Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
In Natura si distingue:
Vita saprofita: nell’ambiente a spese di materiale inanimato Vita parassitaria: a carico di organismi superiori Parassiti Obbligati Facoltativi (Maggioranza dei batteri) (Minoranza) Il tipo di rapporto nel parassitismo può essere di diverso tipo: simbionti (utili all’organismo parassitato), commensali (indifferenti) e patogeni (dannosi) 1 1
 (Minoranza) Il tipo di rapporto nel parassitismo può essere di diverso tipo: simbionti (utili all’organismo parassitato), commensali (indifferenti) e patogeni (dannosi)")
2
Parassiti Rapporti tra batteri e ospite: Commensali (indifferenti)
Simbionti (utili) Opportunisti (talvolta possono dare malattia) Patogeni (nocivi) (moltiplicazione in vivo e tossigenicità) Simbionti: produzione di vitamine, commensali della cute. Opportunisti Pseudomonas, Candida.I Patogeni non sono in grado di dare malattia in tutte le specie, ma solo in alcune o solo in 1. I batteri a circolazione interumana che danno infezioni esogene sono di norma batteri patogeni obbligati, ossia in grado di provocare un’infezione tutte le volte che raggiungono un organismo sano e non immune. La loro presenza nell’organismo è legata ad un processo infettivo anche se non sempre si hanno manifestazioni morbose (portatori sani che fungono però da reservoir di infezione). Malattia lieve, cronica che non porta rapidemente a morte l’ospite è importante per facilitare la disseminazione del germe. Diversa è la situazione per le infezioni endogene: le malattie sono la conseguenza di 1- dislocazione accidentale di un germe da una zona ad un’altra es E.coli intestino-apparato urinario 2- diminuzione delle difese locali es Clostridi nell’intestino Definizione di batterio patogeno Germe in grado di invadere i tessuti e di moltiplicarsi, danneggiando le funzioni dell’ospite con produzione di una o più sostanze tossiche Potere patogeno: Capacità di moltiplicarsi in vivo Tossigenicità Virulenza: misura del grado di patogenicità 2 2
 Opportunisti (talvolta possono dare malattia) Patogeni (nocivi) (moltiplicazione in vivo e tossigenicità) Simbionti: produzione di vitamine, commensali della cute. Opportunisti Pseudomonas, Candida.I Patogeni non sono in grado di dare malattia in tutte le specie, ma solo in alcune o solo in 1. I batteri a circolazione interumana che danno infezioni esogene sono di norma batteri patogeni obbligati, ossia in grado di provocare un’infezione tutte le volte che raggiungono un organismo sano e non immune. La loro presenza nell’organismo è legata ad un processo infettivo anche se non sempre si hanno manifestazioni morbose (portatori sani che fungono però da reservoir di infezione). Malattia lieve, cronica che non porta rapidemente a morte l’ospite è importante per facilitare la disseminazione del germe. Diversa è la situazione per le infezioni endogene: le malattie sono la conseguenza di 1- dislocazione accidentale di un germe da una zona ad un’altra es E.coli intestino-apparato urinario 2- diminuzione delle difese locali es Clostridi nell’intestino. Definizione di batterio patogeno. Germe in grado di invadere i tessuti e di moltiplicarsi, danneggiando le funzioni dell’ospite con produzione di una o più sostanze tossiche. Potere patogeno: Capacità di moltiplicarsi in vivo. Tossigenicità. Virulenza: misura del grado di patogenicità")
3
Popolazione microbica normale
Esistono due gruppi di PMN: residente transito 1) Competizione con nutrienti (interferenza) 2) Competizione con i recettori sulle cellule (tropismo) 3) Produzione di batteriocine 4) Produzione di acidi grassi o altri metaboliti (anaerobi) 5) Continua stimolazione del sistema immune (macrofagi) 6) Stimolazione di fattori immuni protettivi (Ig) 3 3 3 3
 Competizione con nutrienti (interferenza) 2) Competizione con i recettori sulle cellule (tropismo) 3) Produzione di batteriocine. 4) Produzione di acidi grassi o altri metaboliti (anaerobi) 5) Continua stimolazione del sistema immune (macrofagi) 6) Stimolazione di fattori immuni protettivi (Ig)")
4
POPOLAZIONE MICROBICA NORMALE
Microrganismi acquisiti dalla nascita subiscono modifiche in numero e specie lungo tutto il decorso della vita Dipendono da fattori genetici abitudini alimentari e igieniche situazioni di stress fenomeni psicosomatici età Il corpo umano, come quello di tutti gli esseri viventi, viene normalmente colonizzato da microrganismi che sono acquisiti fin dalla nascita (già al contatto con il tratto genito-urinario della madre) e che subiscono modifiche e variazioni in numero e specie lungo tutto il decorso della vita. Esistono numerosi elementi che incidono sulla composizione e sulla qualità delle specie microbiche presenti che includono fattori genetici, le abitudini alimentari, igieniche, situazioni di stress o fenomeni psicosomatici 4 4 4
 e che subiscono modifiche e variazioni in numero e specie lungo tutto il decorso della vita. Esistono numerosi elementi che incidono sulla composizione e sulla qualità delle specie microbiche presenti che includono fattori genetici, le abitudini alimentari, igieniche, situazioni di stress o fenomeni psicosomatici")
5
POPOLAZIONE MICROBICA NORMALE
Residenti, o popolazione microbica normale rappresentano circa l’1% del peso corporeo. 10 volte il numero totale delle cellule somatiche Intestino: più di 500 (1000) specie diverse di batteri molte delle quali non coltivabili con le metodiche tradizionali Si stima che circa il 60% del materiale fecale sia costituito da batteri. Questi batteri presenti costantemente sono chiamati residenti, o popolazione microbica normale e quantitativamente rappresentano circa l’1% del peso corporeo. Dal punto di vista numerico superano di almeno 10 volte il numero totale delle cellule somatiche. L’apparato gastro enterico possiede in termini quantitativi e qualitativi la maggior parte di tutti i germi veicolati dal corpo. Vi possono essere più di 500 specie diverse di batteri, molte delle quali non coltivabili con le metodiche tradizionale e si stima che circa il 60% del materiale fecale, sia costituito da batteri. 5 5 5
 specie diverse di batteri molte delle quali non coltivabili con le metodiche tradizionali. Si stima che circa il 60% del materiale fecale sia costituito da batteri. Questi batteri presenti costantemente sono chiamati residenti, o popolazione microbica normale e quantitativamente rappresentano circa l’1% del peso corporeo. Dal punto di vista numerico superano di almeno 10 volte il numero totale delle cellule somatiche. L’apparato gastro enterico possiede in termini quantitativi e qualitativi la maggior parte di tutti i germi veicolati dal corpo. Vi possono essere più di 500 specie diverse di batteri, molte delle quali non coltivabili con le metodiche tradizionale e si stima che circa il 60% del materiale fecale, sia costituito da batteri")
6
PMN Cute Cavità orale e prime vie aeree Tratto intestinale Uretra
Vagina Occhio La cute e le mucose ospitano una grande quantità di microorganismi che possono essere distinti in residenti e transitori 6 6

7
PMN Cute PM transitoria Pieghe cutanee Secrezioni sebacee
Sudorazione e lavaggi Disinfezione Medicazioni occlusive 7 7

8
PMN cavità orale Sterile alla nascita
4-12 ore dopo si stabiliscono i viridanti (madre) Seguono stafilococchi, anaerobi, neisserie, moraxelle, difteroidi. Eruzione dei denti: spirochete anaerobie, prevotelle, fusobatteri, capnocytophaga Actinomyces tonsille e gengive degli adulti Miceti: Candida 8 8
 Seguono stafilococchi, anaerobi, neisserie, moraxelle, difteroidi. Eruzione dei denti: spirochete anaerobie, prevotelle, fusobatteri, capnocytophaga. Actinomyces tonsille e gengive degli adulti. Miceti: Candida")
9
Bronchioli e alveoli sterili
PMN vie respiratorie Faringe e trachea simili alla cavità orale Bronchi pochi batteri Bronchioli e alveoli sterili 9 9

10
PMN vie respiratorie 10 10

11
POPOLAZIONE MICROBICA NORMALE
La popolazione microbica dell’apparato gastro-enterico è stata definita come "un insieme di microrganismi presenti nel lume intestinale i quali, se convivono in un determinato equilibrio contribuiscono allo stato di salute dell'ospite" 11 11

12
POPOLAZIONE MICROBICA NORMALE
Nello stomaco vi è un alto grado di acidità e il numero dei microrganismi non supera le mille unità, mentre nell'ileo la concentrazione aumenta arrivando nel colon a batteri per g Nello stomaco ove vi è un alto grado di acidità il numero dei microrganismi non supera le mille unità, mentre nell'ileo la concentrazione aumenta arrivando nel colon a batteri per g (Simon, 1986). Essi portano milioni di geni, un numero di gran lunga superiore ai stimati del genoma umano. Il ruolo di questa enorme informazione genetica e quindi la produzione di enzimi per utilizzare i più svariati alimenti è ben evidente nella varietà di composti che possono essere metabolizzati e, quando questa varia, si modifica anche l'assorbimento dei nutrienti da parte dell'organismo, una situazione accertata nelle persone obese e magre che hanno profili batterici radicalmente diversi nei loro intestini 12 12 12
. Essi portano milioni di geni, un numero di gran lunga superiore ai stimati del genoma umano. Il ruolo di questa enorme informazione genetica e quindi la produzione di enzimi per utilizzare i più svariati alimenti è ben evidente nella varietà di composti che possono essere metabolizzati e, quando questa varia, si modifica anche l assorbimento dei nutrienti da parte dell organismo, una situazione accertata nelle persone obese e magre che hanno profili batterici radicalmente diversi nei loro intestini")
13
PMN intestinale Duodeno: 103-106/gr Digiuno-ileo: 105-108/gr
pH acido Duodeno: /gr Digiuno-ileo: /gr Cieco-colon trasverso: /gr Sigma-retto: 1011/gr antibiotici La cute e le mucose ospitano una grande quantità di microorganismi che possono essere distinti in residenti e transitori basico 13 13 13 13

14
POPOLAZIONE MICROBICA NORMALE
Anche il numero e il tipo di specie di microorganismi varia con il variare del sito anatomico 14 14

