Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Fotosintesi

2
Organismi fotoautotrofi
Respirazione sole Organismi fotoautotrofi (CH2O)n + nO2 Organismi eterotrofi nCO2 + nH2O Processo spontaneo: richiede composti ricchi di energia libera Tutta la vita sulla terra dipende ultimamente dalla fotosintesi Fotosintesi Questo non significa che le cellule vegetali non compiano la respirazione!!
n + nO2. Organismi eterotrofi. nCO2 + nH2O. Processo spontaneo: richiede composti ricchi di energia libera. Tutta la vita sulla terra dipende ultimamente dalla fotosintesi. Fotosintesi. Questo non significa che le cellule vegetali non compiano la respirazione!!")
3
Le fasi della fotosintesi
2- Trasferimento di elettroni 3- Biochimica 1- Fotochimica e- C ossidato L’ossigeno viene prodotto a partire dall’acqua, NON dalla CO2! C ridotto

4
Photosynthetic bacteria
Chlorobium-green sulfur bacteria Use green pigment chlorophyll Use H2S (hydrogen sulfide), S (sulfur), Na2S2O3 (sodium thiosulfate) and H2 as e- donors. Chromatium-purple sulfur bacteria Use purple carotenoid pigment, same e-donors Rhodospirillum-non sulfur purple bacteria Use H2 and other organic compounds such as isopropanol etc, as e-donors. Reaction: CO2 + 2H2A -----> CH20 + H20 +2A A is not O La fotosintesi è un processo RedOx! Cornelius van Niel e Robert Hill
, S (sulfur), Na2S2O3 (sodium thiosulfate) and H2 as e- donors. Chromatium-purple sulfur bacteria. Use purple carotenoid pigment, same e-donors. Rhodospirillum-non sulfur purple bacteria. Use H2 and other organic compounds such as isopropanol etc, as e-donors. Reaction: CO2 + 2H2A -----> CH20 + H20 +2A. A is not O. La fotosintesi è un processo RedOx! Cornelius van Niel e Robert Hill.")
5
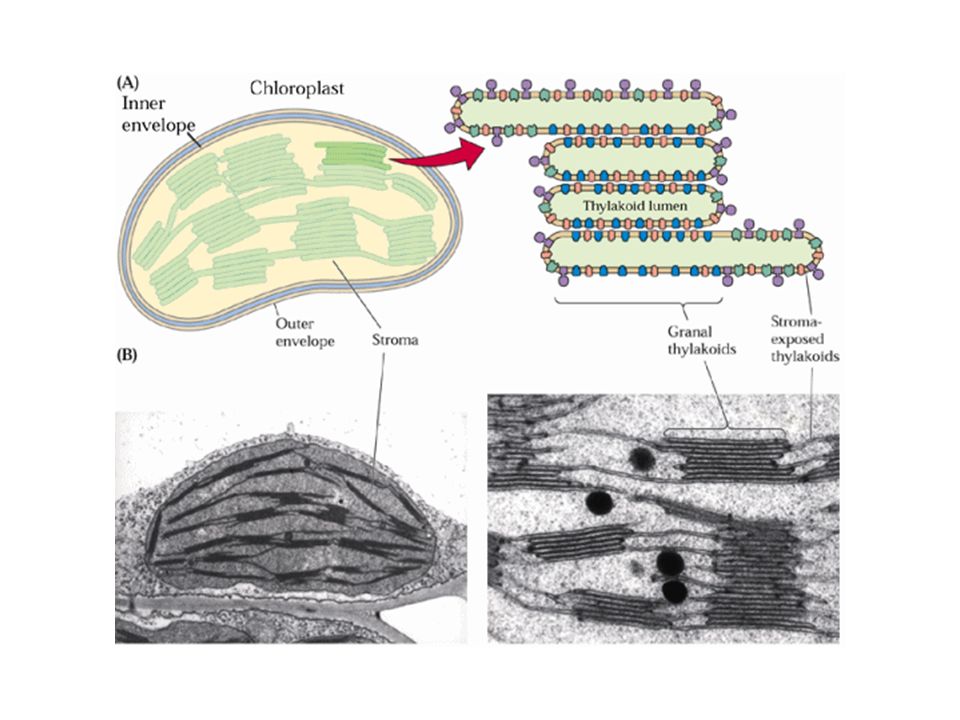
Le reazioni avvengono nel cloroplasto: ultrastruttura
Membrana esterna + interna Lamelle dei grana (pile di tilacoidi) Lamelle stromatiche (tilacoide non appressato)
 Lamelle stromatiche (tilacoide non appressato)")
6
Lamelle stromatiche (tilacoide non appressato)
Lamelle dei grana (tilacoidi appressati)
")
7
Rappresentazione in 3D

