Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Sequenze IRES virali Internal ribosome entry sites (IRESs) are RNA elements that mediate end-independent ribosomal recruitment to internal locations in mRNA. Structurally related viral IRESs use distinct mechanisms, based on non-canonical interactions with eukaryotic initiation factors (eIFs) and/or 40S subunits (see the figure). Initiation on type 1 and type 2 IRESs involves their specific binding to the p50 domain of eIF4G (FIG.3a), which is enhanced by eIF4A112–114, on type 3 IRESs involves their interaction with the eIF3 and 40S subunit components of 43S complexes8,115 and on type 4 IRESs involves their binding to 40S subunits7,116. The eIF4G–eIF4A complex recruits 43S complexes to type 1 and type 2 IRESs without the involvement of eIF4E. Type 3 IRESs directly attach 43S complexes to the initiation codon independently of eIF4F, eIF4B, eIF1 and eIF1A, whereas type 4 IRESs initiate without eIFs or tRNAi Met (the P-site of the 40S subunit is occupied by an IRES domain that mimics codon–anticodon base pairing). Hence, IRES-mediated initiation might be resistant to cellular regulatory mechanisms, such as eIF2 phosphorylation (type 4 IRESs) and/or eIF4E sequestration (all types of IRESs)116. Initiation on some IRESs also requires IRES trans-acting factors (ITAFs) — RNA-binding proteins that are thought to stabilize the optimal three-dimensional IRES conformation117. The list of cellular mRNAs that are thought to contain IRESs is growing and, although a recent stringent test has questioned some of these claims118, it would be prudent to presume that many are still valid. Cellular IRESs show little structural relationship to each other and their underlying mechanism remains largely unknown but probably follows the picornavirus paradigm of binding the eIF4G–eIF4A complex. Importantly, cellular IRES-containing mRNAs can also be translated by the scanning mechanism, which raises the crucial question of what regulates the switch between these modes of initiation. One key parameter might be the intracellular concentration of eIF4G. The concentration of eIF4G (but not eIF4E) is highly elevated in many advanced breast cancers, and in inflammatory breast cancer this results in efficient IRES-dependent translation of p120 catenin and vascular endothelial growth factor (VEGF) mRNAs119. In other breast cancer cell lines with high eIF4G levels, overexpression of eIF4E-binding protein 1 (4E-BP1), to sequester eIF4E, coupled with hypoxia, activates VEGF and hypoxia-inducible factor 1α (HIF1A) IRESs120. Another parameter that may determine which mechanism predominates is the intracellular concentration of ITAFs. REVIEWS 118 | FEBRUARy 2010 | vOlUME 10
 and/or 40S subunits (see the figure). Initiation on type 1 and type 2 IRESs involves their. specific binding to the p50 domain of eIF4G (FIG.3a), which is enhanced by eIF4A112–114, on type 3 IRESs involves their. interaction with the eIF3 and 40S subunit components of 43S complexes8,115 and on type 4 IRESs involves their binding to. 40S subunits7,116. The eIF4G–eIF4A complex recruits 43S complexes to type 1 and type 2 IRESs without the involvement. of eIF4E. Type 3 IRESs directly attach 43S complexes to the initiation codon independently of eIF4F, eIF4B, eIF1 and. eIF1A, whereas type 4 IRESs initiate without eIFs or tRNAi. Met (the P-site of the 40S subunit is occupied by an IRES domain. that mimics codon–anticodon base pairing). Hence, IRES-mediated initiation might be resistant to cellular regulatory. mechanisms, such as eIF2 phosphorylation (type 4 IRESs) and/or eIF4E sequestration (all types of IRESs)116. Initiation on. some IRESs also requires IRES trans-acting factors (ITAFs) — RNA-binding proteins that are thought to stabilize the. optimal three-dimensional IRES conformation117. The list of cellular mRNAs that are thought to contain IRESs is growing and, although a recent stringent test has. questioned some of these claims118, it would be prudent to presume that many are still valid. Cellular IRESs show little. structural relationship to each other and their underlying mechanism remains largely unknown but probably follows the. picornavirus paradigm of binding the eIF4G–eIF4A complex. Importantly, cellular IRES-containing mRNAs can also be. translated by the scanning mechanism, which raises the crucial question of what regulates the switch between these. modes of initiation. One key parameter might be the intracellular concentration of eIF4G. The concentration of eIF4G. (but not eIF4E) is highly elevated in many advanced breast cancers, and in inflammatory breast cancer this results in. efficient IRES-dependent translation of p120 catenin and vascular endothelial growth factor (VEGF) mRNAs119. In other. breast cancer cell lines with high eIF4G levels, overexpression of eIF4E-binding protein 1 (4E-BP1), to sequester eIF4E, coupled with hypoxia, activates VEGF and hypoxia-inducible factor 1α (HIF1A) IRESs120. Another parameter that may. determine which mechanism predominates is the intracellular concentration of ITAFs. REVIEWS. 118 | FEBRUARy 2010 | vOlUME 10")
2
IRES negli mRNA cellulari

3
Strutture secondarie delle IRES
Figure 2 Secondary structural models of c-myc (a) and L-myc IRESs (b). c-myc or L-myc RNA was treated with chemicals (CMCT, kethoxal, DMS) to detect single-stranded regions or with RNase VI to detect double-stranded regions. This information was then used to add constraints to the m-fold algorithm to obtain the structural model shown. The c-myc IRES has a more open and flexible structure than the L-myc IRES. However, interestingly both IRES contain potential pseudoknots close to the region where the ribosome lands
 and L-myc IRESs (b). c-myc or L-myc RNA was treated with chemicals (CMCT, kethoxal, DMS) to detect single-stranded regions or with RNase VI to detect double-stranded regions. This information was then used to add constraints to the m-fold algorithm to obtain the structural model shown. The c-myc IRES has a more open and flexible structure than the L-myc IRES. However, interestingly both IRES contain potential pseudoknots close to the region where the ribosome lands.")
4
Fattori trans-agenti delle IRES