15
PMN intestinale 15 15 15 15

16
POPOLAZIONE MICROBICA NORMALE
Portano milioni di geni (5), un numero di gran lunga superiore ai stimati del genoma umano. Una gran varietà di composti che possono essere metabolizzati (è un organo aggiunto, bioreattore) Quando varia, si modifica anche l'assorbimento dei nutrienti Obesi e magri hanno profili batterici radicalmente diversi nei loro intestini Nello stomaco ove vi è un alto grado di acidità il numero dei microrganismi non supera le mille unità, mentre nell'ileo la concentrazione aumenta arrivando nel colon a batteri per g (Simon, 1986). Essi portano milioni di geni, un numero di gran lunga superiore ai stimati del genoma umano. Il ruolo di questa enorme informazione genetica e quindi la produzione di enzimi per utilizzare i più svariati alimenti è ben evidente nella varietà di composti che possono essere metabolizzati e, quando questa varia, si modifica anche l'assorbimento dei nutrienti da parte dell'organismo, una situazione accertata nelle persone obese e magre che hanno profili batterici radicalmente diversi nei loro intestini 16 16 16
, un numero di gran lunga superiore ai stimati del genoma umano. Una gran varietà di composti che possono essere metabolizzati (è un organo aggiunto, bioreattore) Quando varia, si modifica anche l assorbimento dei nutrienti. Obesi e magri hanno profili batterici radicalmente diversi nei loro intestini. Nello stomaco ove vi è un alto grado di acidità il numero dei microrganismi non supera le mille unità, mentre nell ileo la concentrazione aumenta arrivando nel colon a batteri per g (Simon, 1986). Essi portano milioni di geni, un numero di gran lunga superiore ai stimati del genoma umano. Il ruolo di questa enorme informazione genetica e quindi la produzione di enzimi per utilizzare i più svariati alimenti è ben evidente nella varietà di composti che possono essere metabolizzati e, quando questa varia, si modifica anche l assorbimento dei nutrienti da parte dell organismo, una situazione accertata nelle persone obese e magre che hanno profili batterici radicalmente diversi nei loro intestini")
17
POPOLAZIONE MICROBICA NORMALE
La degradazione nell’intestino crasso, in particolar modo della lignina e della cellulosa, è incompleta portando a particelle di fibre vegetali che persistono fino all’intestino distale dove possono esercitare un ruolo che favorisce il processo di eliminazione delle feci 17 17

18
POPOLAZIONE MICROBICA NORMALE
E’ importante il ruolo dei batteri intestinali nella sintesi e nell'utilizzo delle vitamine. B1, B2, B6, B12, PP, H, acido pantotenico e acido folico. La vitamina K viene sintetizzata dall'Eucobacterium lentum che modifica degli steroidi a livello intestinale I germi intestinali interagiscono con gli acidi biliari. E’ importante il ruolo dei batteri intestinali nella sintesi e nell'utilizzo delle vitamine (Lebonlanger et al.1981). Le vitamine sintetizzate vengono utilizzate sia dagli stessi batteri autoctoni e sia dall'ospite, che le assimila insieme a quelle assunte con la dieta per il proprio fabbisogno. E’ ciò che accade con le vitamine B1, B2, B6, B12, PP, H, acido pantotenico e acido folico. La vitamina K prodotta nell'intestino è sufficiente per l'intero fabbisogno dell'ospite e viene sintetizzata dall'Eucobacterium lentum che modifica degli steroidi a livello intestinale (Meier et al., 1970). Inoltre, i germi intestinali interagiscono con gli acidi biliari (Aldini R., et al., 1979). 18 18 18
. Le vitamine sintetizzate vengono utilizzate sia dagli stessi batteri autoctoni e sia dall ospite, che le assimila insieme a quelle assunte con la dieta per il proprio fabbisogno. E’ ciò che accade con le vitamine B1, B2, B6, B12, PP, H, acido pantotenico e acido folico. La vitamina K prodotta nell intestino è sufficiente per l intero fabbisogno dell ospite e viene sintetizzata dall Eucobacterium lentum che modifica degli steroidi a livello intestinale (Meier et al., 1970). Inoltre, i germi intestinali interagiscono con gli acidi biliari (Aldini R., et al., 1979)")
19
POPOLAZIONE MICROBICA NORMALE
Gli acidi biliari derivano dal metabolismo del colesterolo nel fegato, coniugati con glicina ed eliminati con la bile. Il metabolismo degli ormoni steroidei (androgeni, estrogeni e corticosteroidi) avviene nel lume intestinale attraverso varie reazioni chimiche Gli ormoni steroidei vengono modificati a livello del fegato ed escreti con la bile. Nell'intestino verranno deconiugati e sottoposti alle modificazioni enzimatiche per poi ritornare al fegato attraverso la vena porta. 19 19
 avviene nel lume intestinale attraverso varie reazioni chimiche. Gli ormoni steroidei vengono modificati a livello del fegato ed escreti con la bile. Nell intestino verranno deconiugati e sottoposti alle modificazioni enzimatiche per poi ritornare al fegato attraverso la vena porta")
20
POPOLAZIONE MICROBICA INTESTINALE
A questo livello vi è una barriera protettiva costituita da uno strato di muco denso che contiene IgA secretorie, peptidi antimicrobici e anche complessi giunzionali che tengono unite le cellule epiteliali adiacenti in modo da regolare la permeabilità del tessuto intestinale e impedire che i microrganismi lascino questa sede 20 20

21
POPOLAZIONE MICROBICA NORMALE
Gli enterociti esprimono recettori PAMP (pathogen-associated molecular pattern), comprese le proteine denominate TLR e NOD. Questi recettori captano la presenza di componenti batterici presenti e conservati in diverse specie microbiche e possono dare inizio ad una cascata pro infiammatoria per la difesa. I batteri possono presentare sulla superficie esterna delle strutture che sono individuate dalle cellule epiteliali e dalle cellule del sistema immunitario Gli enterociti esprimono recettori PAMP (pathogen-associated molecular pattern), comprese le proteine denominate TLR e NOD. Questi recettori captano la presenza di componenti batterici presenti e conservati in diverse specie microbiche e possono dare inizio ad una cascata pro infiammatoria per la difesa. I batteri possono presentare sulla superficie esterna delle strutture che sono individuate dalle cellule epiteliali e dalle cellule del sistema immunitario, queste Al fine di discriminare tra patogeni e commensali, queste molecole sono posizionate all’interno delle cellule o lateralmente sulla parete cellulare o in limitato numero su enterociti non stimolati. 21 21 21
, comprese le proteine denominate TLR e NOD. Questi recettori captano la presenza di componenti batterici presenti e conservati in diverse specie microbiche e possono dare inizio ad una cascata pro infiammatoria per la difesa. I batteri possono presentare sulla superficie esterna delle strutture che sono individuate dalle cellule epiteliali e dalle cellule del sistema immunitario. Gli enterociti esprimono recettori PAMP (pathogen-associated molecular pattern), comprese le proteine denominate TLR e NOD. Questi recettori captano la presenza di componenti batterici presenti e conservati in diverse specie microbiche e possono dare inizio ad una cascata pro infiammatoria per la difesa. I batteri possono presentare sulla superficie esterna delle strutture che sono individuate dalle cellule epiteliali e dalle cellule del sistema immunitario, queste Al fine di discriminare tra patogeni e commensali, queste molecole sono posizionate all’interno delle cellule o lateralmente sulla parete cellulare o in limitato numero su enterociti non stimolati")
22
The commensal microbiota, intestinal epithelial cells, and intestinal immune cells engage in a complex crosstalk. Epithelial cells, M cells, and dendritic cells (DCs) can directly sense and sample the intestinal contents and communicate information about the microbiota to other subsets of immune cells. Toll-like receptors, expressed by epithelial cells, M cells and DCs, and NOD-like receptors, are classes of microbe-sensing molecules. Cytokines, chemokines, and host and microbial metabolites are key molecular mediators of intestinal homeostasis that influence responses of both host and microbe. l 22 22

23
IMPORTANTE IL LIVELLO DEGLI ESTROGENI
PMN vagina Alla nascita pH acido: lattobacilli Dopo alcune settimane fino a pubertà pH neutro: PMN mista cocchi e bacilli Pubertà pH acido: lattobacilli Dopo la menopausa pH neutro: PMN mista cocchi e bacilli IMPORTANTE IL LIVELLO DEGLI ESTROGENI 23 23

24
b) Gram stain of Lactobacillus acidophilus, the predominant organism in the vagina of women between the onset of puberty and the end of menopause. Individual rods are 3–4 µm long. The genitourinary tract of older and younger women is less acidic and is populated by a much more heterogeneous group of microorganisms that grow at neutral to slightly alkaline pH. 24 24

25
25 25

26
Batteri a circolazione interumana:
Infezioni esogene ed endogene Portatore sano 26 26

27
Possibili vie d’accesso per i microorganismi
Canali naturali Vie respiratorie tratto gastro-enterico genito-urinario 27 27

28
Possibili vie d’accesso per i microorganismi
Via Esempi Ingestione Salmonella, Shigella, E. coli, V. cholerae, C. botulinum ecc.. Inalazione Mycobacterium, Mycoplasma, Legionella Chlamydia ecc… Penetrazione diretta (aghi, ferite) C. tetani, S. aureus ecc.. Punture d’artropodi Rickettsia, Francisella, Y. Pestis ecc.. Sessuale N. gonorrhoeae, T. pallidum, C. trachomatis transplacentare T. pallidum 28 28
 C. tetani, S. aureus ecc.. Punture d’artropodi. Rickettsia, Francisella, Y. Pestis ecc.. Sessuale. N. gonorrhoeae, T. pallidum, C. trachomatis. transplacentare. T. pallidum")
29
Inserzioni di: cateteri (urinari, vascolari ecc.)
Possibili vie d’accesso per i microorganismi Canali occasionali Inserzioni di: cateteri (urinari, vascolari ecc.) tubi per ventilazione sondini di drenaggio Somministrazione di farmaci che deprimono la risposta immunitaria o che alterano il microambiente di alcuni distretti dell’organismo (es. antiacidi) interruzioni dell’integrità della superficie della mucosa e della pelle 29 29
 tubi per ventilazione. sondini di drenaggio. Somministrazione di farmaci che deprimono la risposta immunitaria o che alterano il microambiente di alcuni distretti dell’organismo (es. antiacidi) interruzioni dell’integrità della superficie della mucosa e della pelle")
30
Meccanismi di difesa naturale e barriere
L’ambiente in vivo è molto sfavorevole per i batteri: pH, T, O2, flusso dei liquidi organici, cellule ciliate, macrofagi, integrità e composizione biochimica dei tessuti, infiammazione, febbre Strato corneo della pelle 30 30

31
Meccanismi di difesa naturale e barriere
Colonizzazione della cute e delle mucose Meccanismi di difesa naturale e barriere Mucose: muco, lisozima, IgA, epiteli ciliati, movimento del contenuto del lume del canale mucoso (intestino, uretra)… PMN residente Cellule di sfaldamento 31 31
… PMN residente. Cellule di sfaldamento")
32
Meccanismi di difesa naturale
Fagocitosi Funzioni dei fagociti: chemiotassi, ingestione e uccisione. I batteri elaborano sostanze che attraggono i fagociti, la fagocitosi è favorita dalle opsonine-anticorpi che rivestono la superficie batterica a) l’anticorpo agisce da opsonina b) l’anticorpo + antigene attiva il complemento c) opsonine possono essere prodotte da sistemi termolabili che attivano il complemento La fagocitosi richiede consumo di energia. E’ posseduta dai granulociti e macrofagi, il meccanismo non è del tutto noto, è stato osservato un aumento della produzione di acqua ossigenata e liberazione di lisozima nei vacuoli (fagosoma, proteine antibatteriche, lisozima, pH, fosfatasi, lactoferrina, perossidasi) 32 32
 l’anticorpo agisce da opsonina. b) l’anticorpo + antigene attiva il complemento. c) opsonine possono essere prodotte da sistemi termolabili che attivano il complemento. La fagocitosi richiede consumo di energia. E’ posseduta dai granulociti e macrofagi, il meccanismo non è del tutto noto, è stato osservato un aumento della produzione di acqua ossigenata e liberazione di lisozima nei vacuoli (fagosoma, proteine antibatteriche, lisozima, pH, fosfatasi, lactoferrina, perossidasi)")
33
33 33

