Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
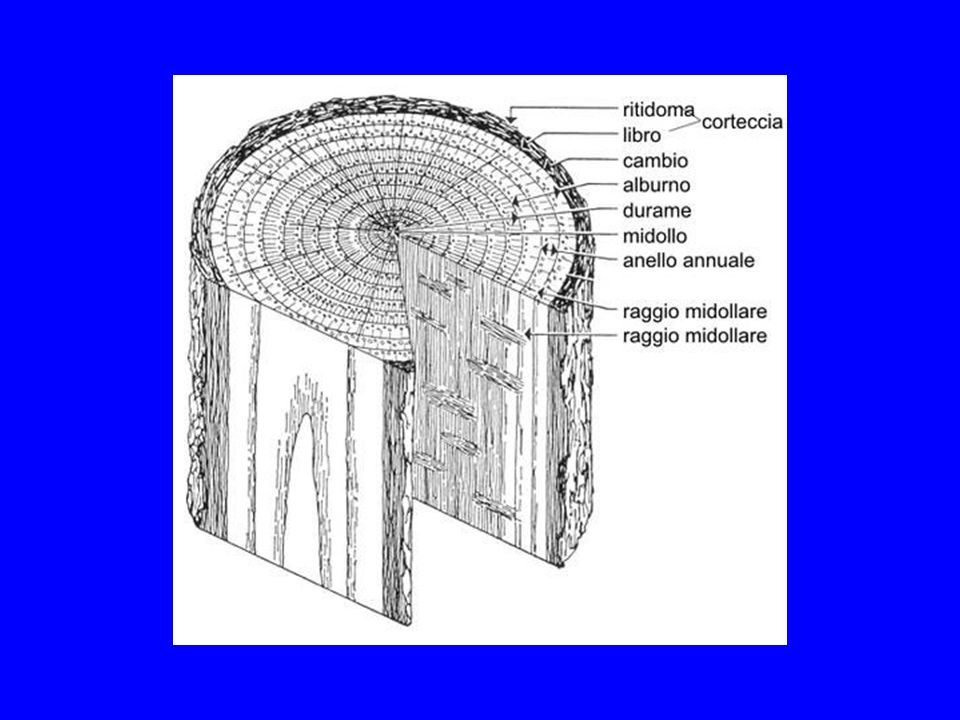
Struttura della Parete vegetale

2
Struttura della Parete vegetale
Lamella mediana: pectina Parete Primaria: emicellulose Parte Secondaria: cellulosa e lignina

3
LE ARMI DEL PATOGENO STRUTTURE MORFOLOGICHE SPECIALIZZATE (Appressori e Austori) Con queste armi il patogeno, in un ospite non resistente o suscettibile, è in grado di aggredire con maggiore o minore successo una pianta. ENZIMI DEGRADATORI TOSSINE

4
Modalità di decomposizione
IFA OH Lip MnPOX e LAC e OH e OH LAC MnPOX Gluc Glc Glc Glc Glc Glc Glc Glc Glc

5
Austori

6
La cellulosa Polimero strutturale della parete vegetale in cui le unità monomeriche di glucosio sono legate con legami b 1-4, l’unità ripetuta è il dimero CELLOBIOSIO La cellulosa è il composto più abbondante sulla terra Gli animali non possiedono enzimi capaci di scindere i legami presenti nella cellulosa

7
La CELLULOSA si presenta con:
Una parte amorfa (più facilmente attaccabile dai microrganismi), costituita da fibrille non legate da legami a idrogeno Una parte cristallina (ad alta organizzazione polimerica e più difficile da aggredire)
, costituita da fibrille non legate da legami a idrogeno. Una parte cristallina (ad alta organizzazione polimerica e più difficile da aggredire)")
8
Degradazione della cellulosa
La cellulosolisi è un processo complesso che richiede la cooperazione di più enzimi Tali enzimi sono secreti all’esterno (enzimi esocellulari) della cellula essendo la cellulosa un polimero insolubile. I microrganismi applicano diverse strategie di degradazione e, di conseguenza, producono tipi di enzimi con diversi meccanismi d’azione. La cellulosa amorfa è degradata più facilmente della cellulosa cristallina
 della cellula essendo la cellulosa un polimero insolubile. I microrganismi applicano diverse strategie di degradazione e, di conseguenza, producono tipi di enzimi con diversi meccanismi d’azione. La cellulosa amorfa è degradata più facilmente della cellulosa cristallina.")
9
Gli enzimi coinvolti nella degradazione della lignina e della cellulosa sono enzimi extracellulari ed in prevalenza enzimi inducibili, cioè espressi solo in presenza del loro substrato. Alcuni di questi enzimi sono espressi costitutivamente (anche in assenza del loro substrato) ma in genere in basse quantità e solo in presenza del substrato la loro produzione aumenta
 ma in genere in basse quantità e solo in presenza del substrato la loro produzione aumenta.")
10
Classificazione degli enzimi cellulosolitici
Gli enzimi cellulosolitici sono rappresentati da un complesso enzimatico a più componenti. La classificazione degli enzimi cellulosolitici non è semplice. Secondo il meccanismo d’azione, gli enzimi cellulosolitici possono essere suddivisi in tre grandi gruppi:

11
Esoglucanasi (cellobioidrolasi): degradano la molecola attaccando le estremità della catena della cellulosa staccando un dimero di cellobiosio Endoglucanasi: degradano attaccando i legami in mezzo alla catena di cellulosa diminuendone in questo modo il grado di polimerizzazione.Sono poco efficaci nella degradazione della cellulosa cristallina (dove sono presenti molti legami H tra le catene). b-glucosidasi degradano i dimeri che si originano dalla cellulosa per azione degli altri enzimi.
. b-glucosidasi degradano i dimeri che si originano dalla cellulosa per azione degli altri enzimi.")
12
Zuccheri semplici inibiscono la sintesi delle cellulasi, la cellulosa comincia ad essere degradata soltanto dopo che sono state consumate altre, più semplici, fonti di carbonio

13
Domini catalitici di diverse glicosil idrolasi
Domini catalitici di diverse glicosil idrolasi. La 5 e la 6 sono due endoglucanasi

14
Azione sinergica di diversi enzimi (endo ed esoglucanasi) nella degradazione della cellulosa
Da Deacon J.W. Micologia moderna Calderini Edagricole, 2000

15
Schema della degradazione della cellulosa ad opera degli enzimi fungini

16
Rappresentazione schematica della demolizione microbica della fibra di cellulosa

17
Strategie di attacco della cellulosa da parte dei microrganismi:
Alcuni funghi possono degradare la cellulosa attraverso l’attività di esoglucanasi, endoglucanasi e b-glucosidasi. Altri degradano producendo solo endoglucanasi. Alcuni batteri anaerobi degradano la cellulosa attraverso un complesso enzimatico multicomposto chiamato cellulosoma.