5
Ruolo delle IRES

6
Meccanismi di traduzione negli eucarioti
Figure 1: Schematic representation of eukaryotic mRNAs. (a) Features of a conventional mRNA. The red circle at the 5_end depicts the cap (m7Gppp); A(n) depicts the poly(A) tail at the 3_end. (b) (c) Schematic of atypical RNA structures, stem loops (black hairpin), or uAUGs, respectively, located in mRNAs translated via cap-dependent initiation. (d) RNA structural elements located in the 3_untranslated region of viral RNAs mediating cap-independent translation (3_CITE). Different types of IRES elements found in the viral RNA of picornaviruses (e), hepatitis C (f), and dicistroviruses (g) are schematically depicted in red.
 Features of a conventional mRNA. The red circle at the 5_end depicts the cap (m7Gppp); A(n) depicts the poly(A) tail at the 3_end. (b) (c) Schematic of atypical RNA structures, stem loops (black hairpin), or uAUGs, respectively, located in mRNAs translated via cap-dependent initiation. (d) RNA structural elements located in the 3_untranslated region of viral RNAs mediating cap-independent translation (3_CITE). Different types of IRES elements found in the viral RNA of picornaviruses (e), hepatitis C (f), and dicistroviruses (g) are schematically depicted in red.")
7
Regolazione della traduzione
generale specifica

8
Initiation Factors prokaryotes eukaryotes
Activity prokaryotes eukaryotes IF3 eIF-1 Fidelity of AUG codon recognition IF1 eIF-1A Facilitate Met-tRNAiMet binding to small subunit eIF-2 Ternary complex formation eIF-2B (GEF) GTP/GDP exchange during eIF-2 recycling eIF-3 (12 subunits) Ribosome antiassociation, binding to 40S eIF-4F (4E, 4A, 4G) mRNA binding to 40S, RNA helicase activity eIF-4A ATPase-dependent RNA helicase eIF-4E 5' cap recognition eIF-4G Scaffold for of eIF-4E and -4A eIF-4B Stimulates helicase, binds with eIF-4F eIF-4H Similar to eIF4B eIF-5 Release of eIF-2 and eIF-3, GTPase IF2 eIF5B Subunit joining eIF-6 Ribosome subunit antiassociation
 GTP/GDP exchange during eIF-2 recycling. eIF-3 (12 subunits) Ribosome antiassociation, binding to 40S. eIF-4F (4E, 4A, 4G) mRNA binding to 40S, RNA helicase activity. eIF-4A. ATPase-dependent RNA helicase. eIF-4E. 5 cap recognition. eIF-4G. Scaffold for of eIF-4E and -4A. eIF-4B. Stimulates helicase, binds with eIF-4F. eIF-4H. Similar to eIF4B. eIF-5. Release of eIF-2 and eIF-3, GTPase. IF2. eIF5B. Subunit joining. eIF-6. Ribosome subunit antiassociation.")
10
Via delle MAP chinasi

11
stress Fig. 2. Signal transduction pathways that regulate the function of general translation factors. Extracellular signals (growth factors, hormones, mitogens and cytokines) stimulate translation initiation via glycogen-synthase kinase 3 (GSK3) and phosphoinositol 3-kinase (PI3K). PI3K activates Akt through phosphorylation of phosphatidylinositol. This leads to activation of an as yet unidentified kinase(s) X and probably also of mTOR (mammalian target of rapamycin) kinase. Kinase(s) X and mTOR facilitate the release of the eukaryotic translation initiation factor 4E (eIF4E) by phosphorylating the 4E-binding proteins (4E-BPs). Released eIF4E can then join the complex eIF4F, leading to cap binding and ribosomal scanning. When sufficient amino acids and ATP are available, mTOR signals to p70S6k and possibly to kinase(s) X. p70S6k phosphorylates the S6 ribosomal subunit that is necessary to release inhibition of 5′ terminal oligopyrimidine tract (5′ TOP) mRNAs. These mainly encode components of the translational machinery. The tumour suppressor, PTEN, inhibits PI3K signalling through dephosphorylation of phosphatidylinositol. The drug, rapamycin, and its pharmacologically used derivative, CCI-779, inhibit mTOR. Active GSK3 phosphorylates eIF2B, thereby inhibiting the recycling of eIF2. When GSK3 is inhibited by extracellular signals, eIF2B is active and translation initiation is stimulated by the recycling of eIF2. The eIF2 cycle is also inhibited by phosphorylation of the eIF2α subunit. Conditions of stress lead to suppression of translation initiation by activation of the eIF2α kinases, haem-regulated inhibitor kinase (HRI), RNA-dependent protein kinase (PKR), PKR-like endoplasmic-reticulum kinase (PERK), and GCN2. Distinct translation initiation factors regulate the translation of specific mRNA pools, either alone or in combination with other factors. Abbreviations: dsRNA, double-stranded RNA; ER, endoplasmic reticulum.
 stimulate translation initiation via glycogen-synthase kinase 3 (GSK3) and phosphoinositol 3-kinase (PI3K). PI3K activates Akt through phosphorylation of phosphatidylinositol. This leads to activation of an as yet unidentified kinase(s) X and probably also of mTOR (mammalian target of rapamycin) kinase. Kinase(s) X and mTOR facilitate the release of the eukaryotic translation initiation factor 4E (eIF4E) by phosphorylating the 4E-binding proteins (4E-BPs). Released eIF4E can then join the complex eIF4F, leading to cap binding and ribosomal scanning. When sufficient amino acids and ATP are available, mTOR signals to p70S6k and possibly to kinase(s) X. p70S6k phosphorylates the S6 ribosomal subunit that is necessary to release inhibition of 5′ terminal oligopyrimidine tract (5′ TOP) mRNAs. These mainly encode components of the translational machinery. The tumour suppressor, PTEN, inhibits PI3K signalling through dephosphorylation of phosphatidylinositol. The drug, rapamycin, and its pharmacologically used derivative, CCI-779, inhibit mTOR. Active GSK3 phosphorylates eIF2B, thereby inhibiting the recycling of eIF2. When GSK3 is inhibited by extracellular signals, eIF2B is active and translation initiation is stimulated by the recycling of eIF2. The eIF2 cycle is also inhibited by phosphorylation of the eIF2α subunit. Conditions of stress lead to suppression of translation initiation by activation of the eIF2α kinases, haem-regulated inhibitor kinase (HRI), RNA-dependent protein kinase (PKR), PKR-like endoplasmic-reticulum kinase (PERK), and GCN2. Distinct translation initiation factors regulate the translation of specific mRNA pools, either alone or in combination with other factors. Abbreviations: dsRNA, double-stranded RNA; ER, endoplasmic reticulum.")
12
Chinasi di eIF2a Stress-Responsive eIF2 Kinases Inhibit General Translation yet Stimulate Expression of a Special Class of Genes (A) Architecture of the four eIF2 Ser-51 kinases. The conserved kinase domains (KD) are depicted in green. The two heme-binding sites in HRI are marked in red. The dsRNA binding domains (dsRBD) in PKR are shown in blue. The N-terminal half of PERK resembles the corresponding domain of the ER stress-responsive IRE1 kinase. Also indicated are signal peptide (SP) and transmembrane (TM) domain of PERK. The regulatory histidyl-tRNA synthetase (HisRS) domain in GCN2 is shown in purple. The locations of the N-terminal GCN1 binding domain (blue), charged region (+/-), and pseudokinase domain (KD), as well as the C-terminal ribosome binding and dimerization domain (RB/DD) in GCN2 are also indicated. The activating condition or ligand for each kinase and the known sources of the kinases are also indicated.
 Architecture of the four eIF2 Ser-51 kinases. The conserved kinase domains (KD) are depicted in green. The two heme-binding sites in HRI are marked in red. The dsRNA binding domains (dsRBD) in PKR are shown in blue. The N-terminal half of PERK resembles the corresponding domain of the ER stress-responsive IRE1 kinase. Also indicated are signal peptide (SP) and transmembrane (TM) domain of PERK. The regulatory histidyl-tRNA synthetase (HisRS) domain in GCN2 is shown in purple. The locations of the N-terminal GCN1 binding domain (blue), charged region (+/-), and pseudokinase domain (KD), as well as the C-terminal ribosome binding and dimerization domain (RB/DD) in GCN2 are also indicated. The activating condition or ligand for each kinase and the known sources of the kinases are also indicated.")
13
Azione delle chinasi di eIF2a