34
Resistenza al sistema immune (fagocitosi, Ig)
Strategie batteriche Resistenza al sistema immune (fagocitosi, Ig) Variazione antigenica (shift) Inibizione della fusione lisosomiale Elusione dei lisosomi e moltiplicazione nel citoplasma Produzione di catalasi che scinde lo ione superossido Resistenza agli enzimi lisosomiali Produzione di IgA proteasi 34 34
 Variazione antigenica (shift) Inibizione della fusione lisosomiale. Elusione dei lisosomi e moltiplicazione nel citoplasma. Produzione di catalasi che scinde lo ione superossido. Resistenza agli enzimi lisosomiali. Produzione di IgA proteasi")
35
Resistenza al sistema immune (fagocitosi, Ig)
Strategie batteriche Resistenza al sistema immune (fagocitosi, Ig) La resistenza ai macrofagi può aversi mediante: 1. produzione di leucocidine che uccidono i macrofagi 2. capsula che impedisce la fagocitosi 3. produzione di superossidismutasi che sequestra l’ossigeno che serve ai macrofagi per la produzione di ATP che è necessario per la fagocitosi 4. mimetismo antigenico 35 35
 La resistenza ai macrofagi può aversi mediante: 1. produzione di leucocidine che uccidono i macrofagi. 2. capsula che impedisce la fagocitosi. 3. produzione di superossidismutasi che sequestra l’ossigeno che serve ai macrofagi per la produzione di ATP che è necessario per la fagocitosi. 4. mimetismo antigenico")
36
Mimetismo antigene È la presenza di componenti strutturali che presentano notevoli analogie di composizione con materiali presenti nei tessuti dell’organismo ospite che vengono quindi difficilmente individuati come non- self Il batterio può quindi sfuggire, almeno nella fase iniziale della invasione dei tessuti, alla difesa immunitaria. Esempi: la capsula di S. pyogenes è formata da acido jaluronico, presnte nel connettivo. Alcuni enterobatteri possiedono nella porzione polisaccaridica del LPS determinanti antigeni simili a quelli presenti nelle emazie della specie normalmente parassitata. In alcune circostanze il mimetismo può avere un ruolo nel determinismo della patologia ad esempio S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto). 36 36
 che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto)")
37
Mimetismo antigene S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemento di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Il batterio può quindi sfuggire, almeno nella fase iniziale della invasione dei tessuti, alla difesa immunitaria. Esempi: la capsula di S. pyogenes è formata da acido jaluronico, presnte nel connettivo. Alcuni enterobatteri possiedono nella porzione polisaccaridica del LPS determinanti antigeni simili a quelli presenti nelle emazie della specie normalmente parassitata. In alcune circostanze il mimetismo può avere un ruolo nel determinismo della patologia ad esempio S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto). 37 37
 che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemento di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Il batterio può quindi sfuggire, almeno nella fase iniziale della invasione dei tessuti, alla difesa immunitaria. Esempi: la capsula di S. pyogenes è formata da acido jaluronico, presnte nel connettivo. Alcuni enterobatteri possiedono nella porzione polisaccaridica del LPS determinanti antigeni simili a quelli presenti nelle emazie della specie normalmente parassitata. In alcune circostanze il mimetismo può avere un ruolo nel determinismo della patologia ad esempio S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto)")
38
Mimetismo antigene Le Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto). Il batterio può quindi sfuggire, almeno nella fase iniziale della invasione dei tessuti, alla difesa immunitaria. Esempi: la capsula di S. pyogenes è formata da acido jaluronico, presnte nel connettivo. Alcuni enterobatteri possiedono nella porzione polisaccaridica del LPS determinanti antigeni simili a quelli presenti nelle emazie della specie normalmente parassitata. In alcune circostanze il mimetismo può avere un ruolo nel determinismo della patologia ad esempio S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto). 38 38
. Il batterio può quindi sfuggire, almeno nella fase iniziale della invasione dei tessuti, alla difesa immunitaria. Esempi: la capsula di S. pyogenes è formata da acido jaluronico, presnte nel connettivo. Alcuni enterobatteri possiedono nella porzione polisaccaridica del LPS determinanti antigeni simili a quelli presenti nelle emazie della specie normalmente parassitata. In alcune circostanze il mimetismo può avere un ruolo nel determinismo della patologia ad esempio S.pyogenes possiede diversi materiali antigenici (proteina M, antigeni di membrana, carboidrati di gruppo) che presentano analogie molecolari ed antigeniche con strutture molecolari presenti nel miocardio, cute, nell’encefalo e nelle articolazioni e la risposta autoimmune che si mette in opera durante l’infezione da S. pyogenes, potrebbe rappresentare un elemnto di rilievo nella patogenesi della febbre reumatica a della cardiomiopatia reumatica. Di recente (1999) è stato dimostrato che la Chlamydie presentano negli involucri esterni alcune proteine ricche in cisteina che hanno analogie antigenetiche con la catena pesante dell’alfa-miosina specifica del muscolo cardiaco ed è stata quindi ipotizzata una base autoimmune nella patologia cardiaca correlata alla infezione di Chlamydia (C. pneumoniae soprattutto)")
39
Neisseria meningitidis
Gram-negative commensal of the human oropharynx, can cause septicaemia by crossing the mucosal barrier and entering the bloodstream or meningitis by crossing the blood–brain barrier into the cerebrospinal fluid. In each case, N. meningitidis uses molecular mimicry to survive exposure to the innate immune system. The ability of the immune system to distinguish foreign cells from self is mediated in part by sialic acid, which coats the surface of human cells and acts as a ‘self signal’. N. meningitidis exploits this feature by decorating its outer surface with sialic acid residues, thus masking itself from human defences. Molecular mimicry is a key component of N. meningitidis pathogenesis and mutants that are unable to sialylate their outer surface are highly attenuated. The in vivo nutritional environment has an impact on N. meningitidis host cell mimicry. N. meningitidis can use lactate and glucose as carbon and energy sources, and lactate and glucose are present in nasopharyngeal tissue, serum and cerebrospinal fluid at 1–2 mM and 6–10 mM levels, respectively. 39

40
Molecular mimicry by Neisseria meningitidis.
a | Glucose catabolism in N. meningitidis proceeds by the Entner–Doudoroff pathway and lactate catabolism feeds directly into the sialic acid pathway. Note the relative number of metabolic steps from glucose to phosphoenolpyruvate compared with that from lactate to phosphoenolpyruvate. b | Sialylated lipopolysaccharide (LPS) on the N. meningitidis surface mimics the surface of eukaryotic cells, preventing deposition of the complement molecule C3. Inactivation of the lactate permease gene lctP results in C3-mediated cell lysis. 40
 on the N. meningitidis surface mimics the surface of eukaryotic cells, preventing deposition of the complement molecule C3. Inactivation of the lactate permease gene lctP results in C3-mediated cell lysis. 40.")
41
Neisseria meningitidis
Interestingly, although both of these carbon sources are present in vivo, N. meningitidis catabolizes lactate at a faster rate than it does glucose, and mutants that are deficient for lactate transport are defective colonizers of nasopharyngeal tissue. Why might a bacterium preferentially metabolize a more oxidized substrate such as lactate over glucose? One likely explanation is that intermediates of lactate consumption feed directly into the sialylation pathway, thus enhancing sialic acid biosynthesis. This, in turn, leads to increased sialylation of the N. meningitidis outer membrane. As expected, a N. meningitidis strain that is unable to transport lactate (DlctP) is highly deficient for sialic acid modification of the outer membrane and is more susceptible to complement-mediated killing. Thus, for N. meningitidis, catabolism of a preferred carbon source in vivo is coupled to a unique immune-evasion strategy. 41
 is highly deficient for sialic acid modification of the outer membrane and is more susceptible to complement-mediated killing. Thus, for N. meningitidis, catabolism of a preferred carbon source in vivo is coupled to a unique immune-evasion strategy. 41.")
42
Quando un microorganismo invade l’ospite
E’ in grado di superare le difese di un individuo sano PATOGENO Sfrutta alcune situazioni di debolezza dell’ospite OPPORTUNISTA 42 42

43
Attributi dei microorganismi che causano malattia
Abilità di causare malattia o produrre lesioni progressive (Patogenicità) Grado di patogenicità, ovvero, malattia causata anche da batteri in numero piuttosto limitato (Virulenza). Queste proprietà possono essere suddivise in tossicità: abilità nel produrre sostanze tossiche ed invasività: abilità nel penetrare i tessuti e di diffondere. Molti di questi caratteri sono controllati da plasmidi. 43 43
 Grado di patogenicità, ovvero, malattia causata anche da batteri in numero piuttosto limitato (Virulenza). Queste proprietà possono essere suddivise in tossicità: abilità nel produrre sostanze tossiche ed invasività: abilità nel penetrare i tessuti e di diffondere. Molti di questi caratteri sono controllati da plasmidi")
44
Isole di patogenicità (PAI)
I geni per i fattori di patogenicità, soprattutto se localizzati a livello cromosomico, possono essere (Gram-) riuniti in segmenti di DNA: PAI I PAI probabilmente sono segmenti di DNA acquisiti orizzontalmente che differenziano i batteri che li posseggono dagli altri della stessa specie 44 44
 riuniti in segmenti di DNA: PAI. I PAI probabilmente sono segmenti di DNA acquisiti orizzontalmente che differenziano i batteri che li posseggono dagli altri della stessa specie")
45
Infezione L’infezione è il processo attraverso il
quale un microorganismo entra in relazione con l’ospite 45 45

46
Infezioni Monomicrobica: un singolo patogeno
Polimicrobica: da più patogeni aerobi o anaerobi Mista: aerobi ed anaerobi insieme Comunitaria: acquisita in ambiente non ospedaliero Nosocomiale: acquisita dopo 72 ore dal ricovero 46 46

47
Fasi di una proliferazione batterica
1) Adesione 2) Invasione 3) Produzione di metaboliti (crescita) 4) Produzione di tossine: enzimi degradativi tossine tipo A-B 5) Endotossine 6) Induzione di infiammazione 7) Resistenza al sistema immune (fagocitosi, Ig) 8) Resistenza agli antibiotici 47 47
 Adesione. 2) Invasione. 3) Produzione di metaboliti (crescita) 4) Produzione di tossine: enzimi degradativi. tossine tipo A-B. 5) Endotossine. 6) Induzione di infiammazione. 7) Resistenza al sistema immune (fagocitosi, Ig) 8) Resistenza agli antibiotici")
48
Fattori che caratterizzano un microorganismo patogeno
Adesività mediata da strutture specializzate ac. lipoteicoici nei gram-positivi proteine nei gram-negativi (fimbrie, pili ecc.) produzione di capsula (slime) capacità biochimica di metabolizzare nelle condizioni nutrizionali fornite dai tessuti dell’ospite capacità di penetrare e moltiplicarsi in tessuti profondi (invasività) capacità di contrastare i meccanismi difensivi dell’ospite produzione di esotossine 48 48
 produzione di capsula (slime) capacità biochimica di metabolizzare nelle condizioni. nutrizionali fornite dai tessuti dell’ospite. capacità di penetrare e moltiplicarsi in tessuti profondi (invasività) capacità di contrastare i meccanismi difensivi dell’ospite. produzione di esotossine")
49
Adesione Uno dei principali eventi che promuovono l’infezione.
Il processo è mediato da strutture poste sulla superficie esterna dei batteri note come adesine, per esempio l’antigene K88 dell’Escherichia coli enteropatogeno, gli acidi lipoteicoici (streptococchi) e altre. Le cellule epiteliali hanno a loro volta, come già detto, strutture che legano le adesine ad esempio fibronectina disposta sulla superficie dell’epitelio orale. (Batteri isogenici privi di adesine non danno infezione) 49 49
 e altre. Le cellule epiteliali hanno a loro volta, come già detto, strutture che legano le adesine ad esempio fibronectina disposta sulla superficie dell’epitelio orale. (Batteri isogenici privi di adesine non danno infezione)")
50
3: Fase di adesione reversibile e irreversibile
Perchè i batteri aderiscono? I batteri non adesi verrebbero eliminati Adesione: prima tappa della colonizzazione e della formazione del biofilm 50