18
Gli enzimi cellulosolitici, a differenza degli enzimi ligninolitici, esplicano la loro funzione attaccando direttamente il substrato senza utilizzare composti intermedi. Gli enzimi cellulosolitici spesso sono presenti in varie isoforme Isoenzimi- enzimi che catalizzano la stessa reazione, presenti nello stesso organismo in forme molecolari multiple con differenti affinità per il substrato

19
Degradazione Cellulosa
La cellulosa è il polimero naturale più abbondante Catene lineari di glucosio Tre tipi di enzimi fungini che la degradano: le CELLULASI

20
Cellulasi endo--1,4-glucanasi: agisce all’interno della catena polisaccaridica, frammentandola in unità più piccole ed esponendo le terminazioni all’azione dell’enzima CBH cellobioidrolasi (CBH): agisce aggredendo il polimero alle estremità, scindendo successivamente il disaccaride cellobiosio -glucosidasi (o cellobiasi): scinde il cellobiosio in due unità di glucosio.
: agisce aggredendo il polimero alle estremità, scindendo successivamente il disaccaride cellobiosio. -glucosidasi (o cellobiasi): scinde il cellobiosio in due unità di glucosio.")
21
Degradazione Pectina E’ il principale costituente della lamella mediana Catene lineari di un altro zucchero: l’acido galatturonico Gli enzimi degradanti prendono il nome di PECTINASI

22
Pectinasi pectinametilesterasi: deesterifica la pectina a pectato e metanolo pectina liasi: attacca direttamente la pectina, tagliando i legami glicosidici interni pectato liasi: svolge la stessa azione della pectina liasi ma attacca i pectati esopoligalatturonato liasi: attacca le estremità del polimero (polipectato), rilasciando solo digalatturonati poligalatturonasi: la reazione di taglio del poligalatturonato avviene per idrolisi dei legami glicosidici all’interno del polimero e produce oligomeri
, rilasciando solo digalatturonati. poligalatturonasi: la reazione di taglio del poligalatturonato avviene per idrolisi dei legami glicosidici all’interno del polimero e produce oligomeri.")
23
Degradazione Lignina Contiene 3 tipi di composti fenolici uniti tra loro a formare una fitta trama Gli enzimi Ligninolitici sono di due tipi: Rottura anello fenolico Rottura della trama

24
Ligninasi Lignina - perossidasi (conosciuta anche come ligninasi), contenente ferro Fe(III), catalizza la perossidazione delle unità non aromatiche della lignina Manganese - perossidasi, che agisce similmente alla lignino - perossidasi, ma ossida il manganese Mn(II)Mn(III), come elemento mediatore nel trasferimento degli elettroni laccasi (una fenolo - ossidasi), che ossida direttamente la molecola di lignina (in generale ossida i composti fenolici a fenossi - radicali)
, contenente ferro Fe(III), catalizza la perossidazione delle unità non aromatiche della lignina. Manganese - perossidasi, che agisce similmente alla lignino - perossidasi, ma ossida il manganese Mn(II)Mn(III), come elemento mediatore nel trasferimento degli elettroni. laccasi (una fenolo - ossidasi), che ossida direttamente la molecola di lignina (in generale ossida i composti fenolici a fenossi - radicali)")
25
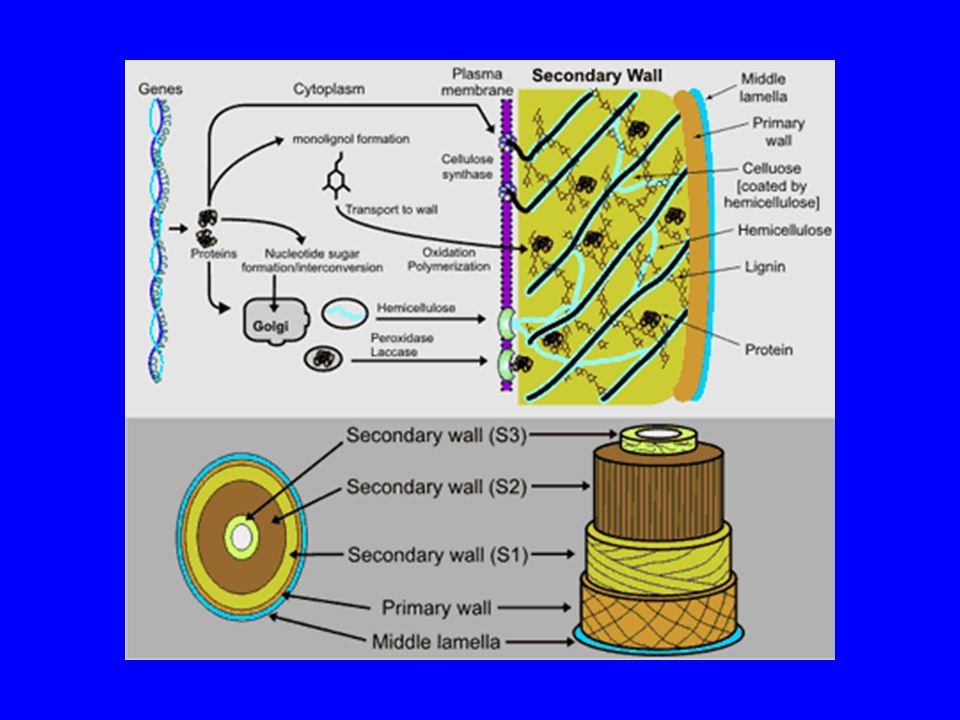
Il legno Parte della pianta che svolge un ruolo di sostegno Composto da diversi tessuti con funzioni distinte Le cellule presentano una parete secondaria ben sviluppata Alcuni strati sono composti da cellule che muoiono a maturità raggiunta (es. xilema)
")
28
Degradazione della lignina
La lignina è un polimero complesso ed insolubile. I “mattoni” costituenti la lignina sono alcool aromatici (alcool cumarilico, coniferilico, sinapilico) La composizione della lignina varia a seconda dei tipi di legno
 La composizione della lignina varia a seconda dei tipi di legno.")
29
Il legno è costituito da cellulosa, emicellulose, lignina e componenti non strutturali
“softwood”: principalmente glucomannani “hardwood”: prevalentemente xilani Componenti non strutturali: estrattivi (composti estraibili con solventi organici) polari (fenoli e tannini) ed apolari (grassi e steroli), composti solubili in acqua (zuccheri ed amidi) La lignina è resistente anche al trattamento con acido solforico concentrato E’ necessaria la cooperazione fra più enzimi per degradare la lignina
 polari (fenoli e tannini) ed apolari (grassi e steroli), composti solubili in acqua (zuccheri ed amidi) La lignina è resistente anche al trattamento con acido solforico concentrato. E’ necessaria la cooperazione fra più enzimi per degradare la lignina.")
30
Modello di struttura della lignina