14
Risposta UPR (Unfolded Protein Response)
A simplified diagram of the initiation of the UPR by prolonged and overwhelming protein malfolding. Grp78 recruitment to chaperone the malfolded proteins results in Grp78 dissociation from its conformational binding state of the transmembrane receptor proteins PERK, IRE1 and ATF6. Dissociation results in receptor homodimerisation and oligomerisation to an active state. The activated cytosolic domain of PERK phosphorylates the eIF2alpha, inhibiting translation and resulting in cell cycle arrest. The activated cytosolic domain of IRE1 cleaves the 252bp intron from its substrate XBP1, facilitating its translation to form the transcription factor XBP1. Activated ATF6 translocates to the Golgi, cleaved by proteases to form an active 50kDa fragment (ATF6 p50). ATF6 p50 and XBP1 bind ERSE promoters in the nucleus to produce upregulation of the proteins involved in the unfolded protein response.
. ATF6 p50 and XBP1 bind ERSE promoters in the nucleus to produce upregulation of the proteins involved in the unfolded protein response.")
15
Risposta UPR (Unfolded Protein Response)
Signal transduction events associated with ER stress and UPR. Upon accumulation of unfolded or misfolded proteins in the ER three major ER stress sensors, PERK, ATF6 and IRE1, are activated following their dissociation from the ER chaperone GRP78. Activated PERK phosphorylates eukaryotic initiation factor 2α (eIF2α), which suppresses global mRNA translation but activates ATF4 translation. ATF4 translocates to the nucleus and induces the transcription of genes required to restore ER homeostasis. Activation of PERK also leads to the induction of CHOP (C/EBP homologous protein), which is involved in pro-apoptotic signaling. ATF6 is activated by proteolysis mediated by proteases S1P and S2P after its translocation from the ER to the Golgi apparatus. Active ATF6 translocates to the nucleus and regulates the expression of ER chaperones and X box-binding protein 1 (XBP1) to facilitate protein folding, secretion, and degradation in the ER. Xbp1 mRNA undergoes unconventional mRNA splicing carried out by IRE1. Spliced XBP1 protein (sXBP1) translocates to the nucleus and controls the transcription of chaperones, the co-chaperones and the PERK-inhibitor P58IPK, as well as genes involved in protein degradation.
, which suppresses global mRNA translation but activates ATF4 translation. ATF4 translocates to the nucleus and induces the transcription of genes required to restore ER homeostasis. Activation of PERK also leads to the induction of CHOP (C/EBP homologous protein), which is involved in pro-apoptotic signaling. ATF6 is activated by proteolysis mediated by proteases S1P and S2P after its translocation from the ER to the Golgi apparatus. Active ATF6 translocates to the nucleus and regulates the expression of ER chaperones and X box-binding protein 1 (XBP1) to facilitate protein folding, secretion, and degradation in the ER. Xbp1 mRNA undergoes unconventional mRNA splicing carried out by IRE1. Spliced XBP1 protein (sXBP1) translocates to the nucleus and controls the transcription of chaperones, the co-chaperones and the PERK-inhibitor P58IPK, as well as genes involved in protein degradation.")
16
Interferenza dei virus nella traduzione negli eucarioti
Fig. 1. Model of key features of cap eukaryotic initiation factor (eIF) 4F-dependent translation initiation in mammalian cells and the sites of interference by animal viruses (indicated in dark blue text). eIF4E cap-binding protein binds the m7G-capped end of the mRNA, together with eIF4G, eIF4A (which together form the cap-binding complex eIF4F), eIF3, poly(A)-binding protein (PABP), and the eIF4E kinase Mnk1. The assembly of all factors is collectively referred to here as the cap-initiation complex, which also includes the 40S ribosome subunit with associated ternary complex consisting of eIF2–GTP–met-tRNAi. The order of assembly of the cap-initiation complex is not well known. It is also not well understood whether the eIF4F complex remains associated with the capped end of the mRNA as drawn here, tracks with the scanning 40S ribosome subunit or dissociates from the mRNA. Interference with the dsRNA-activated protein kinase (PKR) activation by a variety of viruses is shown with identified viral gene products when known. Arrows indicate activating responses, whereas bars indicate inhibiting responses. The eIF4E-sequestering protein, 4E-binding protein 1 (4E-BP1), is inactivated by phosphorylation by the kinase mTOR, which is activated by the phosphatidyl inositol 3 kinase (PI3-K)/Akt pathway. Highlighted in this figure are the multiple mechanisms for inhibition of PKR activation by viral dsRNA: sequestration of dsRNA by viral proteins [herpes simplex virus (HSV) Us11, vaccinia virus (VV) E3L and reovirus s3 (Reo s3)], inhibition of PKR itself by inhibitory viral dsRNA mimetics [adenovirus virus associated (VA) RNAI, Epstein–Barr virus (EBV)- encoded small nuclear RNAs (EBER) and possibly HIV TAR], facilitation of eIF2a recycling (HSV g34.5 protein) and a more poorly characterized mechanism [human cytomegalovirus (HCMV), human herpesvirus 8 (HHV8) interferon regulatory factor (vIRF2) protein]. The components of these pathways are not represented to scale.
 4F-dependent translation initiation in mammalian cells and the sites of interference by animal viruses (indicated in dark blue text). eIF4E cap-binding protein binds the m7G-capped end of the mRNA, together with eIF4G, eIF4A (which together form the cap-binding complex eIF4F), eIF3, poly(A)-binding protein (PABP), and the eIF4E kinase Mnk1. The assembly of all factors is collectively referred to here as the cap-initiation complex, which also includes the 40S ribosome subunit with associated ternary complex consisting of eIF2–GTP–met-tRNAi. The order of assembly of the cap-initiation complex is not well known. It is also not well understood whether the eIF4F complex remains associated with the capped end of the mRNA as drawn here, tracks with the scanning 40S ribosome subunit or dissociates from the mRNA. Interference with the dsRNA-activated protein kinase (PKR) activation by a variety of viruses is shown with identified viral gene products when known. Arrows indicate activating responses, whereas bars indicate inhibiting responses. The eIF4E-sequestering protein, 4E-binding protein 1 (4E-BP1), is inactivated by phosphorylation by the kinase mTOR, which is activated by the phosphatidyl inositol 3 kinase (PI3-K)/Akt pathway. Highlighted in this figure are the multiple mechanisms for inhibition of PKR activation by viral dsRNA: sequestration of dsRNA by viral proteins [herpes simplex virus (HSV) Us11, vaccinia virus (VV) E3L and reovirus s3 (Reo s3)], inhibition of PKR itself by inhibitory viral dsRNA mimetics [adenovirus virus associated (VA) RNAI, Epstein–Barr virus (EBV)- encoded small nuclear RNAs (EBER) and possibly HIV TAR], facilitation of eIF2a recycling (HSV g34.5 protein) and a more poorly characterized mechanism [human cytomegalovirus (HCMV), human herpesvirus 8 (HHV8) interferon regulatory factor (vIRF2) protein]. The components of these pathways are not represented to scale.")
17
Meccanismi virali d'inibizione della fosforilazione di eIF2a