51
Evidenze sperimentali del ruolo dell’adesione nella colonizzazione
I batteri legano recettori isolati o analoghi dei recettori L’adesina purificata o un suo analogo lega il recettore L’adesione è inibita da: Analoghi dei recettori e delle adesine Enzimi che distruggono l’adesina o il recettore Anticorpi specifici diretti contro l’adesina o il recettore 51

52
Cosa utilizzano i batteri per aderire? Adesine batteriche
Proteine espresse sulla superficie Fimbrie Glicocalice, Lipopolisaccaride (G-), Acidi teicoici (G+) Enzimi Enzimi batterici ancorano la cellula al substrato Enzimi batterici possono esporre recettori criptici 52
, Acidi teicoici (G+) Enzimi. Enzimi batterici ancorano la cellula al substrato. Enzimi batterici possono esporre recettori criptici. 52.")
53
Morfologia delle adesine
Afimbrial adhesin Type I fimbriae Type IV fimbriae (= bundle forming pilus) Curli 53 53 53
 Curli")
54
ADESINE BATTERICHE 54

55
Adesività Colonizzazione delle mucose Adesione specifica e selettiva alle cellule epiteliali: adesine-recettore G-: proteine in fimbrie o pili G+: glicolipidi (ac. Lipoteicoici, [LTA]) + proteine (es. F di S.pyogenes) in fibrille Recettore su cellule: porzione glicidica di glicoproteine o glicolipidi o proteine di membrana Materiale capsulare ) Colonizzazione delle mucose è il primo step. L’interazione dei batteri patogeni con le cellule degli epiteli mucosi è mediata da fattori localizzati alla superficie della cellula batterica o secreti all’esterno della cellula stessa. Questa caratteristica batterica prende il nome di adesività. Le adesine sono proteine localizzate sull’estremita di fimbrie e pili 55 55
 + proteine (es. F di S.pyogenes) in fibrille. Recettore su cellule: porzione glicidica di glicoproteine o glicolipidi o proteine di membrana. Materiale capsulare. ) Colonizzazione delle mucose è il primo step. L’interazione dei batteri patogeni con le cellule degli epiteli mucosi è mediata da fattori localizzati alla superficie della cellula batterica o secreti all’esterno della cellula stessa. Questa caratteristica batterica prende il nome di adesività. Le adesine sono proteine localizzate sull’estremita di fimbrie e pili")
56
Adesività S. mitis tutta la mucosa buccale
Colonizzazione delle mucose Ogni specie batterica interagisce con recettori presenti su determinati epiteli: Esiste uno spettro S. mitis tutta la mucosa buccale S. salivarius lingua e gengive S. pyogenes faringe e tonsille N. gonorrhoeae uretra V. cholerae intestino tenue E. coli ileo, colon,tratto urogenitale, ecc. Tossine ciliostatiche 56 56

57
Fibronectina Sulla superficie delle cellule epiteliali vi è una proteina, la fibronectina, con cui interagiscono le adesine dei batteri gram-positivi, prevenendo così l’adesione dei gram-negativi La fibronectina è una glicoproteina prodotta dagli epatociti e distribuita in forma solubile in tutti i liquidi organici, viene escreta nelle cavità naturali dell’organismo dove si deposita sulla superficie delle mucose ed è inoltre presente nei tessuti dove forma un reticolo fibrillare La fibronectina è coinvolta in alcuni processi biologici tra cui la fagocitosi, la rigenerazione dei tessuti e nella differenziazione cellulare. 57 57

58
Acquisizione di patogeni potenziali: malattia di base
Durante il decorso di varie affezioni si determinano variazioni nelle cellule di rivestimento (cute e mucose) con colonizzazione da parte dei bacilli gram-negativi e riduzione della popolazione batterica gram-positiva Volontari o pazienti ricoverati in clinica psichiatrica non hanno dimostrato variazioni della popolazione batterica normale dell’orofaringe Variazioni con la durata della degenza (stesso reparto) 58 58
 con colonizzazione da parte dei bacilli gram-negativi e riduzione della popolazione batterica gram-positiva. Volontari o pazienti ricoverati in clinica psichiatrica non hanno dimostrato variazioni della popolazione batterica normale dell’orofaringe. Variazioni con la durata della degenza (stesso reparto)")
59
FIBRONECTINA: ALTRI EFFETTI SULLA POPOLAZIONE BATTERICA RESIDENTE
La presenza di fibronectina in: ferite accidentali o causate da strumenti, sulla superficie di cateteri predispone all’invasività di cocchi gram-positivi (stafilococchi, streptococchi) che possiedono sulla loro superficie recettori per la fibronectina con la quale creano un legame molto stabile. La colonizzazione dei cateteri avviene con batteri gram- positivi produttori di slime che li rende immuni da qualsiasi trattamento con farmaci e dall’azione dei macrofagi. 59 59
 che possiedono sulla loro superficie recettori per la fibronectina con la quale creano un legame molto stabile. La colonizzazione dei cateteri avviene con batteri gram- positivi produttori di slime che li rende immuni da qualsiasi trattamento con farmaci e dall’azione dei macrofagi")
60
Fibronectina: un modulatore della popolazione batterica residente
Pazienti affetti da malattie organiche o ustioni, traumi ecc. dimostrano un ridotto tasso di fibronectina nel plasma Conseguenze Riduzione di fibronectina dalla superficie cellulare scomparsa dei microorganismi (cocchi) gram-positivi colonizzazione da parte di bacilli gram-negativi Produzione di proteasi batterica e dell’ospite? Ulteriore riduzione del tasso di fibronectina nel sito di colonizzazione. 60 60
 gram-positivi colonizzazione da parte di bacilli gram-negativi. Produzione di proteasi batterica e dell’ospite Ulteriore riduzione del tasso di fibronectina nel sito di colonizzazione")
61
Immunità naturale e Immunità acquisita:
dovuta non dal contatto con l’antigene ma dal genotipo. Es. malattie che colpiscono certe specie e non altre. Nell’uomo certe razze, certi gruppi etnici ecc. resistenza individuale diversa, nutrizione età, malattie varie. Immunità acquisita: passiva dovuta alla somministrazione di anticorpi specifici che danno una copertura temporanea, ma può consentire una immediata ed elevata risposta anticorpale. E’ passiva l’immunità acquisita dalla madre attraverso il flusso sanguigno ed il colostro. 61 61

62
Immunità attiva e Immunità umorale
dovuta al contatto con un antigene, produzione di anticorpi richiede tempi di sviluppo anche lungo ma può dare protezione permanente Immunità umorale produzione attiva di anticorpi neutralizzano tossine e prodotti cellulari effetto battericida diretto o litico con il complemento bloccano l’abilità infettiva dell’agente agglutinano i batteri facilita la fagocitosi opsonizzazione, interagiscono con le componenti di superficie che possono limitare la fagocitosi 62 62

63
Sopravvivenza alla superficie delle mucose
Una volta ancorati alla superficie di un epitelio (ma questo anche nei tessuti profondi) i batteri si moltiplicano e interagiscono con le cellule superiori attraverso la produzione di sostanze variamente tossiche. Queste proteine vengono secrete da appositi sistemi secretori grazie al quale i batteri possono traslocare nel citosol della cellula eucariotica le proteine effettrici che interferiscono con il signalling eucariotico, compromettendo le funzioni cellulari e attivando il processo di apoptosi o alterano strutture particolari e causano necrosi Una volta ancorati alla sup di un epitelio (ma questo anche nei tessuti profondi) devono alimentarsi. In linea di principio i m.i. si trovano in un ambiente estremamente favorevole per la loro moltiplicazione (elevato tenore di acqua pH e Temp controllata, tuttavia devono da una parte difendersi dalle difese dell’organismo e dall’altra approvigionarsi di alimenti che possono essere non facilmente raggiungibili . Esempio Fe. Il Fe liberamente utilizzabile è in conc bassissima perchè legato a lattoferrina,transferrina, ferritina e emina e non suff. al metabolismo batterico. I patogeni possiedono quindi dei meccanismi per catare il Fe. : 1- Siderofori, 2-recettori superf. per legare la transferrina e distacco del Fe a livello di membrana (Neisseria) 3- capacità di usare l’ eme come sorgente di Fe, l’emoglobina, emoglobina-aptoglobina, eme-emopessina, eme-albumina (emofilo). 4- produzione di emolisine con rilascio di Fe-eme e Fe-emoglobina. Una volta ancorati alla superficie i batteri si moltiplicano e interagiscono con le cellule superiori attraverso la produzione di sostanze variamente tossiche. Queste proteine vengono secrete da appositi sistemi secretori molto studiato il secIII che funziona come una microsiringa grazie alla quale i batteri traslocano nel citosol della cellula eucariotica le proteine effettrici che interferiscono con il signalling eucariotico, compromettendo le funzioni cellulari e innesco del processo di apoptosi o alterano strutture particolari e causano necrosi 63 63
 i batteri si moltiplicano e interagiscono con le cellule superiori attraverso la produzione di sostanze variamente tossiche. Queste proteine vengono secrete da appositi sistemi secretori grazie al quale i batteri possono traslocare nel citosol della cellula eucariotica le proteine effettrici che interferiscono con il signalling eucariotico, compromettendo le funzioni cellulari e attivando il processo di apoptosi o alterano strutture particolari e causano necrosi. Una volta ancorati alla sup di un epitelio (ma questo anche nei tessuti profondi) devono alimentarsi. In linea di principio i m.i. si trovano in un ambiente estremamente favorevole per la loro moltiplicazione (elevato tenore di acqua pH e Temp controllata, tuttavia devono da una parte difendersi dalle difese dell’organismo e dall’altra approvigionarsi di alimenti che possono essere non facilmente raggiungibili . Esempio Fe. Il Fe liberamente utilizzabile è in conc bassissima perchè legato a lattoferrina,transferrina, ferritina e emina e non suff. al metabolismo batterico. I patogeni possiedono quindi dei meccanismi per catare il Fe. : 1- Siderofori, 2-recettori superf. per legare la transferrina e distacco del Fe a livello di membrana (Neisseria) 3- capacità di usare l’ eme come sorgente di Fe, l’emoglobina, emoglobina-aptoglobina, eme-emopessina, eme-albumina (emofilo). 4- produzione di emolisine con rilascio di Fe-eme e Fe-emoglobina. Una volta ancorati alla superficie i batteri si moltiplicano e interagiscono con le cellule superiori attraverso la produzione di sostanze variamente tossiche. Queste proteine vengono secrete da appositi sistemi secretori molto studiato il secIII che funziona come una microsiringa grazie alla quale i batteri traslocano nel citosol della cellula eucariotica le proteine effettrici che interferiscono con il signalling eucariotico, compromettendo le funzioni cellulari e innesco del processo di apoptosi o alterano strutture particolari e causano necrosi")
64
Invasione Produzione di enzimi extracellulari o attivi all’esterno dei batteri: collagenasi: distrugge il collageno coagulasi: causa la coagulazione del plasma S.aureus ialuronidasi: idrolizza l’acido ialuronico (tessuto connettivo) streptochinasi: (fibrinolisina) promuove la formazione di plasmina dal plasminogeno, dissolve i coaguli emolisine: lisano i globuli rossi proteasi: idrolizzano le immunoglobuline o altre proteine dell’ospite. La tossicità gioca un ruolo importante nell’invasività ma non è un fattore diretto (capsula polisaccaridica o di acidi ialuronici, proteina M, polipeptidi di superficie, resistenza agli enzimi dei fagociti). 64 64
 streptochinasi: (fibrinolisina) promuove la formazione di plasmina dal plasminogeno, dissolve i coaguli. emolisine: lisano i globuli rossi. proteasi: idrolizzano le immunoglobuline o altre proteine dell’ospite. La tossicità gioca un ruolo importante nell’invasività ma non è un fattore diretto (capsula polisaccaridica o di acidi ialuronici, proteina M, polipeptidi di superficie, resistenza agli enzimi dei fagociti)")
65
Penetrazione nei tessuti profondi
Mucosa Sottomucosa Tessuti profondi Via ematica/linfatica Tutto l’organismo Il danneggiamento dell’epitelio mucoso è per molti batteri la strada per raggiungere la sottomucosa, altri quali Salmonelle e Shigelle utilizzano peculiari meccanismi invasivi che consentono di penetrare direttamente nelle cellule dell’epitelio mucoso. 65 65