8
Lamelle dei grana e stromatiche hanno diversa composizione

9
Fotochimica (assorbimento luce)
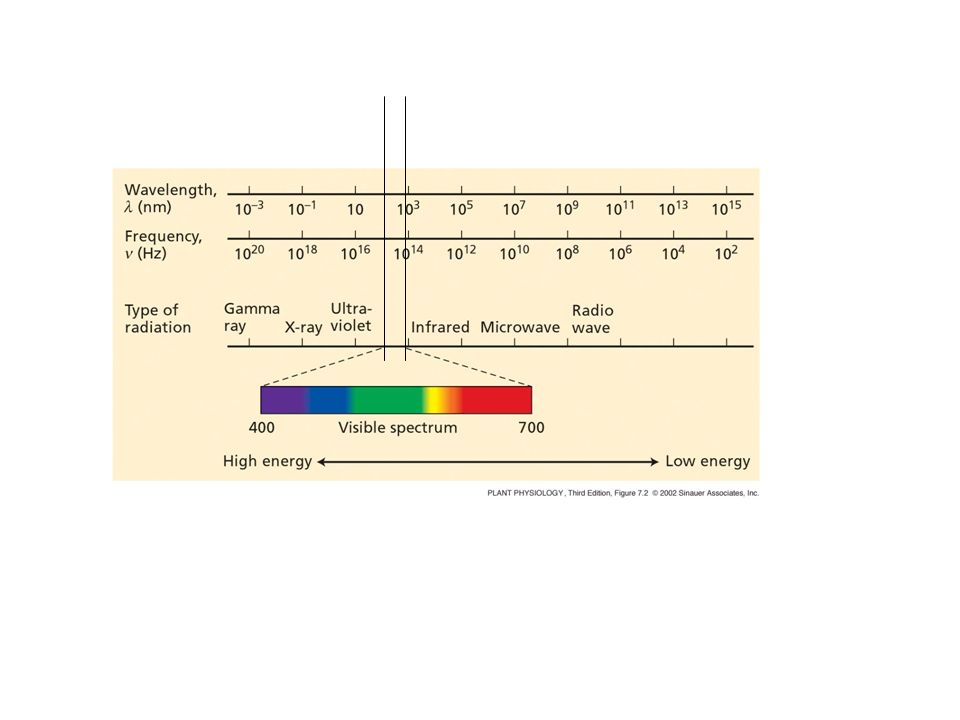
c = = c/ = c/ E = h E = h c/ h= costante di Planck = 6.6 x Js c = 3 x 108 m/s (velocità della luce nel vuoto)
")
10
energia (kJ/mol fotoni) Frequenza (THz)
Lunghezza d’onda (nm) Colore della luce energia (kJ/mol fotoni) Frequenza (THz) 400 violetto 298 700 440 blu 271 650 500 Blu-verde 238 590 600 giallo 199 520 rosso 183 480 rosso lontano 170 Bond Length Energy H--H 74 436 H--C 109 413 C--C 154 348 H--N 101 391 L’energia per rompere un legame (kJ/mol) The amount of energy required to break a bond is called bond energy fotoni con 300 nm (UV) hanno energia sufficiente per rompere i legami
 Colore della luce. energia (kJ/mol fotoni) Frequenza (THz) 400. violetto blu Blu-verde giallo rosso rosso lontano Bond. Length. Energy. H--H H--C C--C H--N L’energia per rompere un legame (kJ/mol) The amount of energy required to break a bond is called bond energy fotoni con 300 nm (UV) hanno energia sufficiente per rompere i legami.")
12
200 400 600 800 1000 1200 1400 Flusso di fotoni Orlo esterno dell’atmosfera , nm superficie della terra Effetto H20, CO2 effetto ozono Assorbimento della clorofilla

13
Molecola N2 ASSORBIMENTO DELLA LUCE DA PARTE DELLE MOLECOLE
Principi della fotochimica -solo la luce assorbita può produrre un cambiamento chimico -ogni fotone assorbito attiva una sola molecola -tutta l’energia del fotone viene trasferita ad un singolo elettrone e ne provoca Io spostamento ad un livello energetico superiore Nella clorofilla gli orbitali π hanno energia inferiore a quelli σ Orbitale atomico Orbitale molecolare di legame Orbitale molecolare di antilegame Molecola N2

14
perché sono delocalizzati su più di un nucleo.
Gli elettroni che assorbono la luce sono gli elettroni di legame di tipo , perché sono delocalizzati su più di un nucleo. L’energia della transizione è più bassa (~luce visibile) Quasi tutte le reazioni fotochimiche e le proprietà spettroscopiche di importanza biologica dipendono dall’assorbimento di fotoni da parte di elettroni . Molecola molecola* * h Il campo elettrico oscillante del fotone genera un dipolo che fa oscillare l’elettrone. L’elettrone riceve l’E del fotone e passa ad un orbitale superiore.
 Quasi tutte le reazioni fotochimiche e le proprietà spettroscopiche di importanza biologica dipendono dall’assorbimento di fotoni da parte di elettroni . Molecola molecola* * h Il campo elettrico oscillante del fotone genera un dipolo che fa oscillare l’elettrone. L’elettrone riceve l’E del fotone e passa ad un orbitale superiore.")
15
energia Stato di base Singoletto (S ,) Stato eccitato (S , *) Tripletto (T , *) S = 0 (2S+1) = 1 S = 0 (2S+1) = 1 S = 1 h/2π (2S+1) = 3 S è il momento angolare dell’elettrone (spin); ha valore di +1/2 o -1/2 h/2π (2S+1) = molteplicità di spin La molteplicità di spin di una molecola può essere: 0 + 1 = 1 singoletto, = 3 tripletto
 = 1. S = 0. (2S+1) = 1. S = 1 h/2π. (2S+1) = 3. S è il momento angolare dell’elettrone (spin); ha valore di +1/2 o -1/2 h/2π. (2S+1) = molteplicità di spin. La molteplicità di spin di una molecola può essere: = 1 singoletto, = 3 tripletto.")
16
Clorofilla PORFIRINA O TETRAPIRROLO La clorofilla è idrofobica. Si trova bene associata a proteine di membrana I cloroplasti contengono un grande quantità di membrane per aumentare la superficie, così da assorbire efficacemente la luce. LEGAME ESTERE ALCOOL FITOLO