18
Regolazione di eIF2a Fosforilazione su ser 51 in risposta a stress da parte di 4 chinasi: PKR, GCN2, PERK, HRI La fosforilazione impedisce il riciclo da parte di eIF2B La traduzione di alcuni mRNA è stimolata da bassi livelli di eIF2a attivo Molti virus hanno sistemi per impedire la fosforilazione di eIF2a

19
eIF4F

20
Fosforilazione di 4E-BP
Fig. 1. mTOR regulates the phosphorylation and function of several proteins involved in mRNA translation. Phosphorylation of S6Ks leads to their activation. S6Ks phosphorylate rpS6 and is thought to play a role in the regulation of the translation of the subset of mRNAs containing a 50- terminal oligopyrimidine tract (TOP) mRNAs. S6Ks also phosphorylate elongation factor 2 (eEF2) kinase leading to a decrease in its activity at basal calcium concentrations although additional inputs are also involved in its control, such an independent input to the phosphorylation of Ser78, which inhibits the binding of CaM to eEF2 kinase (see text). Inactivation of eEF2 kinase leads to the dephosphorylation of eEF2 and the activation of elongation. Phosphorylation of 4E-BP1 at certain sites leads to its release from eIF4E, which can then interact with eIF4G to form initiation complexes than recruit the 40S ribosomal subunit to the 50-end of the mRNA. Amino acids act as positive modulators of mTOR. The mechanisms by which amino acids exert this effect are unclear, but recent evidence suggests this involves sensing of intracellular amino acid levels. Insulin and a range of other stimuli increase the phosphorylation of S6Ks and 4E-BP1. Recent work indicates that this effect is likely mediated through the tuberous sclerosis complex (TSC1/2) [46]. The question mark by the role of S6 phosphorylation in the translation of 50-TOP mRNAs denotes the fact that Tang et al. [47] have recently challenged the prevailing concept that these mRNAs are regulated via S6 kinases/phosphorylation of rpS6, at least in response to amino acids.
 mRNAs. S6Ks also phosphorylate elongation factor 2 (eEF2) kinase leading to a decrease in its activity at basal calcium concentrations although additional inputs are also involved in its control, such an independent input to the phosphorylation of Ser78, which inhibits the binding of CaM to eEF2 kinase (see text). Inactivation of eEF2 kinase leads to the dephosphorylation of eEF2 and the activation of elongation. Phosphorylation of 4E-BP1 at certain sites leads to its release from eIF4E, which can then interact with eIF4G to form initiation complexes than recruit the 40S ribosomal subunit to the 50-end of the mRNA. Amino acids act as positive modulators of mTOR. The mechanisms by which amino acids exert this effect are unclear, but recent evidence suggests this involves sensing of intracellular amino acid levels. Insulin and a range of other stimuli increase the phosphorylation of S6Ks and 4E-BP1. Recent work indicates that this effect is likely mediated through the tuberous sclerosis complex (TSC1/2) [46]. The question mark by the role of S6 phosphorylation in the translation of 50-TOP mRNAs denotes the fact that Tang et al. [47] have recently challenged the prevailing concept that these mRNAs are regulated via S6 kinases/phosphorylation of rpS6, at least in response to amino acids.")
21
Regolazione di eIF4E Figure 1 The roads to eIF4E. The PI3K/AKT/mTOR pathway is activated by growth factors, mitogens and hormones. Nutrients (amino acids, glucose) also activate mTOR. Activation of PI3K initiates a cascade of events: PDK1 activates AKT which phosphorylates TSC2, thereby rendering TSC1/TSC2 complex unstable and inactive. Rheb, the small G protein, is no longer inhibited by the GAP (GTPase-activating protein) activity of TSC2. The AMP-activated protein kinase (AMPK) also phosphorylates and enhances the activity of TSC2 under energy starvation. Through yet uncharacterized mechanisms, Rheb leads to activation of mTOR. mTOR, which consists of a large multi-protein complex contains two known partners, raptor and GbL, that promote the activation of the translational activator S6K and the hierarchical phosphorylation the 4E-BPs. Hyperphosphorylated 4E-BPs release eIF4E, thereby allowing for cap-dependent translation to occur. Activated S6K phosphorylates the ribosomal protein S6 and causes an increase in translation of 5’TOP containing mRNAs. Most TOP mRNAs encode components of the translation machinery such as ribosomal proteins, elongation factors and the poly (A) binding proteins. Rapamycin and its derivatives CCI-779 and RAD001 specifically interact the immunophilin FKBP12. This complex then binds and inhibits mTOR signaling to downstream targets. These compounds are currently being used in clinical trials as anticancer agents. Another signal transduction pathway, the Ras/Raf/ MAP kinase pathway, is also activated by growth factors and stress. Activation of Ras and MEKK leads to a cascade of events such as JNKK and MAPK activation which culminates in Mnk activation. Mnk phosphorylates eIF4E within the eIF4F complex and permits the recruitment of the ribosome to an mRNA at the initiation codon.The dashed lines indicate possible links.
 also activate mTOR. Activation of PI3K initiates a cascade of events: PDK1 activates AKT which phosphorylates TSC2, thereby rendering TSC1/TSC2 complex unstable and inactive. Rheb, the small G protein, is no longer inhibited by the GAP (GTPase-activating protein) activity of TSC2. The AMP-activated protein kinase (AMPK) also phosphorylates and enhances the activity of TSC2 under energy starvation. Through yet uncharacterized mechanisms, Rheb leads to activation of mTOR. mTOR, which consists of a large multi-protein complex contains two known partners, raptor and GbL, that promote the activation of the translational activator S6K and the hierarchical phosphorylation the 4E-BPs. Hyperphosphorylated 4E-BPs release eIF4E, thereby allowing for cap-dependent translation to occur. Activated S6K phosphorylates the ribosomal protein S6 and causes an increase in translation of 5’TOP containing mRNAs. Most TOP mRNAs encode components of the translation machinery such as ribosomal proteins, elongation factors and the poly (A) binding proteins. Rapamycin and its derivatives CCI-779 and RAD001 specifically interact the immunophilin FKBP12. This complex then binds and inhibits mTOR signaling to downstream targets. These compounds are currently being used in clinical trials as anticancer agents. Another signal transduction pathway, the Ras/Raf/ MAP kinase pathway, is also activated by growth factors and stress. Activation of Ras and MEKK leads to a cascade of events such as JNKK and MAPK activation which culminates in Mnk activation. Mnk phosphorylates eIF4E within the eIF4F complex and permits the recruitment of the ribosome to an mRNA at the initiation codon.The dashed lines indicate possible links.")