66
Penetrazione nei tessuti profondi
Il danneggiamento dell’epitelio mucoso è per molti batteri la strada per raggiungere la sottomucosa, altri quali Salmonelle e Shigelle utilizzano peculiari meccanismi invasivi che consentono di penetrare direttamente nelle cellule dell’epitelio mucoso. Il danneggiamento dell’epitelio mucoso è per molti batteri la strada per raggiungere la sottomucosa, altri quali Salmonelle e Shigelle utilizzano peculiari meccanismi invasivi che consentono di penetrare direttamente nelle cellule dell’epitelio mucoso. 66 66

67
Sopravvivenza nella sottomucosa e nei tessuti profondi
Sfuggire alle difese immunitarie (capsula o resistenza al killing dei fagociti) moltiplicazione Tempo di moltiplicazione in vivo (8-10h ?) (situazione simile a chemostato*, ma effetto del s.i.) Siderofori ecc. Tropismo d’organo * sangue e linfa portano nutrimenti e allontanano scorie Una volta introdotti all’interno di fagociti professionali, alcuni batteri patogeni sono in grado di evadere rapidamente dal fagosoma (prima della fusione con i lisosomi), mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno- dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). In alcuni distretti la presenza di certe sostanze che un patogeno può utilizzare a scopo nutrizionale può rappresentare un elemento determinante per favorirne la localizzazione in quel distretto che diviene bersaglio preferenziale es. P. mirabilis provoca inf. Urinarie per la frande capacità di produrre ureasi che gli consente di utilizzare l’urea come sorgente di N. es Brucella spp. Causa nei bovini malattia di modesta gravità nell’animale gravido causa aborto per l’invasione degli annessi fetali che sono ricchi di eritritolo, un carboidrato che per le brucelle è molto utile. 67 67
 moltiplicazione. Tempo di moltiplicazione in vivo (8-10h ) (situazione simile a chemostato*, ma effetto del s.i.) Siderofori ecc. Tropismo d’organo. * sangue e linfa portano nutrimenti e allontanano scorie. Una volta introdotti all’interno di fagociti professionali, alcuni batteri patogeni sono in grado di evadere rapidamente dal fagosoma (prima della fusione con i lisosomi), mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno- dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). In alcuni distretti la presenza di certe sostanze che un patogeno può utilizzare a scopo nutrizionale può rappresentare un elemento determinante per favorirne la localizzazione in quel distretto che diviene bersaglio preferenziale es. P. mirabilis provoca inf. Urinarie per la frande capacità di produrre ureasi che gli consente di utilizzare l’urea come sorgente di N. es Brucella spp. Causa nei bovini malattia di modesta gravità nell’animale gravido causa aborto per l’invasione degli annessi fetali che sono ricchi di eritritolo, un carboidrato che per le brucelle è molto utile")
68
Capsula Colonizzazione delle mucose Contrasta gli effetti potenzialmente nocivi dell’interazione adesine-recettore fagocita Maschera le adesine, varia la carica di superficie batterica Ostacola la deposizione di C3b e l’ancoraggio di Ab Strategia + comune nei G+ Nei G- mancata produzione di fimbrie Attività antifagocitaria : fibrille (acidi teicoci + prot M) in S.pyognes o capsula in numerosi batteri esempi Pneumococchi, klebsiella emofili 68 68
 in S.pyognes o capsula in numerosi batteri esempi Pneumococchi, klebsiella emofili")
69
Sopravvivenza nella sottomucosa e nei tessuti profondi
Una volta all’interno di fagociti professionali, alcuni batteri patogeni sono in grado di evadere dal fagosoma (prima della fusione con i lisosomi), mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno-dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). Una volta introdotti all’interno di fagociti professionali, alcuni batteri patogeni sono in grado di evadere rapidamente dal fagosoma (prima della fusione con i lisosomi), mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno- dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). In alcuni distretti la presenza di certe sostanze che un patogeno può utilizzare a scopo nutrizionale può rappresentare un elemento determinante per favorirne la localizzazione in quel distretto che diviene bersaglio preferenziale es. P. mirabilis provoca inf. Urinarie per la frande capacità di produrre ureasi che gli consente di utilizzare l’urea come sorgente di N. es Brucella spp. Causa nei bovini malattia di modesta gravità nell’animale gravido causa aborto per l’invasione degli annessi fetali che sono ricchi di eritritolo, un carboidrato che per le brucelle è molto utile. 69 69
, mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno-dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). Una volta introdotti all’interno di fagociti professionali, alcuni batteri patogeni sono in grado di evadere rapidamente dal fagosoma (prima della fusione con i lisosomi), mentre altri sono in grado di moltiplicarsi all’interno del fagosoma alterando la membrana impedendo la fusione con il lisosoma, oppure produzione di catalasi e SOD per contrastare i meccanismi di killing ossigeno- dipendenti, eliminado i derivati dell’ossigeno molecolare provvisti di azione microbicida (perossido di idrogeno, anione superossido etc). In alcuni distretti la presenza di certe sostanze che un patogeno può utilizzare a scopo nutrizionale può rappresentare un elemento determinante per favorirne la localizzazione in quel distretto che diviene bersaglio preferenziale es. P. mirabilis provoca inf. Urinarie per la frande capacità di produrre ureasi che gli consente di utilizzare l’urea come sorgente di N. es Brucella spp. Causa nei bovini malattia di modesta gravità nell’animale gravido causa aborto per l’invasione degli annessi fetali che sono ricchi di eritritolo, un carboidrato che per le brucelle è molto utile")
70
70 70

71
Le shigelle non essendo in grado di accedere all’interno degli enterociti della mucosa del colon, attraverso la membrana apicale di tali cellule , guadagnano l’accesso alla sottomucosa attraverso le cellule M per transcitosi. I pochi batteri che arrivano alla sottomucosa vengono prontamente fagocitati dai macrofagi residenti nella sottomucosa. Le shigelle fagocitate però evadono rapidamente dal fagosoma all’interno del citosol innescando il processo di apoptosi. L’uccisione e la lisi dei macrofagi provoca la liberazione nell’ambiente delle shigelle fagocitate e di notevoli quantità di IL-1 beta che è un interleuchina che stimola il reclutamento i PMN nel sito d’infezione. L’intensa trasmigrazione di leucociti attraverso l’epitelio verso il lume intestinale compromette l’integrità della barriera epiteliale mucosa, permettendo ad un massiccio numero di batteri presenti alla superficie dell’epitelio mucoso, di raggiungere la zona baso-laterale degli enterociti che, a questo punto vengono invasi dai batteri, con la produzione degli estesi danneggiamenti della mucosa e degli estesi infiltrati infiammatori caratteristici della shigellosi. Le shigelle non essendo in grado di accedere all’interno degli enterociti della mucosa del colon, attraverso la membrana apicale di tali cellule, guadagnano l’accesso alla sottomucosa attraverso le cellule M per transcitosi. I pochi batteri che arrivano alla sottomucosa vengono prontamente fagocitati dai macrofagi residenti nella sottomucosa. 71 71

72
Le salmonelle penetrano attraverso la porzione apicale della membrana cellulare all’interno delle cellule dell’ epitelio mucoso del COLON . Questa proprietà è mediata da proteine effetrici che vengono traslocate nel citosl mediante sistema secIII e provocan o riorganizzazione dei filamenti di actina con inglobazione del batterio nel citosol. Con meccanismo analogo il batterio passa anche nelle cellule contigue 72 72
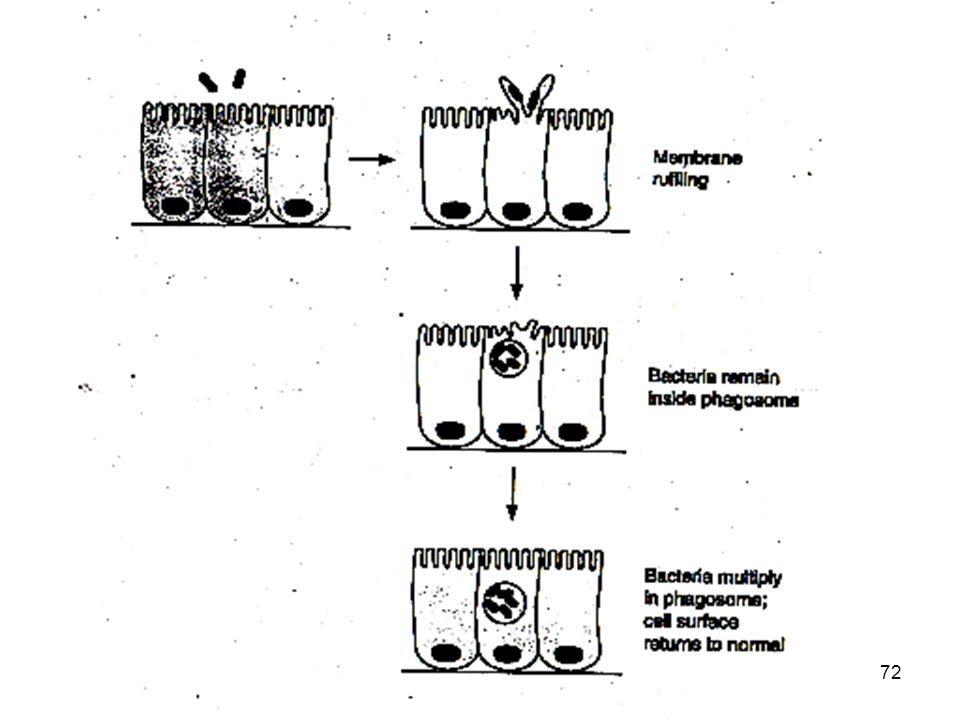
73
Produzione di metaboliti
Sono costituiti da prodotti originati dalla crescita batterica in seguito alla demolizione di composti complessi, molti di questi, come l’acido lattico, alcoli, acetone o aldeidi o comunque composti organici possono esercitare un effetto tossico sulle cellule. 73 73