17
In questa regione i doppi legami sono DELOCALIZZATI
Gli elettroni di legame sono distribuiti sull’intero sistema coniugato Strutture di risonanza della clorofilla La presenza di molti doppi legami coniugati abbassa l’energia richiesta per la transizione da π a π*

18
Coefficiente di estinzione molare della Clorofilla in metanolo
cm-1/M (λ= )
")
19
Lo spettro d’azione E’ uno spettro, ma non di assorbimento! Descrive l’efficacia di varie nel determinare una risposta biologica specifica Fa capire che non tutte le lunghezze d’onda sono ugualmente efficaci

20
Lo spettro d’azione della fotosintesi e quello di assorbimento della Chl sono molto simili la Chl è il recettore

21
Pigmenti accessori I carotenoidi complementano bene lo spettro di assorbimento della clorofilla (assorbono bene la luce verde) Man mano che aumenta il numero di doppi legami coniugati, la molecola assorbe a lunghezze d’onda visibili Tetrapirrolo aperto

22
Chl b carotene Chl a Ficoeritrobilina

23
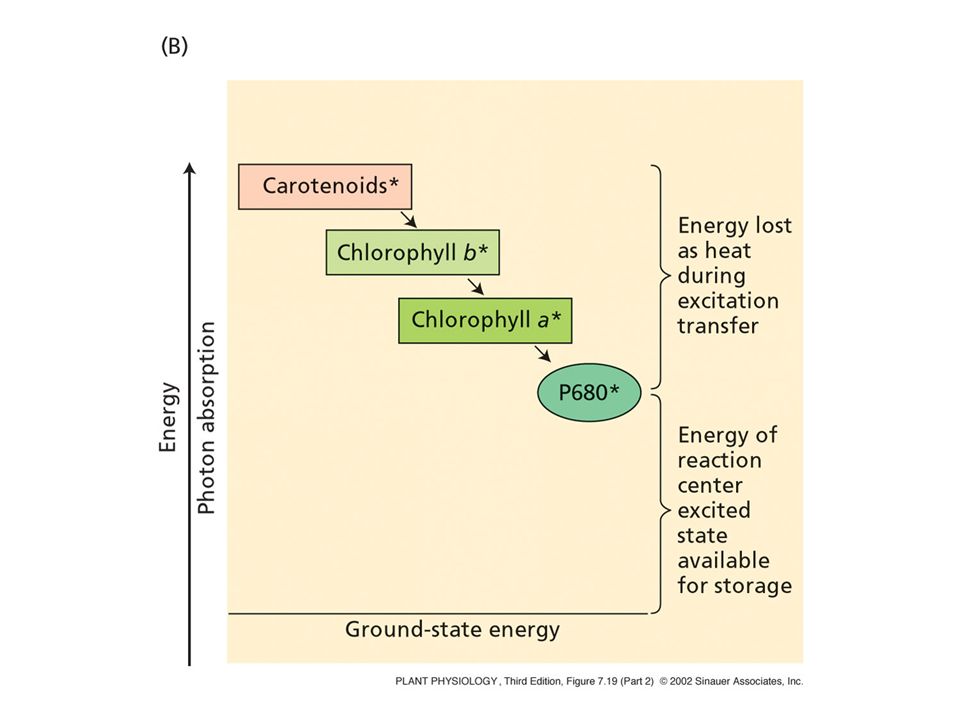
Chl + h = Chl* Eccitazione della clorofilla energia Sb ,* calore
Sa ,* Sb ,* T ,* energia calore Chl + h = Chl* Il livello con la maggiore molteplicità di spin ha energia minore Assorbimento

24
Una molecola eccitata può avere destini diversi
energia Sb ,* calore 1 s trasferimento Sa ,* fluorescenza fotochimica calore T ,* fosforescenza 1 s

25
Assorbimento ed emissione di luce della clorofilla
Sottostati vibrazionali Fluorescenza: la clorofilla eccitata si disfa dell’energia in parte come calore (un aumento del moto disordinato) e in parte come luce a lunghezza d’onda maggiore rispetto a quella di eccitazione
 e in parte come luce a lunghezza d’onda maggiore rispetto a quella di eccitazione.")
26
La fluoroscenza della clorofilla:
Assorbe nel blu e riemette nel rosso It’s like seeing the first time the skyline of New York arriving with a ship. You’re not quite prepared…

27
Percorsi di eccitazione in competizione:
Dispersione termica Fluorescenza Fosforescenza Trasferimento ad un’altra clorofilla Fotochimica k1 Chl* > Chl + calore k2 Chl* > Chl+ h k3 Chl* > Chl + h k4 Chl1*+ Chl > Chl1+ Chl2* k5 Chl* > Chl + e* Resa quantica: resa in prodotti della fotochimica / numero totale di quanti assorbiti Una caratteristica che li distingue è la K di VELOCITA’ con cui avvengono ( s-1) Se la costante di velocità K di una reazione è molto più grande di quella delle reazioni in competizione, lo stato eccitato diverrà de-eccitato prevalentemente attraverso quel processo.
 Se la costante di velocità K di una reazione è molto più grande di quella delle. reazioni in competizione, lo stato eccitato diverrà de-eccitato prevalentemente. attraverso quel processo.")
28
K1 Esempio: fluorescenza S,* > S, + h È una reazione di primo ordine. La scomparsa dello stato eccitato segue la seguente equazione differenziale: -dS/dt = K1S La soluzione è la funz. esponenziale St = S0 e-K1t in cui la fluorescenza decade nel tempo con una funzione esponenziale. S,* tempo k1 =1/ =lifetime =Tempo perchè S passi a 1/e, 37% del livello iniziale. 1/e PROCESSO Dissipazione termica 1 s Fluorescenza s Fosforescenza s Trasferimento Fotochimica Quanto più è piccolo, tanto più veloce il processo