22
Regolazione eIF4E Fosforilazione su ser 209 da parte di Mnk1 (e Mnk2) correlata con attivazione Mnk interagiscono con eIF4G e sono attivate dalla via delle MAP chinasi Doppio KO per Mnk1 e 2 è normale eIF4E è inibito dall’interazione con 4E-BP (1, 2 e 3) 4E-BP sono inibite da fosforilazione dipendente dalla via di mTOR
 4E-BP sono inibite da fosforilazione dipendente dalla via di mTOR.")
23
Alterazione della traduzione da parte dei virus
Fig. 2. Model for inhibition of cellular protein synthesis by three animal viruses and promotion of viral specific mRNA translation. In the center of the figure is a schematic representation of a typical mammalian mRNA with the cap-dependent translation-initiation complex. Only eukaryotic initiation factor (eIF) 4GI is depicted because it is not yet known whether there are substantial differences in translational activity for the different isoforms or whether they impact on viral infection. Rotavirus nonstructural protein 3 (NSP3) competes with or ejects the binding of poly(A)-binding protein (PABP) with eIF4G, and selectively binds to the rotavirus 30 UTR, depriving cellular mRNAs of crucial PABP and selectively activating viral mRNA translation. Adenovirus 100k protein competes with MAP kinase-interacting kinase 1 (Mnk1) for an overlapping binding site on eIF4G, resulting in reduced phosphorylation of eIF4E and downregulation of cellular mRNA translation. Adenovirus mRNAs contain the tripartite leader 50 UTR that efficiently utilizes the modified cap-initiation complex for translation initiation by ribosome shunting. The poliovirus (a picornavirus) protease 2A-pro cleaves eIF4G, severing the N-terminal third (which binds eIF4E and PABP) from the C-terminal two-thirds (which contain eIF4A, eIF3 and 40S ribosome subunits). Cellular cap eIF4F-dependent mRNA translation is therefore inhibited, whereas the poliovirus internal ribosome entry site (IRES) selectively recruits the C-terminal eIF4G fragment and associated factors to the IRES for translation of poliovirus mRNAs.
 4GI is depicted because it is not yet known whether there are substantial differences in translational activity for the different isoforms or whether they impact on viral infection. Rotavirus nonstructural protein 3 (NSP3) competes with or ejects the binding of poly(A)-binding protein (PABP) with eIF4G, and selectively binds to the rotavirus 30 UTR, depriving cellular mRNAs of crucial PABP and selectively activating viral mRNA translation. Adenovirus 100k protein competes with MAP kinase-interacting kinase 1 (Mnk1) for an overlapping binding site on eIF4G, resulting in reduced phosphorylation of eIF4E and downregulation of cellular mRNA translation. Adenovirus mRNAs contain the tripartite leader 50 UTR that efficiently utilizes the modified cap-initiation complex for translation initiation by ribosome shunting. The poliovirus (a picornavirus) protease 2A-pro cleaves eIF4G, severing the N-terminal third (which binds eIF4E and PABP) from the C-terminal two-thirds (which contain eIF4A, eIF3 and 40S ribosome subunits). Cellular cap eIF4F-dependent mRNA translation is therefore inhibited, whereas the poliovirus internal ribosome entry site (IRES) selectively recruits the C-terminal eIF4G fragment and associated factors to the IRES for translation of poliovirus mRNAs.")
24
mTOR è un regolatore centrale
Figure 1 | Target of rapamycin is a central regulator of cell growth and proliferation in response to environmental and nutritional conditions. Target of rapamycin (TOR) signalling is regulated by growth factors, amino acids, ATP and O2 levels; second messengers (for example, phosphatidic acid); and, possibly, mitochondrial stress. Signalling through TOR seems to regulate several downstream pathways that impinge on cell-cycle progression, translation initiation, transcriptional stress responses, protein stability and survival. Dashed lines indicate pathways that are best described in yeast. ATP, adenosine triphosphate; ASK1, apoptosis-signal-regulating kinase 1; S6K1, ribosomal p70 S6 kinase 1; eIF4E, eukaryotic translation initiation factor 4E; 4E-BP1, eIF4E-binding protein 1
 signalling is regulated by growth factors, amino acids, ATP and O2 levels; second messengers (for example, phosphatidic acid); and, possibly, mitochondrial stress. Signalling through TOR seems to regulate several downstream pathways that impinge on cell-cycle progression, translation initiation, transcriptional stress responses, protein stability and survival. Dashed lines indicate pathways that are best described in yeast. ATP, adenosine triphosphate; ASK1, apoptosis-signal-regulating kinase 1; S6K1, ribosomal p70 S6 kinase 1; eIF4E, eukaryotic translation initiation factor 4E; 4E-BP1, eIF4E-binding protein 1.")
25
Struttura di mTOR Figure 2 | Domain structure of target of rapamycin. The protein consists of a catalytic kinase domain (CD), an FKBP12-rapamycin-binding (FRB) domain, a putative auto-inhibitory domain (‘repressor domain’, RD) near the carboxyl terminus and up to 20 tandemly repeated HEAT (huntingtin, EF3, a subunit of PP2A and TOR) motifs at the amino terminus, as well as FAT (FRAPATM- TRRAP) and FATC (FAT C terminus) domains.
, an FKBP12-rapamycin-binding (FRB) domain, a putative auto-inhibitory domain (‘repressor domain’, RD) near the carboxyl terminus and up to 20 tandemly repeated HEAT (huntingtin, EF3, a subunit of PP2A and TOR) motifs at the amino terminus, as well as FAT (FRAPATM- TRRAP) and FATC (FAT C terminus) domains.")
26
Inibizione della rapamicina