74
Potere aggressivo dei batteri
Sostanze non tossiche: capsula, coagulasi, catalasi (H2O2) SOD (O2-), collagenasi, ialuronidasi ecc. Sostanze tossiche: Esotossine: esocitate, proteiche Endotossine: legate al batterio LPS Divisione non netta i.e. endotossine proteiche: V. cholerae Le tossine sensu stricto comprendono le esotossine e la endotossina. L’endotossina presenta un diverso gradi di tossigenicità nei diversi gruppi dei G-, ma possiede un unico meccanismo d’azione. Le esotossine no. Le esotossine essendo di natura proteica sono ottimi antigeni, sono generalmente termolabili (no enterotossine stafilococciche) e vengono distrutte dai succhi gastrici ( ne enterotossine stsf. E tossina botulino). Molte esotossine in seguito ad una blanda denaturazione chimica non sono più in grado di interagire con le cellule bersaglio o comunque non sono in grado di esplicare la loro azione tossica, ma mantengono intatta la configurazione antigenica (anatossine o tossoidi) per cui possono essere impiegate per l’allestimento dei vaccini (es tetano e difterite). Le esotossine interagiscono con strutture superficiali cellulari (recettori) e possono interagire con più popolazioni di cellule sensibili (pantrope) o con uno specifico tipo cellulare (tossina esfoliativa, leucocidina su macrofagi e granulociti, tossina tetanica e botulinica: neurotrope) 74 74
 SOD (O2-), collagenasi, ialuronidasi ecc. Sostanze tossiche: Esotossine: esocitate, proteiche. Endotossine: legate al batterio LPS. Divisione non netta i.e. endotossine proteiche: V. cholerae. Le tossine sensu stricto comprendono le esotossine e la endotossina. L’endotossina presenta un diverso gradi di tossigenicità nei diversi gruppi dei G-, ma possiede un unico meccanismo d’azione. Le esotossine no. Le esotossine essendo di natura proteica sono ottimi antigeni, sono generalmente termolabili (no enterotossine stafilococciche) e vengono distrutte dai succhi gastrici ( ne enterotossine stsf. E tossina botulino). Molte esotossine in seguito ad una blanda denaturazione chimica non sono più in grado di interagire con le cellule bersaglio o comunque non sono in grado di esplicare la loro azione tossica, ma mantengono intatta la configurazione antigenica (anatossine o tossoidi) per cui possono essere impiegate per l’allestimento dei vaccini (es tetano e difterite). Le esotossine interagiscono con strutture superficiali cellulari (recettori) e possono interagire con più popolazioni di cellule sensibili (pantrope) o con uno specifico tipo cellulare (tossina esfoliativa, leucocidina su macrofagi e granulociti, tossina tetanica e botulinica: neurotrope)")
75
Differenze tra esotossine ed endotossine
Escrete da cellule viventi e ritrovate ad elevate concentrazioni nei terreni Polipeptidi PM Relativamente instabili: tossicità spesso rapidamente distrutta col calore>60° Altamente antigeniche, stimolano la formazione di alti titoli di antitossina che neutralizza la tossina Convertibile in antigene (anatossina) da formalina, acidi, calore ecc. Altamente tossiche, fatali per animali da laboratorio in mg o meno Non producono febbre nell’ospite 75 75
 da formalina, acidi, calore ecc. Altamente tossiche, fatali per animali da laboratorio in mg o meno. Non producono febbre nell’ospite")
76
Differenze tra esotossine ed endotossine
Parte integrante della parete dei gram-negativi Complesso lipopolisaccaridico (LPS), il lipide A è la porzione responsabile della tossicità Stabile al calore>60° per ore senza perdere tossicità Non stimola la formazione di antitossina La parte polisaccaridica (molto variabile) stimola la produzione di anticorpi (Antigene somatico O) Non è convertibile in anatossina Debolmente tossico per animali anche a dosi di mg Spesso causa febbre 76 76
, il lipide A è la porzione responsabile della tossicità. Stabile al calore>60° per ore senza perdere tossicità. Non stimola la formazione di antitossina. La parte polisaccaridica (molto variabile) stimola la produzione di anticorpi (Antigene somatico O) Non è convertibile in anatossina. Debolmente tossico per animali anche a dosi di mg. Spesso causa febbre")
77
Classificazione delle esotossine
TOSSINE CITOLITICHE Attive sulla membrana delle cellule TOSSINE NEUROTROPE Attive sulle cellule del sistema nervoso TOSSINE ENTEROTOSSICHE Attive sulle cellule della mucosa intestinale TOSSINE PANTROPE Attive su tutte le cellule, inibiscono la sintesi proteica 77 77

78
Tossine emolitiche Attività fosfolipasica che idrolizza la fosforilcolina della membrana Formazione di pori attraverso la membrana Tossine emolitiche (emolisine e citolisine) Formazione di pori: tipicamente prodotte dai G+ es: emolisina alfa di S.aureus, streptolisina-O di S.pyogenes, listeriolisina di Listeria monocytogenes, pneumolisina di S. pneumoniae. Qs tossine presentano gruppi SH essenziali per la loro attività e sono inattivate in presenza di ossigeno. Sono proteine monomeriche che polimerizzano sulla superficie formano degli eptameri tubolari che si inseriscono nella porzione lipidica della memb. formando pori che alterano profondamente gli scambi della cellula con l’ambiente, causandone la morte in genere per apoptosi. Tra i G- le tossine che formano i pori appartengono alla famiglia RTX (repeats in toxins: sequenze ricche di glicina di 9 peptidi ripetute + volte all’estremo C-terminale e che conferiscono la capacità di legare ioni Ca che risultano essenziali all’attività della tossina es. emolisina di E.coli e P. vulgaris, leucocidina di Pasteurella haemolytica. Un’ altra tossina citolitica che funziona come detergente (gruppi idrofilici e idrofobici agli estremi opposti) è tossina delta di S.aureus. Emolisine che funzionano da fosfolipasi (esoenzimi). Servono per la digestione di materiali utilizzabili a scopo alimentare ( es terreni al tuorlo d’uovo) e per doffondere nei tessuti es. tossina alfa di Clostridium perfringens (fosfolipasi specifica per fosforilcolina, vedi figura). Emolisina beta di S. aureus fosfolipasi C specifica per sfingomielina e lisofosfatidilcolina e agisce sui lipidi della membrana degli eritrociti (ben evidente in vitro dopo passaggio caldo- freddo. Citoliisne PV e gamma di S.aureus Fig 7.3 LA placa 78 78
 Formazione di pori: tipicamente prodotte dai G+ es: emolisina alfa di S.aureus, streptolisina-O di S.pyogenes, listeriolisina di Listeria monocytogenes, pneumolisina di S. pneumoniae. Qs tossine presentano gruppi SH essenziali per la loro attività e sono inattivate in presenza di ossigeno. Sono proteine monomeriche che polimerizzano sulla superficie formano degli eptameri tubolari che si inseriscono nella porzione lipidica della memb. formando pori che alterano profondamente gli scambi della cellula con l’ambiente, causandone la morte in genere per apoptosi. Tra i G- le tossine che formano i pori appartengono alla famiglia RTX (repeats in toxins: sequenze ricche di glicina di 9 peptidi ripetute + volte all’estremo C-terminale e che conferiscono la capacità di legare ioni Ca che risultano essenziali all’attività della tossina es. emolisina di E.coli e P. vulgaris, leucocidina di Pasteurella haemolytica. Un’ altra tossina citolitica che funziona come detergente (gruppi idrofilici e idrofobici agli estremi opposti) è tossina delta di S.aureus. Emolisine che funzionano da fosfolipasi (esoenzimi). Servono per la digestione di materiali utilizzabili a scopo alimentare ( es terreni al tuorlo d’uovo) e per doffondere nei tessuti es. tossina alfa di Clostridium perfringens (fosfolipasi specifica per fosforilcolina, vedi figura). Emolisina beta di S. aureus fosfolipasi C specifica per sfingomielina e lisofosfatidilcolina e agisce sui lipidi della membrana degli eritrociti (ben evidente in vitro dopo passaggio caldo- freddo. Citoliisne PV e gamma di S.aureus. Fig 7.3 LA placa")
79
Esotossine che agiscono a livello delle strutture della superficie cellulare
Tossina esfoliativa sindrome cute ustionata Tossine emolitiche (emolisine o citolisine) Dal circolo arriva allo strato granuloso del derma, dopo fissazione allo strato attivazione di serino-proteasi che agisce su desmosomi 79 79
 Dal circolo arriva allo strato. granuloso del derma, dopo fissazione allo strato. attivazione di serino-proteasi. che agisce su desmosomi")
80
TOSSINE ENTEROTOSSICHE
Citotossiche che causano danni evidenti alle cellule Citotoniche danni non evidenti Colerica è il prototipo ( daltons) Subunità A e 5-6 subunità B identiche Le sub. B si legano al recettore sulla membrana e facilitano l’ingresso della A La A induce una ADP-ribosilante NAD-dipendente sul GTP che attiva in modo permanente l’adenilciclasi questo provoca un perturbazione nel passaggio di acqua ed elettroliti (perdita di acqua attraverso il lume intestinale) Con meccanismo simile si ritrova in E. coli, K. pneumoniae, S. typhimurium e S. enteritidis 80 80
 Subunità A e 5-6 subunità B identiche Le sub. B si legano al recettore sulla membrana e facilitano l’ingresso della A. La A induce una ADP-ribosilante NAD-dipendente sul GTP che attiva in. modo permanente l’adenilciclasi. questo provoca un perturbazione nel passaggio di acqua ed elettroliti. (perdita di acqua attraverso il lume intestinale) Con meccanismo simile si ritrova in E. coli, K. pneumoniae, S. typhimurium e S. enteritidis")
81
Il frammento A1, in presenza di NAD provoca la ADP-ribosilazione di una proteina accettrice di GTP che è la proteina attivatrice della adenilato-ciclasi e che viene immobilizzata nello stato attivato provocando a sua volta l’attivazione ininterrotta dell’enzima con produzione di una notevole quantità di AMP-ciclico che interviene nella regolazione degli scambi elettrolitici. La tossina della pertosse lega il gruppo ADP-riboso ad una proteina regolatrice della adenilato-ciclasi. La tossina della pertosse ha come substrato una proteina in grado di accettare GTP con funzione inibitoria che viene resa inattiva. 81 81