29
energia Sb ,* calore trasferimento Sa ,* fotochimica T ,* fluorescenza fosforescenza

30
TRASFERIMENTO FOTOCHIMICA Fotosistema (centro di reazione) Chl680, Chl700 Chl speciali fanno FOTOCHIMICA

31
Il sistema fotosinthetico è saturato dalla luce
Emerson and Arnold 1932 Evidenza dell’esistenza dell’antenna 2500 clorofille: Ogni centro di reazione ha associato centinaia di pigmenti Deve funzionare più volte prima che venga rilasciata una molecola di O2 1 molecola O2/ 2500 molecole di clorofilla Lampi brevi di luce (10-5 s) e intervalli di buio molto più lungo (0.1 s). Resa quantica = 1 O2/ 9-10 fotoni assorbiti = 0.1 Produzione di O2 Il sistema fotosinthetico è saturato dalla luce Energia del flash di luce (numero di fotoni)
 e intervalli di buio molto più lungo (0.1 s). Resa quantica = 1 O2/ 9-10 fotoni assorbiti. = 0.1. Produzione di O2. Il sistema fotosinthetico è saturato dalla luce. Energia del flash di luce (numero di fotoni)")
32
il centro di reazione riceve gli elettroni attraverso l’antenna esterna (LHC light harvesting complex) e i complessi antenna minori
 e i complessi antenna minori")
33
Le molecole di clorofilla a e b sono disposte in modo diverso nei fotosistemi e nelle antenne
LHC legano Chl a e b PS I core solo Chl a CP43 e CP47 solo Chl a (antenna interna) D1/D2 centro di reazione
 D1/D2 centro di reazione.")
34
Struttura di LHCII, light harvesting complex del PSII
Ogni LHC lega clorofille (13 in LHCII) e carotenoidi LHCII is the main complex and contains about 40% of the PSII chlorophyll – it is the most abundant chlorophyll protein in nature.
 e carotenoidi. LHCII is the main complex and contains about 40% of the PSII chlorophyll – it is the most abundant chlorophyll protein in nature.")
35
Come avviene il trasferimento di energia all’interno dell’antenna?

36
Il trasferimento di energia avviene per risonanza (effetto Förster)
Le molecole devono essere molto vicine l’elettrone eccitato della Chl*1 oscilla e genera un campo elettrico variabile che causa una oscillazione simile nell’elettrone della vicina Chl2 -il processo è simile al trasferimento di E dal fotone alla Chl. Chl1 * + Chl2 Chl1 + Chl2 * -carotene* + Chl -carotene + Chl * Accettore e donatore devono essere separati da piccole distanze La velocità di trasferimento dipende dalla distanza secondo la legge n6 (n= distanza tra le molecole) e dipende anche dall’orientazione Il trasferimento avviene solo verso molecole a energia uguale o più bassa
 e dipende anche dall’orientazione. Il trasferimento avviene solo verso molecole a energia uguale o più bassa.")
38
La Fotochimica avviene nel centro di reazione (in PSII in due clorofille speciali legate alla proteine D1)
")
39
4Fe3+ + 2H2O 4Fe2+ +O2 + 4H+ (isolated chloroplasts)
Robert Hill (1937) Lo ione ferrico funge da accettore degli e- prodotti dall’ossidazione dell’acqua) Potenziale redox (V) 1 -0.5 Chl 680 Chl 680* Buon riducente Ottimo ossidante
 Lo ione ferrico funge da accettore degli e- prodotti dall’ossidazione dell’acqua) Potenziale redox (V) Chl 680. Chl 680* Buon riducente. Ottimo ossidante.")
40
La clorofilla eccitata può tornare allo stato fondamentale rilasciando l’energia come calore o luce o trasferire l’e- ad un accettore. Il risultato è la generazione di 1) donatore(+) e 2) accettore(–) la separazione di carica avviene nei centri di reazione. Energia luminosa energia chimica
 donatore(+) e 2) accettore(–) la separazione di carica avviene nei centri di reazione. Energia luminosa energia chimica.")
41
Nel PSII esiste una coppia speciale di clorofille (P680) dove ha luogo la fotochimica
P680* trasferisce l’e- alla feofitina A separazione di carica. Il P680+ (forma ossidata) viene ridotto a P680 dagli e- che derivano dalla ossidazione dell’acqua
 viene ridotto a P680 dagli e- che derivano dalla ossidazione dell’acqua.")
42
Evidenze dell’esistenza di due fotosistemi: la caduta nel rosso
Resa quantica = (O2 prodotto/ quanti assorbiti) 680
 680.")
43
Evidenze dell’esistenza di due fotosistemi:
Evidenze dell’esistenza di due fotosistemi: Effetto di amplificazione di Emerson >680

44
Effetto antagonistico: if both of far-red light and green light are present, they have opposite effects on cytochrome oxidation

45
Attraverso l’uso di elettrodi e di uno spettrofotometro è possibile misurare lo stato di riduzione di una proteina se cambia la sua assorbanza