Figure 3 | Regulation of target of rapamycin signalling by raptor. a | Under amino-acid restriction, raptor binds with higher affinity to target of rapamycin (TOR), inactivating the kinase and preventing the phosphorylation of substrates, such as 4E-BP1, which are bound to raptor through TOR signalling (TOS) motifs. b | Under nutrient-replete conditions, raptor binds less tightly to TOR, presumably as a consequence of allosteric changes in TOR structure, which activates the kinase activity of TOR. This facilitates phosphorylation of 4E-BP1 and assembly of the eIF4F complex, shown in Fig. 4. The question remains whether the binding of the rapamycin–FKBP12 complex to TOR directly inhibits kinase activity and therefore mimics nutrient deprivation, as depicted in c, or induces conformational changes in TOR that displace substrates from the catalytic domain, as depicted in d.
, inactivating the kinase and preventing the phosphorylation of substrates, such as 4E-BP1, which are bound to raptor through TOR signalling (TOS) motifs. b | Under nutrient-replete conditions, raptor binds less tightly to TOR, presumably as a consequence of allosteric changes in TOR structure, which activates the kinase activity of TOR. This facilitates phosphorylation of 4E-BP1 and assembly of the eIF4F complex, shown in Fig. 4. The question remains whether the binding of the rapamycin–FKBP12 complex to TOR directly inhibits kinase activity and therefore mimics nutrient deprivation, as depicted in c, or induces conformational changes in TOR that displace substrates from the catalytic domain, as depicted in d.")
27
mTOR è un regolatore centrale
Figure 1. mTOR is a central effector of growth factor and amino acid signals. Mitogenic and growth signals are represented by insulin or insulin-like growth factor (IGF). The cascade of events starts by activation of the receptor through ligand binding, which leads to phosphorylation of the insulin substrate (IRS). The latter, in turn, binds and activates the phosphatidylinositol 3-kinase (PI3-K). This kinase converts phosphatidylinositol (4,5)-bisphosphate (PtdIns[4,5]P2) into phosphatidylinositol (3,4,5)- trisphosphate (PtdIns[3,4,5]P3), which localizes protein kinase B (PKB, also known as Akt) to the membrane, where it can be phosphorylated and activated by 3- phosphoinositide-dependent kinase 1 (PDK1). Activated PKB phosphorylates tumor suppressor protein tuberous sclerosis 2 (TSC2), resulting in inhibition of the tumor suppressor function of the TSC1–TSC2 complex. Rheb, a small guanine nucleotide phosphatase (GTPase) that is inactivated by the GTPase-activating protein (GAP) activity of TSC2, positively modulates the function of mTOR. In turn, mTOR phosphorylates and inhibits 4E-BP1, thereby derepressing eIF4E and facilitating upregulation of protein synthesis, cell size and cell-cycle progression. In parallel, mTOR, together with PDK1, activates S6K1 and S6K2, which are involved in regulating protein synthesis, cell size, cell-cycle progression, glucose homeostasis and survival of newborns. Activation of mTOR by amino acids is carried out by an ill-defined mechanism. TOP mRNAs, which contain a 50-terminal oligopyrimidine tract and are translationally activated by growth factors and amino acids, direct enhanced synthesis of components of the translational apparatus (including ribosomal proteins and elongation factors among others). The role of mTOR in this mode of regulation remains questionable, however, because inhibition of mTOR by rapamycin leads to conflicting results regarding the translational repression of TOP mRNAs. Arrows indicate activation, bars indicate inhibition, and question marks indicate unknown or questionable pathways.
. The cascade of events starts by activation of the receptor through ligand binding, which leads to phosphorylation of the insulin substrate (IRS). The latter, in turn, binds and activates the phosphatidylinositol 3-kinase (PI3-K). This kinase converts phosphatidylinositol (4,5)-bisphosphate (PtdIns[4,5]P2) into phosphatidylinositol (3,4,5)- trisphosphate (PtdIns[3,4,5]P3), which localizes protein kinase B (PKB, also known as Akt) to the membrane, where it can be phosphorylated and activated by 3- phosphoinositide-dependent kinase 1 (PDK1). Activated PKB phosphorylates tumor suppressor protein tuberous sclerosis 2 (TSC2), resulting in inhibition of the tumor suppressor function of the TSC1–TSC2 complex. Rheb, a small guanine nucleotide phosphatase (GTPase) that is inactivated by the GTPase-activating protein (GAP) activity of TSC2, positively modulates the function of mTOR. In turn, mTOR phosphorylates and inhibits 4E-BP1, thereby derepressing eIF4E and facilitating upregulation of protein synthesis, cell size and cell-cycle progression. In parallel, mTOR, together with PDK1, activates S6K1 and S6K2, which are involved in regulating protein synthesis, cell size, cell-cycle progression, glucose homeostasis and survival of newborns. Activation of mTOR by amino acids is carried out by an ill-defined mechanism. TOP mRNAs, which contain a 50-terminal oligopyrimidine tract and are translationally activated by growth factors and amino acids, direct enhanced synthesis of components of the translational apparatus (including ribosomal proteins and elongation factors among others). The role of mTOR in this mode of regulation remains questionable, however, because inhibition of mTOR by rapamycin leads to conflicting results regarding the translational repression of TOP mRNAs. Arrows indicate activation, bars indicate inhibition, and question marks indicate unknown or questionable pathways.")
28
Complessi formati da mTOR
Figure 2. Model of the mTOR Signaling Network in Mammalian Cells The mTOR signaling network consists of two major branches, each mediated by a specific mTOR complex (mTORC). Rapamycin-sensitive mTORC1 controls several pathways that collectively determine the mass (size) of the cell. Rapamycin- insensitive mTORC2 controls the actin cytoskeleton and thereby determines the shape of the cell. mTORC1 and possibly mTORC2 respond to growth factors (insulin/IGF), energy status of the cell, nutrients (amino acids), and stress. mTORC1 (and likely mTORC2) are multimeric, although are drawn as monomers. Arrows represent activation, whereas bars represent inhibition.
. Rapamycin-sensitive mTORC1 controls several pathways that collectively determine the mass (size) of the cell. Rapamycin- insensitive mTORC2 controls the actin cytoskeleton and thereby determines the shape of the cell. mTORC1 and possibly mTORC2 respond to growth factors (insulin/IGF), energy status of the cell, nutrients (amino acids), and stress. mTORC1 (and likely mTORC2) are multimeric, although are drawn as monomers. Arrows represent activation, whereas bars represent inhibition.")
29
Bersagli di mTOR

