume, ossa cave, piccole dimensioni corporee e alti tassi metabolici. Rimaneva però aperta la questione su quando gli arti anteriori si fossero trasformati in ali, rendendo possibile il volo. Hans C.E. Larsson e Alexander T. Dececchi della canadese McGill University, dopo aver esaminato numerosi reperti fossili del periodo, sostengono che durante la maggior parte dell'arco di esistenza dei dinosauri carnivori la lunghezza degli arti sia rimasta di proporzioni relativamente stabili rispetto alle dimensioni del corpo. La proporzione degli arti sarebbe tuttavia mutata improvvisamente: sia gli arti anteriori che posteriori andarono incontro a un drammatico disallineamento rispetto alla dimensione del corpo. Questo cambiamento sarebbe stato essenziale nel permettere ai primi uccelli di volare, e quindi di sfruttare dall'alto il manto forestale. Gli arti anteriori si sarebbero allungati, diventando sufficientemente estesi da servire come profilo alare, mentre gli arti inferiori si sarebbero ristretti, aiutando a rifinire il controllo in volo. I ritrovamenti suggeriscono che gli uccelli siano andati incontro a un improvviso mutamento nel loro meccanismo di sviluppo, tale da permettere ai loro arti anteriori e posteriori di essere soggetti a differenti controllori della lunghezza. Zampe più corte avrebbero permesso di ridurre l'attrito in volo e di muoversi più agilmente sui rami degli alberi. La combinazione di ali migliori con zampe più compatte sarebbe risultata fondamentale per la sopravvivenza degli uccelli in un periodo in cui un altro gruppo di rettili volanti, gli pterosauri, dominava i cieli.

32
Polmone e ventilazione negli uccelli
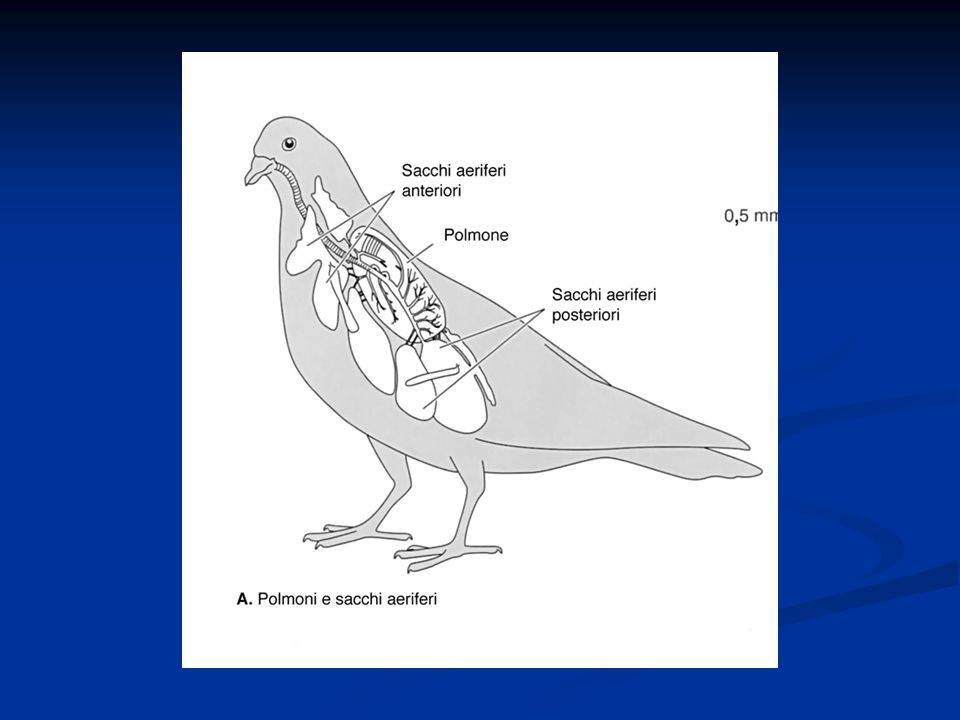
I polmoni soni piccoli, poco elastici e n comunicazione con dei sacchi aerei dotati di parete sottile non respiratoria, distribuiti tra i viscere e all’interno delle ossa cave. Sono presenti inoltre due bronchi principali, uno per ciascun polmone, che dopo aver attraversato il rispettivo polmone sboccano nei sacchi aerei. Sono presenti inoltre dei parabronchi, connessi a una serie di piccole cavità che costituiscono la vera superficie di scambio gassoso

35
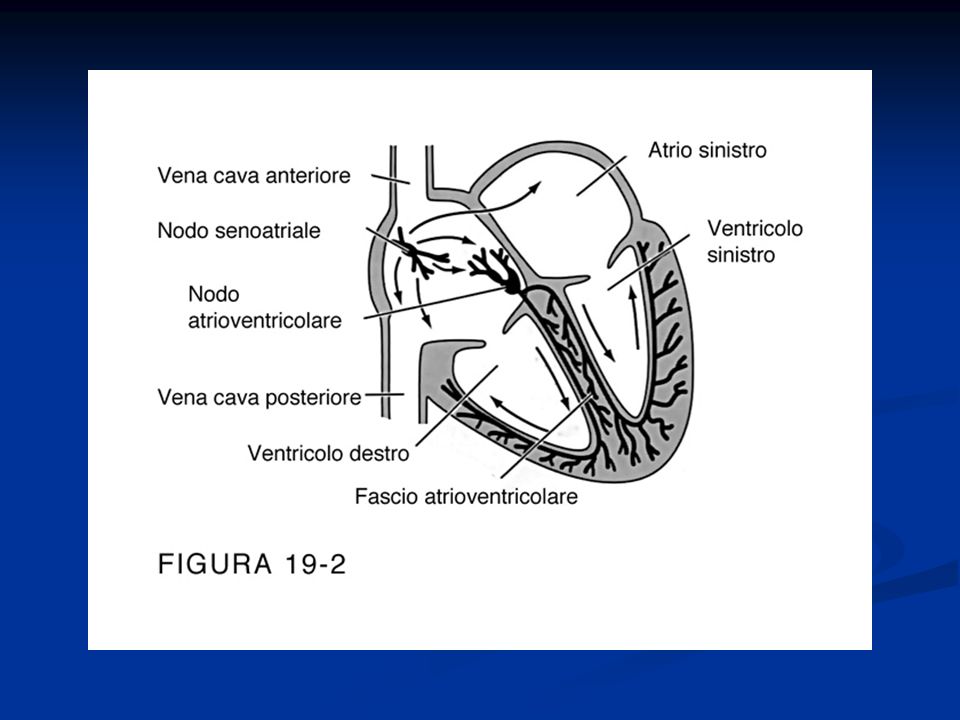
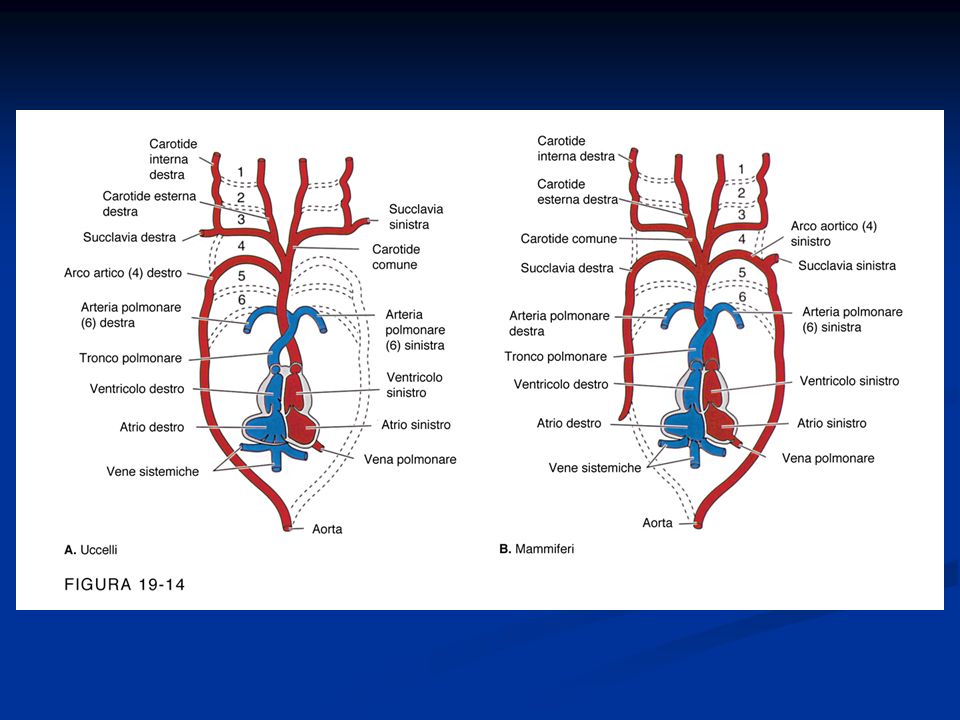