46
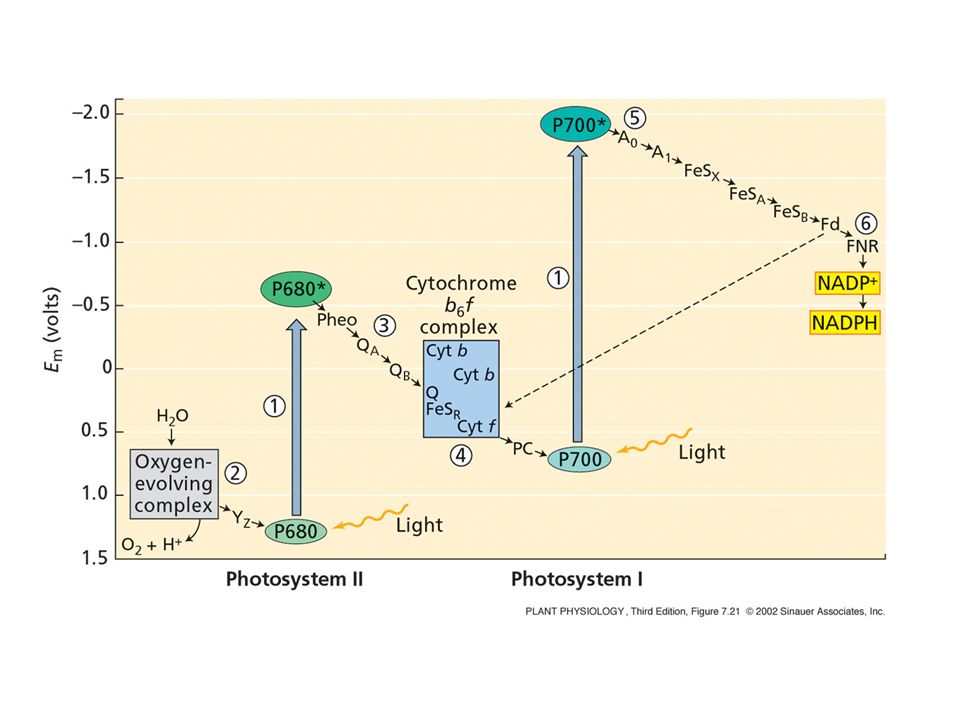
Lo schema a Z 2. Il trasferimento degli elettroni
Gli e- hanno potenziale riducente diverso a seconda del trasportatore a cui sono legati

48
PSII-OEC PSI ATP sintasi Cytb6f
I quattro complessi proteici principali coinvolti nella fotosintesi PSII-OEC PSI ATP sintasi Cytb6f PSII comprende anche l’OEC (Oxygen evolving complex)
")
49
Organization and structure of the four major protein complexes

50
Punto importante: ogni fotosistema contiene molte subunità
Punto importante: ogni fotosistema contiene molte subunità. Di alcune di esse non è chiara la funzione

51
Plastoidrochinone Residuo di Tirosina Donatori e accettori di e- nel processo di separazione di carica in PSII

52
Chinone A riceve un elettrone e lo passa al chinone B che si semi-riduce
Chinone A riceve un secondo elettrone e lo passa a B che si riduce completamente assorbendo 2H+ dallo STROMA. A questo punto può prendere due protoni dal mezzo (stroma) e migrare nella membrana
 e migrare nella membrana.")
53
OEC Oxygen Evolving Complex: - Rompe l’acqua e produce elettroni e H+

54
S1 S2 S0 S3 S4 COMPLESSO CHE EVOLVE L’OSSIGENO
Modello: S0-S4 sono forme sempre più ossidate dell’enzima che contiene Manganese S0 S2 S3 S4 S1 2H2O 4H+ + 02 e-