31
Gli enzimi ligninolitici fungini sono simili per struttura a quelli delle piante coinvolti nella sintesi della lignina: - Appartengono alle stesse classi enzimatiche (per esempio perossidasi) - Le catene peptidiche negli enzimi fungini sono più lunghe, ma presentano molte zone di omologia con quelle delle piante - Il sito attivo contiene un metallo (Fe nelle perossidasi e Cu nella laccasi) - Gli enzimi di diverse specie fungine differiscono tra di loro principalmente per le catene laterali glucidiche
 - Le catene peptidiche negli enzimi. fungini sono più lunghe, ma presentano. molte zone di omologia con quelle delle. piante. - Il sito attivo contiene un metallo (Fe nelle. perossidasi e Cu nella laccasi) - Gli enzimi di diverse specie. fungine differiscono tra di loro principalmente per le. catene laterali glucidiche.")
32
Tutti gli enzimi coinvolti nella degradazione della lignina, fino ad ora studiati, sono extracellulari ed inducibili Il substrato di questi enzimi NON E’ la lignina, ma altre molecole (ad esempio H2O2, veratril alcool) dalle quali si formano radicali liberi, i veri degradatori della lignina. Questo sistema è molto simile a quello utilizzato dalle piante per costruire la lignina
 dalle quali si formano radicali liberi, i veri degradatori della lignina. Questo sistema è molto simile a quello utilizzato dalle piante per costruire la lignina.")
33
Schema dell’azione degli enzimi ligninolitici

34
Gli enzimi ligninolitici sono capaci di degradare altri composti aromatici come fenoli e polifenoli
L’ampia varietà di substrati, è giustificata dal fatto che sono i radicali liberi, creati da questi enzimi, gli agenti degradatori della lignina I più noti enzimi ligninolitici sono: ligninasi (lignina perossidasi), perossidasi manganese dipendente (Mn perossidasi), laccasi,
, perossidasi manganese dipendente (Mn perossidasi), laccasi,")
35
In diversi generi fungini sono presenti diverse combinazioni di enzimi ligninolitici
La laccasi e la Mn perossidasi sono i due tra i più presenti e più studiati Ogni enzima può essere presente in più forme isoenzimatiche L’ossigeno è uno dei regolatori della attività di questi enzimi: concentrazioni troppo basse o troppo alte di O2 sono inibenti

36
Metodi di studio dell’ attività ligninolitica
Lo studio degli enzimi ligninolitici si avvale della capacità di questi enzimi di ossidare composti fenolici diversi dalla lignina Sono stati scelti alcuni composti ossidabili da enzimi ligninolitici in cui la forma ossidata è di colore diverso dalla forma ridotta Uno dei composti utilizzati è la syringaldazina. Normalmente di colore giallino, una volta ossidata diventa viola scuro.

37
Uno degli schemi proposti della ossidazione di un bisfenolo

38
Pathogen ? Host is Rz,Rz or Rz,? ?,? P H AVRz,AVRz or AVRz, ? ?,?
Gene-for-Gene Hypothesis Host is Rz,Rz or Rz,? ?,? AVRz,AVRz or AVRz, ? ?,? H P ? Resistance Disease Pathogen Specific recognition in upper left box only

39
What if the pathogen is a necrotrophic fungus that produces a host selective toxin?
S,S or S,? ?, ? Tox, Tox or Tox, ? ?, ? Disease!! Pathogen P P ? H Resistant Gene-for-gene II Tox = gene for toxin S = gene for receptor Specific recognition in upper left box only ? ? ? H Resistant Resistant

40
Tossine host-selective e fattori di avirulenza
Le tossine host-selective un gruppo di metaboliti strutturalmente complessi e chimicamente diversi prodotte dai fitopatogeni di alcune specie fungine funzionano come determinanti di patogenicità o avirulenza Le risposte biochimiche e molecolari a questi fattori di patogenicità rivela delle risposte tipicamente associate con le difese dell’ospite e l’incompatibilità indotta dai fattori di avirulenza L’azione delle tossine host-selective mostra quindi delle similarità con quelle dei fattori di avirulenza

41
Le HST Le HST sono riportate come “host selective” in quanto sono attive solo contro le piante che sono ospite per quei patogeni che le producono Attualmente la produzione di HST è riportata in almeno 20 patogeni Le HST sono molecole che variano da metaboliti a basso peso molecolare a macromolecole; tutte le HST note sono prodotte da funghi e sono richieste per la patogenicità, infatti la malattia non avviene in assenza della produzione di tossina Nella maggior parte dei casi la sensibilità dell’ospite ad un dato HST è condizionata, generalmente, da un singolo gene e la malattia non avviene in assenza della sensibilità alla tossina La produzione di tossina da parte del patogeno e la sensibilità della pianta alla tossina sono ambedue condizioni causali della malattia Quindi le HST sono dei veri e propri agenti di compatibilità e mediano questo evento inducendo la morte della cellula ospite