82
Tossina pertossica Il frammento A1, in presenza di NAD provoca la ADP-ribosilazione di una proteina accettrice di GTP che è la proteina attivatrice della adenilato-ciclasi e che viene immobilizzata nello stato attivato provocando a sua volta l’attivazione ininterrotta dell’enzima con produzione di una notevole quantità di AMP-ciclico che interviene nella regolazione degli scambi elettrolitici. La tossina della pertosse lega il gruppo ADP-riboso ad una proteina regolatrice della adenilato-ciclasi. La tossina della pertosse ha come substrato una proteina in grado di accettare GTP con funzione inibitoria che viene resa inattiva. La tossina della pertosse è una tossina pantropa che diffonde dalla mucosa tracheo-bronchiale e può interagire con diversi tipi di cellule provocando diverse manifestazioni. Inibizione dell’attività fagocitaria dei macrofagi, leucocitosi, aumento della sensibilità all’istamina, aumentata produzione di insulina.. 82 82

83
amminoacidi sulla catena peptidica nascente.
L’esotossina dissenterica è prodotta dalla Shigella dysenteriae di tipo 1. Neurotossica ed enterotossica. L’inibizione della sintesi proteica avviene con un meccanismo non ancora noto. Si osserva un’inibizione del trasferimento degli amminoacidi sulla catena peptidica nascente. Tossina della pertosse. Proteina ( D) composta da 5 subunità. La produzione di cAMP è regolata da due proteine situate all’interno della membrana cellulare: Gs che ha funzione stimolante e Gi che svolge attività inibente. La tossina colerica ha come bersaglio finale la Gs mantenendola in uno stato attivo permanente, mentre la tossina della pertosse interagisce con Gi impedendo qualsiasi attività inibente. 83 83
 composta da 5 subunità. La produzione di cAMP è regolata da due proteine situate all’interno della. membrana cellulare: Gs che ha funzione stimolante e Gi che svolge. attività inibente. La tossina colerica ha come bersaglio finale la Gs. mantenendola in uno stato attivo permanente, mentre la tossina della pertosse. interagisce con Gi impedendo qualsiasi attività inibente")
84
TOSSINE PANTROPE Tossina difterica (prototipo), catena polipeptidica di D unita da 2 ponti S-S, la molecola è tagliata da proteasi in due frammenti A e B che restano uniti da S-S Il frammento B riconosce il bersaglio cellulare, mediante la sua parte COOH terminale, sulla superficie della cellula. Si lega alla parte idrofoba della membrana e crea un canale attraverso il quale il frammento A penetra all’interno Il glutatione (presente all’interno della cellula) libera il frammento A, enzimaticamente attivo (ADP-ribosilante). Agisce sul NAD staccando ADP-riboso che interagisce con il fattore EF2 bloccando la traslocazione sul ribosoma Il complesso EF2-ADP potrebbe ancora reagire con il ribosoma e il GTP (che fornisce energia), tuttavia è bloccata l’idrolisi del GTP, non si ha quindi energia e la sintesi proteica è interrotta. 84 84
, catena polipeptidica di D unita da 2 ponti S-S, la molecola è tagliata da proteasi in due frammenti A e B che restano uniti da S-S. Il frammento B riconosce il bersaglio cellulare, mediante la sua parte COOH terminale, sulla superficie della cellula. Si lega alla parte idrofoba della membrana e crea un canale attraverso il quale il frammento A penetra all’interno. Il glutatione (presente all’interno della cellula) libera il frammento A, enzimaticamente attivo (ADP-ribosilante). Agisce sul NAD staccando ADP-riboso che interagisce con il fattore EF2 bloccando la traslocazione sul ribosoma. Il complesso EF2-ADP potrebbe ancora reagire con il ribosoma e il GTP (che fornisce energia), tuttavia è bloccata l’idrolisi del GTP, non si ha quindi energia e la sintesi proteica è interrotta")
85
Esotossine che inibiscono la sintesi proteica cellulare
La tossina difterica è codificata dal gene tox di un fago temperato integrato nel cromosoma che viene trascritto in condizioni di stress da carenza di ferro. La tossina di tipo A-B, è sintetizzata come un unico peptide formato da 535 aa, del p.m. complessivo di , il cui estremo carbossi terminale forma il componente B, mentre la porzione amino-terminale forma il componente A. La tossina diffonde nell’organismo e si lega con la porzione B a recettori glicoproteici ampiamente diffusi nelle cellule animali (tossina pantropa) dove viene scissa da proteasi di membrana nei due componenti A e B (21500kd e kd) che rimangono uniti da un ponte disolfuro. La tossina viene introdotta nella cellula per endocitosi mediata dal recettore e dopo l’interruzione del ponte disolfurico ad opera del glutatione (sostanza riducente), il componente A viene traslocato nel citosol attraverso la membrana della vescicola endocitica. Il componente A possiede un’ attività ADP-ribosilante, simile a quella delle tossine colerica, LT e pertossica. Il bersaglio dell’attività ADP-ribosil-trasferasica di A è il fattore di allungamento EF-2 che interviene nella sintesi della catena peptidica a livello della traslocazione sul ribosoma. Il complesso che ne deriva EF-2-ADP-riboso che ne risulta è inattivo e di conseguenza la sintesi proteica viene bloccata, segue morte della cellula. 85 85
 dove viene scissa da proteasi di membrana nei due componenti A e B (21500kd e 40500kd) che rimangono uniti da un ponte disolfuro. La tossina viene introdotta nella cellula per endocitosi mediata dal recettore e dopo l’interruzione del ponte disolfurico ad opera del glutatione (sostanza riducente), il componente A viene traslocato nel citosol attraverso la membrana della vescicola endocitica. Il componente A possiede un’ attività ADP-ribosilante, simile a quella delle tossine colerica, LT e pertossica. Il bersaglio dell’attività ADP-ribosil-trasferasica di A è il fattore di allungamento EF-2 che interviene nella sintesi della catena peptidica a livello della traslocazione sul ribosoma. Il complesso che ne deriva EF-2-ADP-riboso che ne risulta è inattivo e di conseguenza la sintesi proteica viene bloccata, segue morte della cellula")
86
Esotossine citotossiche per azione sul citoscheletro
Sono tutte tossine binarie, formate cioè da due distinti componenti A e B che sono secreti separatamente e si riuniscono solo alla superficie della cellula bersaglio, possiedono tutte un attività catalitica ADP- ribosiltransferasica ed hanno come specifico bersaglio l’actina. C2 di C. botulinum, fattore citotossico necrotizzante prodotto da molti stipiti di E. coli uropatogeni nell’uomo e animali e la tossina dermonecrotica di Bordetella spp. Tossine sono denominate CDT (cytolethal distending toxins) dimostrate in in E.coli, Shigella e Salmonella. induce danni nel DNA che innescano i sistemi di riparo e il blocco del ciclo cellulare Le cellule bersaglio principali sono monociti- macrofagi. Sono tutte tossine binarie, formate cioè da due distinti componenti A e B che sono secreti separatamente e si riuniscono solo alla superficie della cellula bersaglio, possiedono tutte un attività catalitica ADP-ribosiltransferasica ed hanno come specifico bersaglio l’actina. L’esempio più studiato è quello della tossina C2 di C. botulinum. I due componenti C2I e C2II sono secreti dal batterio (soprattutto durante la fase di sporulazione) separatamente ed in rapporti differenti. C2II attivato proteoliticamente si lega alla superficie della cellula esponendo un sito che consente il successivo legame di C2I. Il complesso che ne risulta viene introdotto nella cellula per endocitosi e successivamente traslocata nel citosol. La sub-U C2I possiede un’attività ADP-ribosil-transferasica in grado di catalizzare l’ADP-ribosilazione dell’actina monomerica e dell’actina F non polimerizzata, alterando il citoscheletro e portando a morte la cellula. Altri esempi: fattore citotossico necrotizzante prodotto in due varietà antigene da molti stipiti di E. coli uropatogeni nell’uomo e gli animali o causa di diarree e setticemie nei ruminanti e la tossina dermonecrotica di Bordetella spp. Esiste poi un gruppo di tossine altamente conservate che comportano indirettamente una serie di alterazioni del citoscheletro. Tali tossine sono denominate CDT (cytolethal distending toxins) dimiostrate in in E.coli, Shigella e Salmonella. Sono di tipo AB la porzione A dal citoplasma passa al nucleo dove induce danni nel DNA che innescano i sistemi di riparo e il blocco del ciclo cellulare in G2 con aumento del volume cellulare e connesse modificazioni del citoscheletro, cui può far seguito la morte cellulare . Le cellule bersaglio principali sono monociti- macrofagi. 86 86
 dimostrate in in E.coli, Shigella e Salmonella. induce danni nel DNA che innescano i sistemi di riparo e il blocco del ciclo cellulare. Le cellule bersaglio principali sono monociti- macrofagi. Sono tutte tossine binarie, formate cioè da due distinti componenti A e B che sono secreti separatamente e si riuniscono solo alla superficie della cellula bersaglio, possiedono tutte un attività catalitica ADP-ribosiltransferasica ed hanno come specifico bersaglio l’actina. L’esempio più studiato è quello della tossina C2 di C. botulinum. I due componenti C2I e C2II sono secreti dal batterio (soprattutto durante la fase di sporulazione) separatamente ed in rapporti differenti. C2II attivato proteoliticamente si lega alla superficie della cellula esponendo un sito che consente il successivo legame di C2I. Il complesso che ne risulta viene introdotto nella cellula per endocitosi e successivamente traslocata nel citosol. La sub-U C2I possiede un’attività ADP-ribosil-transferasica in grado di catalizzare l’ADP-ribosilazione dell’actina monomerica e dell’actina F non polimerizzata, alterando il citoscheletro e portando a morte la cellula. Altri esempi: fattore citotossico necrotizzante prodotto in due varietà antigene da molti stipiti di E. coli uropatogeni nell’uomo e gli animali o causa di diarree e setticemie nei ruminanti e la tossina dermonecrotica di Bordetella spp. Esiste poi un gruppo di tossine altamente conservate che comportano indirettamente una serie di alterazioni del citoscheletro. Tali tossine sono denominate CDT (cytolethal distending toxins) dimiostrate in in E.coli, Shigella e Salmonella. Sono di tipo AB la porzione A dal citoplasma passa al nucleo dove induce danni nel DNA che innescano i sistemi di riparo e il blocco del ciclo cellulare in G2 con aumento del volume cellulare e connesse modificazioni del citoscheletro, cui può far seguito la morte cellulare . Le cellule bersaglio principali sono monociti- macrofagi")
87
TOSSINE NEUROTROPE Tetanica e del carbonchio attive sulle cellule del SNC Botulinica attiva sulle cellule del sistema periferico La botulinica blocca la liberazione di acetilcolina nella giunzione neuromuscolare si ha quindi paralisi flacida (meccanismo non noto) La tetanica blocca l’inibitore del riflesso si ha quindi contrazione, diffonde attraverso il midollo spinale e giunge ai centri nervosi, blocca i mediatori (glicina, acido gamma-aminobutirrico, nucleotidi ciclici) si fissa sui recettori gangliosidici della membrana dei motoneuroni a livello presinaptico (paralisi spastica) 87 87
 La tetanica blocca l’inibitore del riflesso si ha quindi contrazione, diffonde attraverso il midollo spinale e giunge ai centri nervosi, blocca i mediatori (glicina, acido gamma-aminobutirrico, nucleotidi ciclici) si fissa sui recettori gangliosidici della membrana dei motoneuroni a livello presinaptico (paralisi spastica)")
88
Le tossine neurotrope ( tetanica e botulinica )interferiscono con la trasmissione degli impulsi nervosi rispettivamente a livello del SNC e SNP. Tetano: le spore di C. tetani vengono introdotte nei tessuti in seguito ad un trauma, se si realizzano le condizioni di anaerobiosi le spore germinano e la forma vegetativa produce la tossina che per via ematica risale centripetamente lungo i nervi periferici e raggiunge il SNC dove blocca gli impulsi inibitori della contrazione muscolare riflessa provocando una serie di spasmi generalizzati (paralisi spastica). La tossina (di cui si conosce una sola configurazione antigene ed è codificata da un plasmide) è sintetizzata come un’unica catena peptidica di circa 150 Kd che si libera in seguito alla lisi del batterio ed è tagliata da una proteasi in due peptidi di 100 H (heavy) e L 50 kd che rimangono uniti da un ponte disolfurico. H si lega alle cellule nervose e consente la penetrazione di L che è una zincopeptidasi che ha il suo bersaglio nella sinaptobrevina (proteina associata alla membrana delle vescicole) presente nell’apparato preposto all’esocitosi del GABA (acido gamma-amino- butirrico) GABA 88 88
. La tossina (di cui si conosce una sola configurazione antigene ed è codificata da un plasmide) è sintetizzata come un’unica catena peptidica di circa 150 Kd che si libera in seguito alla lisi del batterio ed è tagliata da una proteasi in due peptidi di 100 H (heavy) e L 50 kd che rimangono uniti da un ponte disolfurico. H si lega alle cellule nervose e consente la penetrazione di L che è una zincopeptidasi che ha il suo bersaglio nella sinaptobrevina (proteina associata alla membrana delle vescicole) presente nell’apparato preposto all’esocitosi del GABA (acido gamma-amino- butirrico) GABA")
89
C. botulinum è largamente diffuso nell’ambiente, può contaminare alimenti in presenza di parziale o totale anaerobiosi (es. conserve sott’olio..). La tossina è distrutta a 80°C per 30min, non è quindi presente nei cibi adeguatamente sterilizzati, ma è resistente ai succhi gastrici. Introdotta con gli alimenti si assorbe nell’intestino e diffonde con bersaglio il SNP impedendo il rilascio di acetilcolina a livello della sinapsi colinergica a livello delle giunzioni neuromuscolari, con conseguente paralisi flaccida. Della tossina botulinica si conoscono almeno 7 diversi tipi antigenici A-G prodotti da differenti stipiti di C. botulinum e codificati da geni di volta in volta a collocazione cromosomica (A), plasmidica (G) o in un fago temperato integrato (C-D). Anche la t. botulinica viene sintetizzata come un unico peptide che viene poi scisso in 2 frammenti H e L tenuti insieme da un ponte disolfurico. Anche in questo caso H si lega al recettore ed L è una zincopeptidasi che interagisce con proteine che intervengono nel rilascio dell’acetilcolina: sinaptobrevina, proteina SNAP 25 e sintaxina. L’inibizione dell’esocitosi di acetilcolina blocca la trasmisssione dell’impulso nervoso alla muscolatura con conseguente paralisi flaccida. 89 89
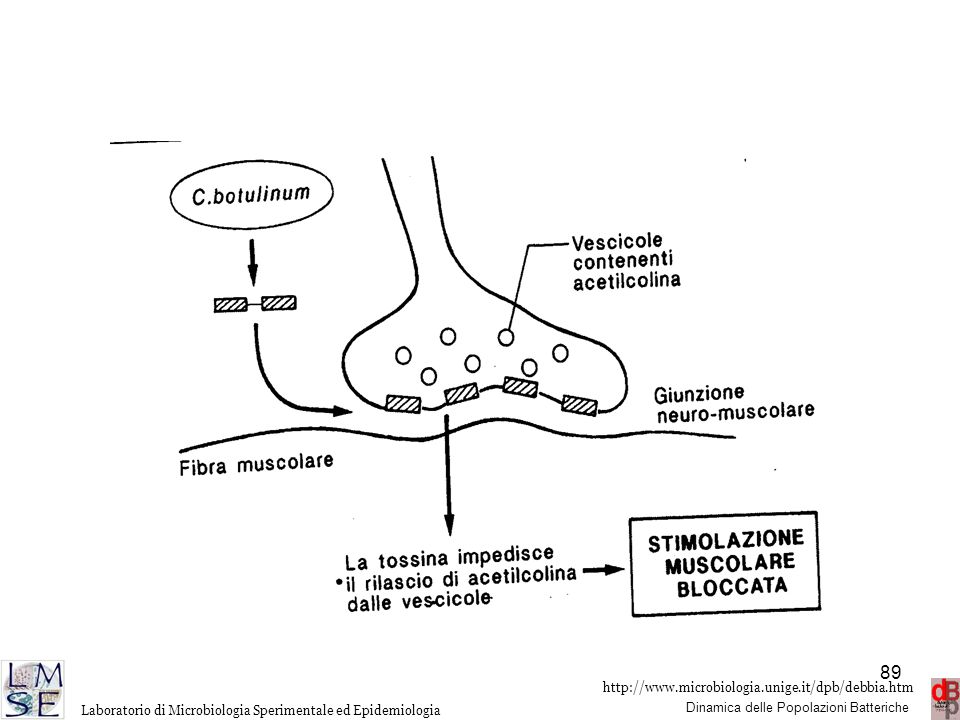
90
Diagramma semplificato del controllo nervoso dei muscoli che consentono la flessione e l'estensione dell'avambraccio, nel soggetto normale (a sinistra), nell'intossicazione botulinica (al centro) e nell'intossicazione tetanica (a destra). Nel soggetto normale, l'impulso proveniente dal cervello (cerchio piccolo) eccita il neurone motore(1) nel midollo spinale (cerchio grande), dal quale l'impulso viene trasmesso al bicipite che si contrae provocando la fles sione dell'avambraccio. La contrazione del bicipite provoca lo stiramento del tricipite dai cui recettori (2) parte un impulso destinato ad eccitare i motoneuroni del tricipite (3) i quali provocherebbero la contrazione del tricipite se non fossero sotto il controllo di un neurone inibitore (4). Nell'intossicazione botulinica si ha il blocco della liberazione di acetilcolina a livello della giunzione neuromuscolare (5). In questo modo l'impulso nervoso non raggiunge il muscolo e si ha una paralisi flaccida. Nell'intossicazione tetanica è bloccato l'impulso nervoso inibitore (6) e si ha contemporaneamente la contrazione del bicipite e del tricipite con la conseguente paralisi spastica. 90 90