Il cuore di mammiferi ed uccelli è composto da quattro cavità, due atri e due ventricoli, diversamente da quello degli anfibi, che presenta, oltre ai due atri, un solo ventricolo. Nei rettili, invece, la situazione è molto più complessa: i sauri (serpenti e lucertole) presentano tre cavità, i cheloni (le tartarughe) possiedono un setto interventricolare solo parzialmente sviluppato, mentre i loricati (i coccodrilli) hanno un cuore con due ventricoli completi. Questo gruppo di vertebrati è dunque il più indicato per comprendere le basi molecolari dell'evoluzione del cuore a quattro cavità, fondamentale per l'omeostasi termica tipica degli animali a sangue caldo. La divisione in due camere ventricolari consente, infatti, la completa separazione tra circolazione polmonare (piccola circolazione) e circolazione sistemica (grande circolazione), responsabile, insieme al metabolismo elevato e al rivestimento isolante del corpo, della capacità di regolazione della temperatura corporea (omeotermia) di uccelli e mammiferi. Sebbene queste differenze nell'organizzazione cardiaca dei vertebrati siano note da tempo, pochi studi ne hanno indagato le basi molecolari. La rivista Nature pubblica sul suo ultimo fascicolo un interessante studio che ha individuato lo stesso gene coinvolto nella formazione del setto interventricolare nei mammiferi nello sviluppo del cuore di lucertole e tartarughe. Si tratta del gene Tbx5, che codifica per un fattore di trascrizione, già noto per essere coinvolto in alcune patologie cardiache umane. La ricerca ha analizzato l'espressione di questo gene in embrioni di Anolis carolinensis, una lucertola, e Trachemys scripta elegans, una tartaruga. I risultati indicano che i livelli di questa proteina rimangono costanti durante l'intero sviluppo della lucertola, mentre variano nel corso dell'embriogenesi della tartaruga, in modo tale da generare un gradiente di concentrazione longitudinale: nella parte destra del cuore in formazione, infatti, i livelli di espressione rimangono più alti che a sinistra e nel punto di confine di questa differenza concentrazione si forma il setto interventricolare, anche se solo parzialmente. Nella lucertola, al contrario, non si ha alcuna separazione del ventricolo. Per valutare se la diversa espressione del gene Tbx5 sia realmente responsabile della formazione del setto, i ricercatori hanno seguito lo sviluppo del cuore durante l'embriogenesi di alcuni topi geneticamente modificati in modo tale da presentare basse quantità di tale proteina. Come previsto, questi individui non hanno sviluppato la divisione della cavità cardiaca in due ventricoli e, come conseguenza di questa malformazione, hanno manifestato un alto livello di mortalità.