42
Fattori di avirulenza In contrasto alle HST i fattori di avirulenza sono degli agenti di incompatibilità Nella maggior parte dei casi il fenotipo predominante dell’incompatibilità è la morte localizzata dell’ospite, comunemente riferita come HR Il ruolo della morte cellulare nella resistenza è in alcuni casi (interazioni con necrotrofi) incerto, ciononostante è una manifestazione tipica della risposta di resistenza La comune associazione della morte cellulare dell’ospite sia con la suscettibilità (HST) che con la resistenza (Avr) fa sorgere la seguente domanda: Qual è la distinzione in queste risposte dell’ospite che tiene conto di tali differenti esiti nelle interazioni ospite-patogeno?
 incerto, ciononostante è una manifestazione tipica della risposta di resistenza. La comune associazione della morte cellulare dell’ospite sia con la suscettibilità (HST) che con la resistenza (Avr) fa sorgere la seguente domanda: Qual è la distinzione in queste risposte dell’ospite che tiene conto di tali differenti esiti nelle interazioni ospite-patogeno")
43
Sintomi Ai più semplici livelli dell’espressione dei sintomi un confronto delle conseguenze di introdurre un HST o un fattore di avirulenza dentro un ospite idoneo nel sito tissutale giusto rivela che entrambi inducono una risposta cultivar-specifica e la morte cellulare rapida seguita dalla morte di zone localizzate o di tutta la pianta L’attività del HST è normalmente saggiata attraverso un’esposizione diretta della pianta o del tessuto alla tossina e simili saggi possono essere utilizzati per determinare l’attività dei fattori di avirulenza Ad. es. i fattori di avirulenza Avr4, Avr9 ed Ecp2 di C. fulvum la proteina INF1 di P. parasitica e la proteina NIP1 di Rhyncosporium secalis sono in grado di elicitare delle risposte di morte cellulare che, macroscopicamente, sono simili a quelle indotte dalla maggior parte degli HST quando introdotti nelle foglie di piante geneticamente appropriate