55
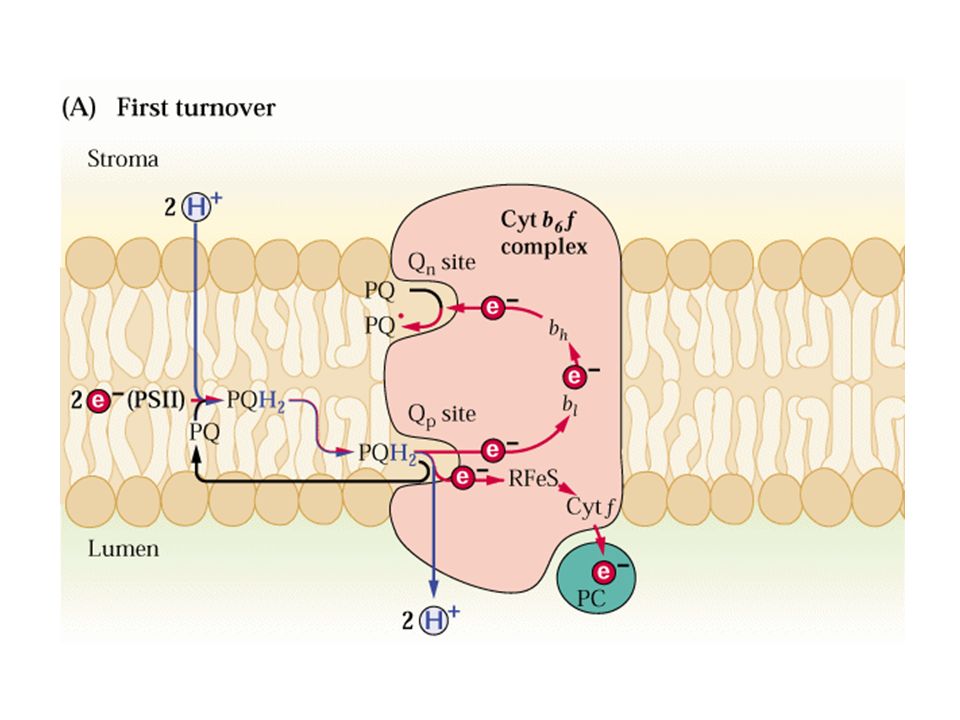
Cytb6f

56
Citocromo b I citocromi dei mitocondri hanno una catena idrofobica GRUPPO EME

57
Citocromo f (e citocromo c)
")
58
Citocromo b6f

59
Il ciclo Q -raddoppia i protoni trasportati dagli elettroni di PQH2
-fra PQH2 e citb6/f c’è un V = -0.4 che viene conservato come pH.

62
PSI

63
Ferredossina Plastocianina Tempi, donatori e accettori di e- nel processo di separazione di carica in PSI

64
The PSI reaction centre complex has a core consisting of 13 different subunits, all the electron transport factors, and about 90 molecules of chlorophyll

65
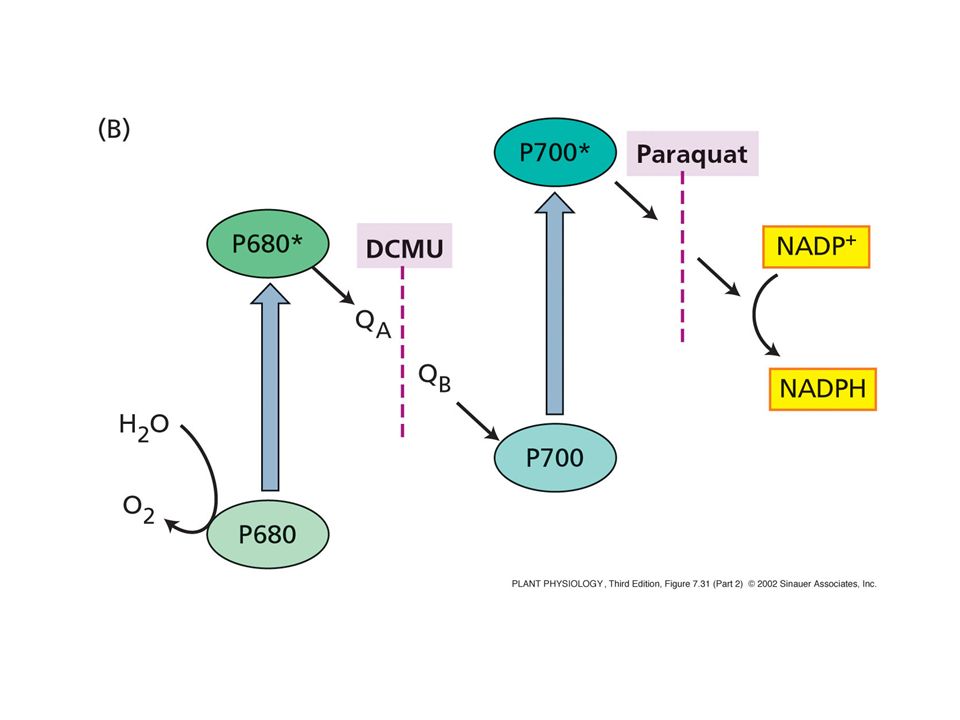
Azione dei diserbanti

67
Accettori di PSI Bild mit PS I Akzeptoren

68
Tutti gli accettori di PSI
1. Il ciclo di Calvin Tutti gli accettori di PSI FNR = fd NADP reduttasi

69
2- Fotofosforilazione ciclica

70
3. La pseudo-ciclica (Mehler)
2 H2O O2 + 4H+ PSII PSI 4 fotoni Fx 2 Ferrodoxin ox 2 Ferredoxin red 2 O2 2 O2 2 H+ O2 Mehler reaction Superossido dismutasi H2O2 Ascorbato perossidasi 2 ascorbato red 2 mono deidro Ascorbato ox

71
4. Assimilazione dell’azoto
A livello delle foglie sia la nitrito reduttasi che la GOGAT consumano equivalenti riducenti. Il flusso di assimilazione primaria è piccolo rispetto a quello della FR GS/GOGAT GS: Glutamin Synthetase (Km = 5 µM) (alles NH4+ wird gebunden) Es gibt auch eine Isoform der GS im Cytosol Glutamat S. 145 und Glutamin aus Lehninger Abb.5.5 zeigt, das weiteres N in Glutamin eingebaut wir und GOGAT Glutamat Synthase (Glutamin-Oxoglutarat-Aminotransferase) Bilder der Enzyme im Lehninger Sinn: das C-Skelett in -Ketogluterarat erhält N Glutamat S. 145 und -Ketoglutarat (Lehninger S. 619) und Glutamin aus Lehninger Bilder der Glutamin-Synthetase 22-5 und Glutamin-Amidotransferase Lehninger 22-8
 (alles NH4+ wird gebunden) Es gibt auch eine Isoform der GS im Cytosol. Glutamat S. 145 und Glutamin aus Lehninger Abb.5.5. zeigt, das weiteres N in Glutamin eingebaut wir. und GOGAT Glutamat Synthase (Glutamin-Oxoglutarat-Aminotransferase) Bilder der Enzyme im Lehninger. Sinn: das C-Skelett in -Ketogluterarat erhält N. Glutamat S. 145 und -Ketoglutarat (Lehninger S. 619) und Glutamin aus Lehninger. Bilder der Glutamin-Synthetase 22-5 und Glutamin-Amidotransferase Lehninger")
72
5. Esporto di equivalenti riducenti al citosol / mitocondrio