 presentano tre cavità, i cheloni (le tartarughe) possiedono un setto interventricolare solo parzialmente sviluppato, mentre i loricati (i coccodrilli) hanno un cuore con due ventricoli completi. Questo gruppo di vertebrati è dunque il più indicato per comprendere le basi molecolari dell evoluzione del cuore a quattro cavità, fondamentale per l omeostasi termica tipica degli animali a sangue caldo. La divisione in due camere ventricolari consente, infatti, la completa separazione tra circolazione polmonare (piccola circolazione) e circolazione sistemica (grande circolazione), responsabile, insieme al metabolismo elevato e al rivestimento isolante del corpo, della capacità di regolazione della temperatura corporea (omeotermia) di uccelli e mammiferi. Sebbene queste differenze nell organizzazione cardiaca dei vertebrati siano note da tempo, pochi studi ne hanno indagato le basi molecolari. La rivista Nature pubblica sul suo ultimo fascicolo un interessante studio che ha individuato lo stesso gene coinvolto nella formazione del setto interventricolare nei mammiferi nello sviluppo del cuore di lucertole e tartarughe. Si tratta del gene Tbx5, che codifica per un fattore di trascrizione, già noto per essere coinvolto in alcune patologie cardiache umane. La ricerca ha analizzato l espressione di questo gene in embrioni di Anolis carolinensis, una lucertola, e Trachemys scripta elegans, una tartaruga. I risultati indicano che i livelli di questa proteina rimangono costanti durante l intero sviluppo della lucertola, mentre variano nel corso dell embriogenesi della tartaruga, in modo tale da generare un gradiente di concentrazione longitudinale: nella parte destra del cuore in formazione, infatti, i livelli di espressione rimangono più alti che a sinistra e nel punto di confine di questa differenza concentrazione si forma il setto interventricolare, anche se solo parzialmente. Nella lucertola, al contrario, non si ha alcuna separazione del ventricolo. Per valutare se la diversa espressione del gene Tbx5 sia realmente responsabile della formazione del setto, i ricercatori hanno seguito lo sviluppo del cuore durante l embriogenesi di alcuni topi geneticamente modificati in modo tale da presentare basse quantità di tale proteina. Come previsto, questi individui non hanno sviluppato la divisione della cavità cardiaca in due ventricoli e, come conseguenza di questa malformazione, hanno manifestato un alto livello di mortalità..")
Presentazioni simili










 mettere in discussione l’idea del progetto 2) rivalutare il.>")







, specializzato per la distribuzione di: gas.>")