44
Sintomi Quindi considerando i fattori di avirulenza al di fuori del contesto della resistenza e basandosi sulle risposte fenotipiche, potrebbero essere percepiti come fitotossine selettive Visti da questa prospettiva e considerando le HST come chiari fattori di virulenza, bisogna notare che in origine i fattori di avirulenza NIP erano considerati come tossine, infatti NIP1 stimola l’H+-ATPasi della membrana plasmatica dell’orzo in modo simile alla fusicoccina e probabilmente funziona come fattore di virulenza in assenza del suo corrispondente fattore R (pianta suscettibile) Anche la proteina Ecp2 di C. fulvum è stata inizialmente caratterizzata come fattore di virulenza, infatti i mutanti ecp2 mostravano una scarsa abilità colonizzatrice, una ridotta crescita e conidiazione nell’ospite Quando molti di questi fattori di avirulenza sono introdotti direttamente nel citosol delle piante, evitando quindi lo step di riconoscimento extracellulare, la pianta va in necrosi in modo simile alla reazione causata dalla maggior parte delle HST
 Anche la proteina Ecp2 di C. fulvum è stata inizialmente caratterizzata come fattore di virulenza, infatti i mutanti ecp2 mostravano una scarsa abilità colonizzatrice, una ridotta crescita e conidiazione nell’ospite. Quando molti di questi fattori di avirulenza sono introdotti direttamente nel citosol delle piante, evitando quindi lo step di riconoscimento extracellulare, la pianta va in necrosi in modo simile alla reazione causata dalla maggior parte delle HST.")
45
La vittorina Le similarità nelle risposte indotte dalle HST e dai fattori di avirulenza sono chiaramente illustrati nel Victoria blight (Cochliobolus victoriae) dell’avena a causa della associazione genetica della suscettibilità a questa malattia con la resistenza ad altri patogeni (Puccinia) Il C. victoriae è patogenico grazie alla sua capacità di produrre l’HST vittorina. Tutti gli isolati che producono questa tossina sono patogenici. La sensibilità dell’avena alla vittorina e quindi la suscettibilità al patogeno è condizionata dalla presenza dell’allele dominante Vb Tutti i genotipi che presentano Vb in dominanza sono sia sensibili alla vittorina che suscettibili al patogeno mentre tutti i genotipi omozigoti recessivi sono resistenti a tossina e patogeno Quindi il blight indotto da questo patogeno si ha solo quando un ceppo che produce vittorina attacca una pianta d’avena che porta un allele dominante nel locus Vb
 dell’avena a causa della associazione genetica della suscettibilità a questa malattia con la resistenza ad altri patogeni (Puccinia) Il C. victoriae è patogenico grazie alla sua capacità di produrre l’HST vittorina. Tutti gli isolati che producono questa tossina sono patogenici. La sensibilità dell’avena alla vittorina e quindi la suscettibilità al patogeno è condizionata dalla presenza dell’allele dominante Vb. Tutti i genotipi che presentano Vb in dominanza sono sia sensibili alla vittorina che suscettibili al patogeno mentre tutti i genotipi omozigoti recessivi sono resistenti a tossina e patogeno. Quindi il blight indotto da questo patogeno si ha solo quando un ceppo che produce vittorina attacca una pianta d’avena che porta un allele dominante nel locus Vb.")
46
La vittorina Le piante suscettibili al blight sono resistenti alla Puccinia (crown rust). Questa resistenza è conferita dal gene dominante Pc2 e l’interazione tra avena e crown rust sembra essere la tipica interazione gene-per-gene Il gene Pc2 e il Vb sono probabilmente o “linked” o identici. Questo risultato è derivato da una analisi dei varianti somaclonali generati in presenza della vittorina e indicava, inoltre, che tutti i varianti che diventavano vittorina-insensibili perdevano anche la resistenza al crown rust Quindi la vittorina funzionerebbe come un elicitore per indurre una risposta di resistenza in modo simile a come fa un fattore di avirulenza. Ma perché è percepita come tossina e induce la necrosi per facilitare la patogenesi da parte di un fungo necrotrofico? La vittorina induce una forma di PCD che condivide molte delle caratteristiche morfologiche e biochimiche dell'apoptosi, come il DNA laddering, la condensazione dell'eterocromatina, il disseccamento cellulare e l'attivazione delle proteasi
. Questa resistenza è conferita dal gene dominante Pc2 e l’interazione tra avena e crown rust sembra essere la tipica interazione gene-per-gene. Il gene Pc2 e il Vb sono probabilmente o linked o identici. Questo risultato è derivato da una analisi dei varianti somaclonali generati in presenza della vittorina e indicava, inoltre, che tutti i varianti che diventavano vittorina-insensibili perdevano anche la resistenza al crown rust. Quindi la vittorina funzionerebbe come un elicitore per indurre una risposta di resistenza in modo simile a come fa un fattore di avirulenza. Ma perché è percepita come tossina e induce la necrosi per facilitare la patogenesi da parte di un fungo necrotrofico La vittorina induce una forma di PCD che condivide molte delle caratteristiche morfologiche e biochimiche dell apoptosi, come il DNA laddering, la condensazione dell eterocromatina, il disseccamento cellulare e l attivazione delle proteasi.")
47
La vittorina Risultati preliminari indicano che l'attivazione delle proteasi in cellule vegetali inoculate con vittorina avviene attraverso una cascata che coinvolge la partecipazione di enzimi tipo caspasi La vittorina inoltre contribuisce al malfunzionamento dei mitocondri che è un processo comunemente associato con l'induzione della PCD dato che questa tossina si lega ad alcune proteine del complesso mitocondriale della glicina decarbossilasi (GDC-ciclo dell'azoto) Nelle cellule animali l'alterazione della funzione mitocondriale associata con l'apoptosi è evidenziata da una perdita del potenziale transmembranoso dei mitocondri dovuto ad una transizione di permeabilità (MPT) probabilmente facilitata dall'apertura delle membrane ai soluti con MW<1500 Da Quando i mitocondri dell'avena vanno incontro ad una MPT, la vittorina ha accesso alla matrice mitocondriale dove si lega alla GDC anche se probabilmente questa non è il target primario della tossina per indurre la PCD, che probabilmente si trova a monte della GDC (probabilmente il prodotto del gene Vb)
 Nelle cellule animali l alterazione della funzione mitocondriale associata con l apoptosi è evidenziata da una perdita del potenziale transmembranoso dei mitocondri dovuto ad una transizione di permeabilità (MPT) probabilmente facilitata dall apertura delle membrane ai soluti con MW<1500 Da. Quando i mitocondri dell avena vanno incontro ad una MPT, la vittorina ha accesso alla matrice mitocondriale dove si lega alla GDC anche se probabilmente questa non è il target primario della tossina per indurre la PCD, che probabilmente si trova a monte della GDC (probabilmente il prodotto del gene Vb)")
48
Victorin toxin is bound by 100 kD protein in susceptible plants protein is the P protein component of the glycine decarboxylase complex (GDC) part of an enzyme complex in the mitochondrion involved in the photorespiratory cycle victorin sensitivity gene (LOV1) has been identified in Arabidopsis - typical NBS-LRR gene Are Vb and Pc-2 the same gene? more discussion of this next Tuesday - Sweat et al paper The glycine cleavage system is also known as the glycine decarboxylase complex or GCS. The system is a series of enzymes that are triggered in response to high concentrations of the amino acid glycine. The glycine cleavage system is composed of four proteins: the T-protein, P-protein, L-protein, and H-protein. They do not form
 has been identified in Arabidopsis - typical NBS-LRR gene. Are Vb and Pc-2 the same gene more discussion of this next Tuesday - Sweat et al paper. The glycine cleavage system is also known as the glycine decarboxylase complex or GCS. The system is a series of enzymes that are triggered in response to high concentrations of the amino acid glycine. The glycine cleavage system is composed of four proteins: the T-protein, P-protein, L-protein, and H-protein. They do not form.")
49
Le tossine AAL Le tossine AAL sono un gruppo di HST strutturalmente correlate prodotte da Alternaria alternata con le forme prevalenti TA e TB. Il patogeno causa il cancro dello stelo del pomodoro e la resistenza alla malattia e la sensibilità alle tossine sono condizionate dal locus Asc La resistenza è normalmente dominante, mentre la sensibilità alla tossina è in dominanza incompleta. L'omozigote recessivo è completamente sensibile alla tossina mentre l'eterozigote mostra una sensibilità intermedia. Isolati di Alternaria incapaci di produrre le tossine AAL non causano la malattia e i mutanti compromessi nella sua produzione perdono il fenotipo patogenico Le tossine AAL inducono una risposta di tipo apoptotico in pomodoro sensibile come dimostrato dal DNA laddering e dalla formazione dei corpi di tipo apoptotico. Questa risposta di PCD come nell'HR coinvolge il calcio. L'etilene e quindi mostra similarità ad una risposta di resistenza indotta Le tossine AAL sono delle sfinganine (sphinganine analog mycotoxins – SAM). Altre SAM, note come fumonisine, sono prodotte da Fusarium. Sia le AAL toxins che le fumonisine inducono apoptosi in cellule animali. La fumonisina B1 è selettivamente tossica nei genotipi di pomodoro sensibili come le AAL.
. Altre SAM, note come fumonisine, sono prodotte da Fusarium. Sia le AAL toxins che le fumonisine inducono apoptosi in cellule animali. La fumonisina B1 è selettivamente tossica nei genotipi di pomodoro sensibili come le AAL.")
50
Le tossine AAL Questa similarità di risposta tra le fumonisine e le tossine AAL fa ipotizzare che la PCD indotta da queste coinvolge il signaling delle ceramidi e la distruzione del ciclo cellulare, infatti ambedue le tossine inibiscono la ceramide sintasi con una conseguente alterazione della concentrazione e delle basi sfingoidi nei microcorpi La recente caratterizzazione del locus Asc in pomodoro supporta questa ipotesi, infatti questo gene è omologo al gene di lievito LAG1 (longevity assurance gene) e il suo prodotto compensa la deplezione delle ceramidi facilitando un meccanismo di trasporto sfingolipide-dipendente sia attraverso la produzione di ceramide alternativa oppure codificando una SAM- insensitive sfinganina-N-acetiltrasferasi Le fumonisine - FB1 (e in modo simile le AAL toxins) inducono in At un HR- like response che include la PCD, la formazione di ROS, la deposizione di composti fenolici e del callosio, l'accumulo di fitoalessine e l'espressione delle PRP, inoltre queste risposte richiedono l'attivazione delle pathway del SA, JA ed Et e quindi è marcatamente simile alle risposte di difesa elicitate da un fattore di avirulenza Le piante di At resistenti alla FB1 non risultano, come succedeva in avena con la vittorina, compromesse nella risposta al fattore di avirulenza di P. syringae AvrRpt2, mostrano anzi, una resistenza aumentata nei confronti degli isolati virulenti di questo patogeno
 e il suo prodotto compensa la deplezione delle ceramidi facilitando un meccanismo di trasporto sfingolipide-dipendente sia attraverso la produzione di ceramide alternativa oppure codificando una SAM- insensitive sfinganina-N-acetiltrasferasi. Le fumonisine - FB1 (e in modo simile le AAL toxins) inducono in At un HR- like response che include la PCD, la formazione di ROS, la deposizione di composti fenolici e del callosio, l accumulo di fitoalessine e l espressione delle PRP, inoltre queste risposte richiedono l attivazione delle pathway del SA, JA ed Et e quindi è marcatamente simile alle risposte di difesa elicitate da un fattore di avirulenza. Le piante di At resistenti alla FB1 non risultano, come succedeva in avena con la vittorina, compromesse nella risposta al fattore di avirulenza di P. syringae AvrRpt2, mostrano anzi, una resistenza aumentata nei confronti degli isolati virulenti di questo patogeno.")
51
Genetics of AAL-Toxin Akamatsu et al. 1997 Transformed A. alternata tomato pathotype with plasmid pAN7-1 in presence of restriction enzymes (REMI) Screened for loss of toxin production Obtained toxin-deficient mutants Mutants could not produce any disease on tomato toxin is a pathogenicity factor - compare to virulence factor Conclusion: AAL-toxin is required for pathogenicity on tomato