30
mTOR regola l'inizio e l'allungamento
Fig. 1. mTOR regulates the phosphorylation and function of several proteins involved in mRNA translation. Phosphorylation of S6Ks leads to their activation. S6Ks phosphorylate rpS6 and is thought to play a role in the regulation of the translation of the subset of mRNAs containing a 50- terminal oligopyrimidine tract (TOP) mRNAs. S6Ks also phosphorylate elongation factor 2 (eEF2) kinase leading to a decrease in its activity at basal calcium concentrations although additional inputs are also involved in its control, such an independent input to the phosphorylation of Ser78, which inhibits the binding of CaM to eEF2 kinase (see text). Inactivation of eEF2 kinase leads to the dephosphorylation of eEF2 and the activation of elongation. Phosphorylation of 4E-BP1 at certain sites leads to its release from eIF4E, which can then interact with eIF4G to form initiation complexes than recruit the 40S ribosomal subunit to the 50-end of the mRNA. Amino acids act as positive modulators of mTOR. The mechanisms by which amino acids exert this effect are unclear, but recent evidence suggests this involves sensing of intracellular amino acid levels. Insulin and a range of other stimuli increase the phosphorylation of S6Ks and 4E-BP1. Recent work indicates that this effect is likely mediated through the tuberous sclerosis complex (TSC1/2) [46]. The question mark by the role of S6 phosphorylation in the translation of 50-TOP mRNAs denotes the fact that Tang et al. [47] have recently challenged the prevailing concept that these mRNAs are regulated via S6 kinases/phosphorylation of rpS6, at least in response to amino acids.
 mRNAs. S6Ks also phosphorylate elongation factor 2 (eEF2) kinase leading to a decrease in its activity at basal calcium concentrations although additional inputs are also involved in its control, such an independent input to the phosphorylation of Ser78, which inhibits the binding of CaM to eEF2 kinase (see text). Inactivation of eEF2 kinase leads to the dephosphorylation of eEF2 and the activation of elongation. Phosphorylation of 4E-BP1 at certain sites leads to its release from eIF4E, which can then interact with eIF4G to form initiation complexes than recruit the 40S ribosomal subunit to the 50-end of the mRNA. Amino acids act as positive modulators of mTOR. The mechanisms by which amino acids exert this effect are unclear, but recent evidence suggests this involves sensing of intracellular amino acid levels. Insulin and a range of other stimuli increase the phosphorylation of S6Ks and 4E-BP1. Recent work indicates that this effect is likely mediated through the tuberous sclerosis complex (TSC1/2) [46]. The question mark by the role of S6 phosphorylation in the translation of 50-TOP mRNAs denotes the fact that Tang et al. [47] have recently challenged the prevailing concept that these mRNAs are regulated via S6 kinases/phosphorylation of rpS6, at least in response to amino acids.")
31
Translation and oncogenesis
Hypertrophic nucleoli were a prominent feature of malignant cell Ribosome alterations are associated with malignancies Common cancer-related mutations are found in pathways feeding into the translation machinery Translation initiation factors are frequently amplified or dysregulated in tumours