73
NADP reductase 4 photons 4 photons 2H+ + 2NADP NADPH 2 H+ Fd PQ CYT B6f O2 + 4H+ PSII PSI 4 e- PC 2H2O The Path of Electron and Proton Flow in Photosynthetic Electron Transport

74
CYCLIC PHOTOPHOSPHORYLATION
Under conditions where NADP+ regeneration is slow, or ATP demand is high, the leaf can cycle electrons between plastoquinone and PSI, and in doing so pump protons across the membrane. This is termed cyclic photophosphorylation. NADP reductase 2 H+ Fd PQ CYT B6f PC PSII PSI 4 H+ CYCLIC PHOTOPHOSPHORYLATION

75
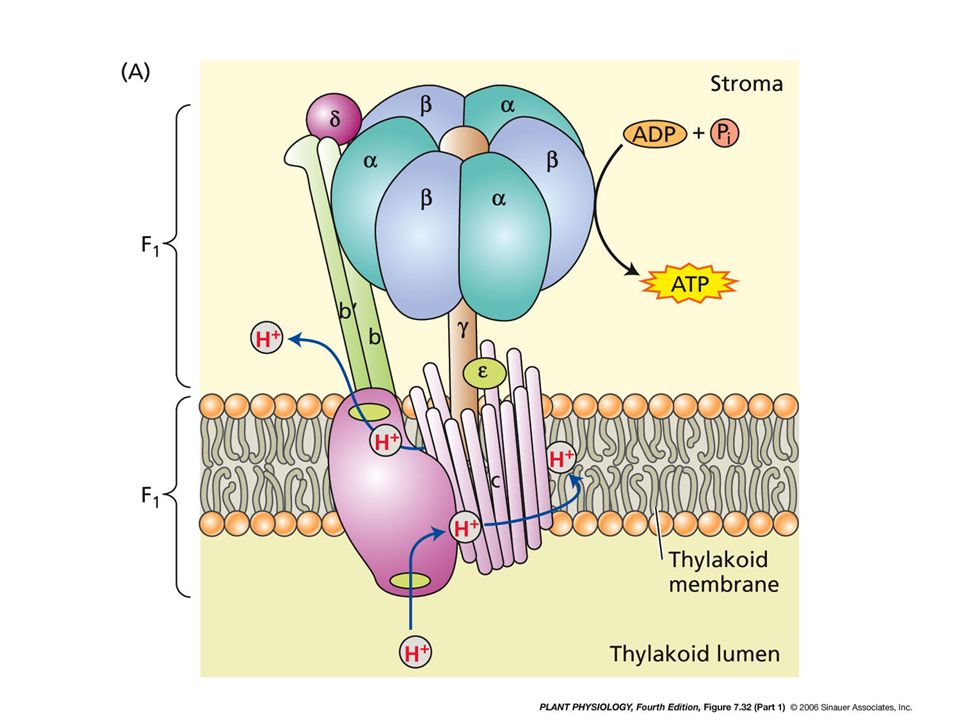
PHOTOPHOSPHORYLATION
Chloroplast stroma – region of high pH ATP Synthase (F-type ATPase) H+ H+ H+ H+ H+ H+ H+ H+ H+ ADP + Pi H+ ATP H+ PHOTOSYSTEM I H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ 3 H+ Thylakoid Lumen – compartment of low pH
 H+ H+ H+ H+ H+ H+ H+ H+ H+ ADP. + Pi. H+ ATP. H+ PHOTOSYSTEM I. H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ 3 H+ Thylakoid Lumen – compartment of low pH.")
76
Tanti accettori per PSI, tra cui alcuni sempre abbondanti cosa significa?
Difficilmente PSI andrà incontro a blocco nel trasporto di e- Riesce sempre a scaricare l’energia assorbita Per PSII la situazione è diversa: può solo (e quindi deve) trasferire gli e- al Cyt b6f (attraverso al plastochinone) e poi a PSI (attraverso la plastocianina) Il PQH2 deve essere ossidato alla stessa velocità con cui viene ridotto. Il pool di plastochinone è piccolo! Quando questo non accade si ha un eccesso di energia su uno dei due fotosistemi. Se è su PSII è pericoloso! Occorre assorbire energia in eccesso dalle clorofille eccitate: esistono diversi meccanismi di fotoprotezione
 trasferire gli e- al. Cyt b6f (attraverso al plastochinone) e poi a PSI (attraverso la plastocianina) Il PQH2 deve essere ossidato alla stessa velocità con cui viene ridotto. Il pool di plastochinone è piccolo! Quando questo non accade si ha un eccesso di energia su uno dei due fotosistemi. Se è su PSII è pericoloso! Occorre assorbire energia in eccesso dalle clorofille eccitate: esistono diversi meccanismi di fotoprotezione.")
77
Se l’eccesso riguarda uno solo dei PS si hanno transizioni di stato
Fotoinibizione light-induced reduction in the photosynthetic capacity Se l’eccesso riguarda uno solo dei PS si hanno transizioni di stato carotenoidi Ciclo delle xantofille Forme reattive dell’ossigeno (radicale Superossido O2-) Riparare le proteine danneggiate
 Riparare le proteine danneggiate.")
79
Zona dove PS II è più abbondante
Zona dove PS I è più abbondante I fotosistemi (PSI e PSII) operano in serie per produrre NADPH, per cui la quantità di eccitazione su essi deve essere paragonabile per massimizzare la performance
 operano in serie per produrre NADPH, per cui la quantità di eccitazione su essi deve essere paragonabile per massimizzare la performance.")
80
Transizioni di stato Quando il PQ è nella forma ridotta, vuol dire che arriva troppa luce su PS II. In questa situazione viene attivata una risposta che fa migrare parte di LHCII La kinasi è attivata da PQH2 membrane tilacoidali impilate LHCII Thr OH libere O-P P ATP ADP Proteina kinasi fosfatasi