52
AAL-Toxin and Programmed Cell Death
Wang et al 1996 AAL-toxin inhibits sphinganine N-acyltransferase (ceramide synthase) in plants and animals AAL-toxin-induces programmed cell death (PCD) in plant and animal cells PCD likely involves ceramide (lipid) signalling and disruption of the cell cycle Sphingolipids and sphinganine bases are secondary messengers involved in PCD
 in plants and animals. AAL-toxin-induces programmed cell death (PCD) in plant and animal cells. PCD likely involves ceramide (lipid) signalling and disruption of the cell cycle. Sphingolipids and sphinganine bases are secondary messengers involved in PCD.")
53
De novo sphingolipid synthesis in plants
ceramide synthase De novo sphingolipid synthesis in plants. This scheme has been adapted from a recent report (3) in which enzyme activities and sphingolipid composition in plants were summarized. The conversion of sphinganine to ceramide is inhibited by FB1 and AAL toxins. Variations in plant ceramides exist by the presence of hydroxylated or (poly)unsaturated sphinganine as the sphingoid base moiety or by 2-hydroxylated fatty acyl groups. Predominant complex sphingolipids are glycosylceramides and inositolphosphorylceramides. No plant genes involved in this process have been characterized yet. Brandwagt et al. PNAS 2000;97: ©2000 by The National Academy of Sciences
 in which enzyme activities and sphingolipid composition in plants were summarized. The conversion of sphinganine to ceramide is inhibited by FB1 and AAL toxins. Variations in plant ceramides exist by the presence of hydroxylated or (poly)unsaturated sphinganine as the sphingoid base moiety or by 2-hydroxylated fatty acyl groups. Predominant complex sphingolipids are glycosylceramides and inositolphosphorylceramides. No plant genes involved in this process have been characterized yet. Brandwagt et al. PNAS 2000;97: ©2000 by The National Academy of Sciences.")
54
AAL Toxin Sensitivity to AAL toxin in tomato conferred by a single genetic locus (Gilchrist and Grogan 1976) Named Asc (Alternaria stem canker) locus - codominant Susceptible - asc/asc Resistant - Asc/Asc Intermediate - Asc/asc toxin is effective against plant protoplasts as well as intact leaf tissue ceramide synthases in resistant and susceptible plants are both inhibited equally by AAL toxin - no specificity at this level
 locus - codominant. Susceptible - asc/asc. Resistant - Asc/Asc. Intermediate - Asc/asc. toxin is effective against plant protoplasts as well as intact leaf tissue. ceramide synthases in resistant and susceptible plants are both inhibited equally by AAL toxin - no specificity at this level.")
55
Genetics of Resistance to AAL-Toxin
Brandwagt et al. 2000 cloned and characterized the Asc gene homologous to yeast longevity assurance gene LAG - associated with life span in yeast LAG facilitate ER-to-Golgi transport of glycolsylphosphtidylinositol (GPI)-anchored proteins Asc can partially complement LAG mutations in yeast asc has a premature stop codon relative to Asc and likely codes for a truncated protein (33 kD protein in resistant plants - 10 kD in susceptible plants) does Asc prevent apoptosis by restoring ER-to-Golgi transport?
-anchored proteins. Asc can partially complement LAG mutations in yeast. asc has a premature stop codon relative to Asc and likely codes for a truncated protein (33 kD protein in resistant plants - 10 kD in susceptible plants) does Asc prevent apoptosis by restoring ER-to-Golgi transport")
56
La tossina T La tossina T è l'HST prodotta da Cochliobolus heterostrophus razza T e da Mycospaerella maydis è un polichetide tossico per il mais. Gli isolati che producono tossina sono notevolmente più virulenti dei non produttori. C. heterostrophus causa il southern corn leaf blight una malattia dei genotipi di mais che presentano l'allele cms-T (texas cytoplasm for male sterility), infatti la T-toxin è selettivamente tossica per il mais cms-T I mitocondri sono il target della T-toxin, dove induce il disaccoppiamento della fosforilazione ossidativa, la stimolazione del 4 step della respirazione guidato dal succinato o dal NADH , l'inibizione del 3 step della respirazione guidato dal malato, la perdita di ioni calcio e NAD+ e il rigonfiamento dei mitocondri L'analisi del genoma del mitocondrio ha portato alla scoperta di un unico gene (T-urf3) nei mitocondri di mais cms-T che codifica una proteina di 13KDa localizzata nella membrana mitocondriale. L'espressione di URF13 in coli ha reso le cellule sensibili alla T- toxin. URF13 forma un tetramero nelle membrane del mitocondrio e legandosi direttamente alla T- toxin va incontro ad un cambiamento conformazionale che porta alla formazione di un poro in membrana. Questo induce gli effetti tipici della somministrazione della T-toxin
, infatti la T-toxin è selettivamente tossica per il mais cms-T. I mitocondri sono il target della T-toxin, dove induce il disaccoppiamento della fosforilazione ossidativa, la stimolazione del 4 step della respirazione guidato dal succinato o dal NADH , l inibizione del 3 step della respirazione guidato dal malato, la perdita di ioni calcio e NAD+ e il rigonfiamento dei mitocondri. L analisi del genoma del mitocondrio ha portato alla scoperta di un unico gene (T-urf3) nei mitocondri di mais cms-T che codifica una proteina di 13KDa localizzata nella membrana mitocondriale. L espressione di URF13 in coli ha reso le cellule sensibili alla T- toxin. URF13 forma un tetramero nelle membrane del mitocondrio e legandosi direttamente alla T- toxin va incontro ad un cambiamento conformazionale che porta alla formazione di un poro in membrana. Questo induce gli effetti tipici della somministrazione della T-toxin.")
57
La tossina T Il ruolo della T-toxin nel formare un poro di membrana è particolarmente importante nel contesto della PCD. Infatti i mitocondri sono dei regolatori chiave della PCD sia negli animali che nelle piante. Negli animali la risposta apoptotica è spesso associata con la permeabilizzazione delle membrane mitocondriali attraverso la formazione di pori. Un certo numero di proteine pro-apoptotiche come Bax portano alla formazione di questi pori sia direttamente (come URF13) o indirettamente attraverso l'interazione con un meccanismo interno di formazione dei pori. L'espressione di Bax in tabacco porta a risposte il cui fenotipo ha delle somiglianze con l'HR, tra cui anche l'induzione dell'HR. Dato che la T-toxin induce la formazione di un poro interagendo con URF13 è possibile che questa tossina produca risposte simili all'espressione di Bax Questa ipotesi è stata confermata dall'associazione tra il genotipo cms-T e la PCD