32
Alterazione della traduzione e tumori
Figure 1 Role of protein translation control in tumorigenesis. (a) In a first scenario, the ribosome is structurally normal, but mRNA translation is abnormal because of aberrant signaling (e.g. sustained mTOR activation), leading to deregulated or excessive mRNA translation. (b) Ribosome-driven tumorigenesis. In a second scenario, a malfunctioning ribosome may directly contribute to cancer pathogenesis by affecting either globally or specifically mRNA translation
 In a first scenario, the ribosome is structurally normal, but mRNA translation is abnormal because of aberrant signaling (e.g. sustained mTOR activation), leading to deregulated or excessive mRNA translation. (b) Ribosome-driven tumorigenesis. In a second scenario, a malfunctioning ribosome may directly contribute to cancer pathogenesis by affecting either globally or specifically mRNA translation.")
33
Role of ribosome in tumorigenesis tumorigenesis
defective ribosome QUANTITATIVE/QUALITATIVE ALTERATIONOF TRANSLATION tumorigenesis

34
Translation initiation factors and cancers

35
Traduzione e cancro

36
Regolazione della traduzione
generale specifica

37
Transcriptional control Translational control
gene mRNA protein Slow Lasting Transcription-dependent Rapid Short-lived Transcription-independent

38
Indicazione di controllo traduzionale
northern western -Fe +Fe -Fe +Fe mRNA ferritina actina

39
METODI PER STUDIARE IL CONTROLLO TRADUZIONALE
Confronto tra la variazione della quantità di mRNA e quella del prodotto proteico gel poliacrilammide 2D (DIGE) western blot Analisi della stabilità delle proteine mediante l'uso di inibitori della traduzione Analisi della quantità di mRNA sui polisomi (distribuzione)
 western blot. Analisi della stabilità delle proteine mediante l uso di inibitori della traduzione. Analisi della quantità di mRNA sui polisomi (distribuzione)")
40
Polysome analysis

41
TOP mRNA translational control in cultured cells

42
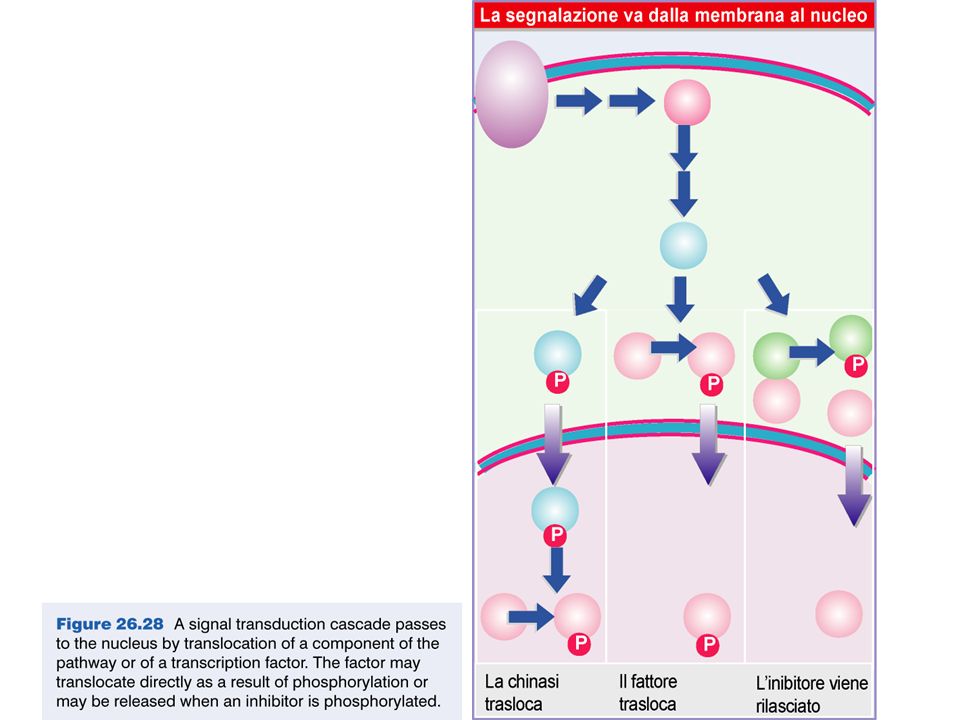
Meccanismi di regolazione traduzionale

43
Meccanismi di regolazione traduzionale

Presentazioni simili







>")










>")
 Piccole molecole di RNA (20-22 nt)>")