81
Long-term changes in illumination lead to changes in photosystem stoichiometry
Short-term changes State transition is a dynamic mechanism that enables plants to respond rapidly to changes in illumination. When PSII is favoured (state 2), the redox conditions in the thylakoids change and result in activation of a protein kinase Takahashi et al., (2006) PNAS 103:477–482 Lunde et al., (2000) Nature 408:
, the redox conditions in the thylakoids change and result in activation of a protein kinase. Takahashi et al., (2006) PNAS 103:477–482. Lunde et al., (2000) Nature 408:")
82
STATO I STATO II

83
Chl + h = Chl* energia Sb ,* calore Sa ,* T ,*
La Chl* trippletto è pericolosa perchè genera facilmente Ossigeno singoletto

84
Carotenoidi duplice funzione:
Condizioni che favoriscono la fotoinibizione: -alta luce -bassa T (rallenta il ciclo di Calvin) Carotenoidi duplice funzione: Carotenoidi fungono da pigmenti accessori -carotene* + Chl -carotene + Chl* Chl + h 1Chl* 3Chl* 3Chl*+ O2 Chl + 1O2 Carotenoidi fungono da dissipatori di energia 1O2 può ossidare numerosi composti: è un composto tossico e occorre eliminarlo Il ciclo delle xantofille ha come scopo proprio quello di dissipare i fotoni In eccesso (quelli che non riescono a essere processati) Plants have mechanisms that protect against adverse effects of strong light. The most studied biochemical protective mechanism is non-photochemical quenching of excitation energy.
 Carotenoidi duplice funzione: Carotenoidi fungono da pigmenti accessori. -carotene* + Chl -carotene + Chl* Chl + h 1Chl* 3Chl* 3Chl*+ O2 Chl + 1O2. Carotenoidi fungono da dissipatori di energia. 1O2 può ossidare numerosi composti: è un composto tossico e occorre eliminarlo. Il ciclo delle xantofille ha come scopo proprio quello di dissipare i fotoni. In eccesso (quelli che non riescono a essere processati) Plants have mechanisms that protect against adverse effects of strong light. The most studied biochemical protective mechanism is non-photochemical quenching of excitation energy.")
85
Schematic representation of singlet oxygen formation upon excitation of chlorophyll (Chl) and the role of carotenoids (Car) in protection against photooxidative damage. The symbol * indicates excited states. Carotenoidi fungono da inattivatori dell’1O2 Carotenoidi dissipano l’eccitazione in eccesso PRIMA che si formi 3Chl*

86
Assorbire energia in eccesso dalle clorofille eccitate
I carotenoidi fungono da dissipatori di energia EPOSSIDAZIONE DE-EPOSSIDAZIONE L’ azione protettiva dei carotenoidi è legata in gran parte al ciclo della violaxantina: in eccesso di luce la violaxantina è convertita in zeaxantina (la reazione è facilitata da un eccesso di H+ nel lume)
")
87
-carotene* + Chl -carotene + Chl * Funzione di antenna
-carotene + Chl* -carotene* + Chl Trasferimento -carotene* -carotene + Calore Dissipazione Cosa decide tra le varie funzioni? la struttura delle molecola (es. violaxantina vs. Zeaxantina) lo stato dei centri di reazione (chiuso/aperto)
 lo stato dei centri di reazione (chiuso/aperto)")
88
Sintesi di ATP La sintesi di ATP funge dissipa il gradiente protonico (un forte gradiente protonico agisce come segnale per aumentare la dissipazione di energia dai complessi antenna) Jagendorf (1967)
 Jagendorf (1967)")
90
Fotosintesi e respirazione

91
Fotosintesi e respirazione
Il lume tilacoidale corrisponde allo spazio intermembrana del mitocondrio

92
Take home messages La fotosintesi comporta l’azione congiunta di 4 complessi Gli elettroni si muovono tra i vari complessi (a motivo dell’energia fornita dalla luce) per mezzo di trasportatori diffusibili (PQ, PC, Fd) Il trasporto di e- è associato ad un trasporto perpendicolare e/o generazione di H+ nel lume ATP e NADPH sono prodotti nella fase luminosa La fotoinibizione è un rischio costante per PSI e numerosi meccanismi cercano di prevenirla
 per mezzo di trasportatori diffusibili (PQ, PC, Fd) Il trasporto di e- è associato ad un trasporto perpendicolare e/o generazione di H+ nel lume. ATP e NADPH sono prodotti nella fase luminosa. La fotoinibizione è un rischio costante per PSI e numerosi meccanismi cercano di prevenirla.")
Presentazioni simili