 o indirettamente attraverso l interazione con un meccanismo interno di formazione dei pori. L espressione di Bax in tabacco porta a risposte il cui fenotipo ha delle somiglianze con l HR, tra cui anche l induzione dell HR. Dato che la T-toxin induce la formazione di un poro interagendo con URF13 è possibile che questa tossina produca risposte simili all espressione di Bax. Questa ipotesi è stata confermata dall associazione tra il genotipo cms-T e la PCD.")
58
La tossina HC Il Cochliobolus carbonum causa il northern leaf spot e l'ear rot del mais, la razza 1 di questo patogeno produce l'HC toxin, un tetrapeptide ciclico con un'attività HS contro i genotipi suscettibili del mais Le razze (2 e 3) non produttrici sono comuni e prevalenti in tutto il mondo ma causano lesioni di dimensione inferiore e minor danno su differenti genotipi di mais Quindi la HC-toxin è considerata un fattore di virulenza dato che la razza 1 induce la formazione di lesioni fogliari estese su mais suscettibili ma la tossina non è richiesta per la patogenicità della specie La resistenza al C. carbonum e l'insensibilità alla HC-toxin sono determinate dall'allele dominante nel locus Hm1. Questo gene codifica per una carbonil reduttasi, l'HC-toxin reduttasi, che inattiva la tossina riducendo la funzione chetonica sulla catena laterale dell'acido 2-amino-8- oxo-9,10-epossioctadecanoico che in aggiunta all'epossido è essenziale per la sua attività tossica I genotipi omozigoti per hm1 hm1 (trasnposon disrupted Hm1) mancano dell'attività HCTR e sono completamente suscettibili alla razza1 di C. carbonum
 non produttrici sono comuni e prevalenti in tutto il mondo ma causano lesioni di dimensione inferiore e minor danno su differenti genotipi di mais. Quindi la HC-toxin è considerata un fattore di virulenza dato che la razza 1 induce la formazione di lesioni fogliari estese su mais suscettibili ma la tossina non è richiesta per la patogenicità della specie. La resistenza al C. carbonum e l insensibilità alla HC-toxin sono determinate dall allele dominante nel locus Hm1. Questo gene codifica per una carbonil reduttasi, l HC-toxin reduttasi, che inattiva la tossina riducendo la funzione chetonica sulla catena laterale dell acido 2-amino-8- oxo-9,10-epossioctadecanoico che in aggiunta all epossido è essenziale per la sua attività tossica. I genotipi omozigoti per hm1 hm1 (trasnposon disrupted Hm1) mancano dell attività HCTR e sono completamente suscettibili alla razza1 di C. carbonum.")
59
La tossina HC Alcuni effetti della HC possono essere “benefici” per la pianta, infatti anziché indurre una perdita di elettroliti, incrementa l'uptake dei soluti organici ed inorganici, stimola l'uptake, l'accumulo e la riduzione dei nitrati e provoca una iperpolarizzazione delle membrane Inoltre non ci sono evidenze che l'HC eliciti risposte di difesa (ROS, fitoalessine etc etc) anzi questa tossina sembra agire come repressore di risposte immunitarie. Infatti l'inoculo di foglie di genotipi di mais suscettibili con i conidi di una razza che non produce tossina in combinazione con l'HC risulta nell'espressione di una piena suscettibilità verso un, altrimenti, debolmente virulento patogeno. Questo suggerisce che le risposte dell'ospite che restringono l'abilità della razza 2 del patogeno di causare infezione sono compromesse dall'HC toxin Inoltre inoculando l'ospite con la razza 2 prima di inocularlo con la razza 1 riduce i sintomi della malattia indotti da quest'ultima, rendendoli simili a quelli indotti da quella meno virulenta. Anche in questo caso l'applicazione di HC sopprime la resistenza indotta, confermando il ruolo dell'HC nel prevenire il set-up di una difesa efficace
 anzi questa tossina sembra agire come repressore di risposte immunitarie. Infatti l inoculo di foglie di genotipi di mais suscettibili con i conidi di una razza che non produce tossina in combinazione con l HC risulta nell espressione di una piena suscettibilità verso un, altrimenti, debolmente virulento patogeno. Questo suggerisce che le risposte dell ospite che restringono l abilità della razza 2 del patogeno di causare infezione sono compromesse dall HC toxin. Inoltre inoculando l ospite con la razza 2 prima di inocularlo con la razza 1 riduce i sintomi della malattia indotti da quest ultima, rendendoli simili a quelli indotti da quella meno virulenta. Anche in questo caso l applicazione di HC sopprime la resistenza indotta, confermando il ruolo dell HC nel prevenire il set-up di una difesa efficace.")
60
La tossina HC Tra le risposte genotipo-specifiche elicitate dall'HC è presente un'alterazione della sintesi proteica, che risulta nell'espressione di poche nuove proteine e nella soppressione della sintesi di altre normalmente espresse da genotipi resistenti L'HC non inibisce direttamente la traduzione di nuove proteine indicando che potrebbe avere effetto o sulla trascrizione o sul turnover proteico. Questa tossina, come molti tetrapeptidi ciclici, inibisce l'attività HDAC (istone deacetilasica) che svolge un ruolo fondamentale nel passaggio da etero ad eucromatina L'esame della modalità d'azione dell'HC toxin indica chiaramente che non induce risposte simili a quelle difensive dell'ospite. Quindi non mostra similarità con i fattori di avirulenza, a questo riguardo, però pur avendo una bassa tossicità si comporta in modo evidente come fattore di virulenza L'attività di inibizione della trascrizione di geni di difesa svolta da questa tossina è molto simile a quella svolta da alcuni fattori di avirulenza, come quelli prodotti da Yersinia sp. (T3Es) capaci di interferire con il signaling di pathways strutturali coinvolti con i processi di difesa dell'ospite Quindi la tossina HC potrebbe condividere delle similarità funzionali con altri fattori Avr
 che svolge un ruolo fondamentale nel passaggio da etero ad eucromatina. L esame della modalità d azione dell HC toxin indica chiaramente che non induce risposte simili a quelle difensive dell ospite. Quindi non mostra similarità con i fattori di avirulenza, a questo riguardo, però pur avendo una bassa tossicità si comporta in modo evidente come fattore di virulenza. L attività di inibizione della trascrizione di geni di difesa svolta da questa tossina è molto simile a quella svolta da alcuni fattori di avirulenza, come quelli prodotti da Yersinia sp. (T3Es) capaci di interferire con il signaling di pathways strutturali coinvolti con i processi di difesa dell ospite. Quindi la tossina HC potrebbe condividere delle similarità funzionali con altri fattori Avr.")
61
Struttura tossine

62
Morte cellulare, compatibilità ed incompatibilità
La maggior parte dei patogeni che producono tossine, inclusi quelli che producono HST, sono considerati come necrotrofi. Almeno qualche HST elicita una risposta che richiede la partecipazione dell'ospite nel regolare la morte cellulare. Questo indica che il patogeno per se non uccide l'ospite ma piuttosto è l'ospite che uccide se stesso. Inoltre l'associazione di markers tipici delle risposte di resistenza con quelle della morte cellulare suggerisce che, in alcuni casi, il patogeno potrebbe avvantaggiarsi di queste risposte difensive per facilitare la patogenesi L'espressione di alcuni geni antiapoptotici, come Bcl-2 e Bcl-x1 umani e Ced9 di C. elegans, in pianta, porta alla resistenza verso una serie di necrotrofi come Sclerotinia sclerotiorum, B. cinerea e Cercospora nicotianae. Questa resistenza è legata alla prevenzione della PCD. Infatti l'inoculo di piante WT con S. sclerotiorum produce DNA laddering e PCD mentre in quelle che esprimono Bcl2, la PCD e l'infezione non avvengono. Questo suggerisce che una colonizzazione di successo dell'ospite da parte di questi patogeni dipende da una partecipazione attiva da parte dell'ospite almeno attraverso la regolazione della PCD La dipendenza di questi patogeni dalla PCD dell'ospite per la patogenesi è particolarmente significativa dato che la generazione di ROS è essenziale per la patogenesi

63
Morte cellulare, compatibilità ed incompatibilità
Quindi eventi comunemente associati con la resistenza, come la PCD e la formazione di ROS, sono coinvolti con la suscettibilità verso i necrotrofi La formazione di ROS e il concomitante redox signaling giocano un ruolo centrale nell'integrazione di diversi tipi di risposte difensive della pianta e la produzione di ROS a prescindere dalla sorgente può essere sufficiente per elicitare la risposta di difesa. Inoltre la generazione di ROS è un marker che indica che le risposte di difesa sono attivate La cercosporina, la tossina prodotta da molte specie di Cercospora, viene attivata dalla luce in presenza di ossigeno ed elicita la produzione di ossigeno singoletto e aso. Anche S. sclerotiorum e B. cinerea inducono la produzione di ROS da parte dell'ospite. La quantità di ROS prodotta è positivamente correlata con l'abilità di un dato isolato di provocare la malattia La morte cellulare dell'ospite è comunemente associata ed è un elemento importante nella resistenza per i biotrofi obbligati. Un ruolo generale per questo tipo di risposta è da escludere in quanto la PCD non contribuisce in modo sostanziale alla resistenza verso i necrotrofi e emibiotrofi

Presentazioni simili

>")

