91
Tossina botulinica Sulla base del valore di LD50 di circa 1 ng/kg [3], poche centinaia di grammi di questa tossina potrebbero teoricamente uccidere ogni essere umano presente sulla Terra (a scopo comparativo, la stricnina richiederebbe 400 tonnellate per uccidere ogni essere umano). Generalmente le dosi letali orali risultano comprese fra 0,5 e 5 mg/kg di alimento ingerito.
. Generalmente le dosi letali orali risultano comprese fra 0,5 e 5 mg/kg di alimento ingerito.")
92
92 92

93
Endotossina Ripetizione di unità diverse nelle diverse specie
Si intende per endotossina il LPS che costitutisce lo strato periferico della membrana esterna dei G-. Il lipide A , formato da un disaccaride (glucosamina) fosforilato, ed esterificato con diversi acidi grassi saturi rappresenta la porzione tossica del LPS. Al lipide A è legata una complessa struttura polisaccaridica formata da un Core e dall’antigene O. Il core è caratterizzato dalla presenza costante di alcuni zuccheri particolari come l’acido cheto- deossioctonico (KDO) e un eptoso rappresentato in genere da L-glicero-D- mannoeptoso. Questa porzione è praticamente identica in tutti i batteri G-. Al core è ancorata la porzione polisaccaridica specifica, formata da una lunga catena talora ramificata, formata dalla ripetizione di sub-u tri-tetra-penta- saccaridiche formate da zuccheri diversi nelle diverse specie batteriche (antigene O). La porzione polisaccaridica che può essere persa (antigene O manca nelle varianti rugose) contribuisce probabilmente alla tossicità del lipide A, influenzandone la idrosolubilità e condizionandone la struttura supramolecolare sopprattutto la parte contenente il KDO. Data la sua natura chimica, la porzione glicolipidica dell’LPS, ovvero la porzione dotata di attività tossica è relativamente termostabile e poco dotata di potere immunogeno (negli ultimi anni numerosi sforzi sono stati dedicati ai tentativi di produzione di anticorpi anti-lipideA da utilizzare nelle forme più gravi da intossicazione endotossica con risultati di scarso rilievo) e non è detossificabile in preparazioni omologabili alle anatossine o tossoidi che si possono preparare da numerose esotossine. Ripetizione di unità diverse nelle diverse specie Contribuisce alla tossicità del Lipide A influenzandone l’idrosolubilità e la struttura Glucosamina fosforilata ed esterificata con acidi grassi saturi 93 93
 fosforilato, ed esterificato con diversi acidi grassi saturi rappresenta la porzione tossica del LPS. Al lipide A è legata una complessa struttura polisaccaridica formata da un Core e dall’antigene O. Il core è caratterizzato dalla presenza costante di alcuni zuccheri particolari come l’acido cheto- deossioctonico (KDO) e un eptoso rappresentato in genere da L-glicero-D- mannoeptoso. Questa porzione è praticamente identica in tutti i batteri G-. Al core è ancorata la porzione polisaccaridica specifica, formata da una lunga catena talora ramificata, formata dalla ripetizione di sub-u tri-tetra-penta- saccaridiche formate da zuccheri diversi nelle diverse specie batteriche (antigene O). La porzione polisaccaridica che può essere persa (antigene O manca nelle varianti rugose) contribuisce probabilmente alla tossicità del lipide A, influenzandone la idrosolubilità e condizionandone la struttura supramolecolare sopprattutto la parte contenente il KDO. Data la sua natura chimica, la porzione glicolipidica dell’LPS, ovvero la porzione dotata di attività tossica è relativamente termostabile e poco dotata di potere immunogeno (negli ultimi anni numerosi sforzi sono stati dedicati ai tentativi di produzione di anticorpi anti-lipideA da utilizzare nelle forme più gravi da intossicazione endotossica con risultati di scarso rilievo) e non è detossificabile in preparazioni omologabili alle anatossine o tossoidi che si possono preparare da numerose esotossine. Ripetizione di unità diverse nelle diverse specie. Contribuisce alla tossicità del Lipide A influenzandone l’idrosolubilità e la struttura. Glucosamina fosforilata ed esterificata con acidi grassi saturi")
94
Endotossina LPS si lega a proteina di memb. CD14 (recettore) dei macrofagi Le LBP prodotte dal fegato e presenti in circolo si combinano con LPS e catalizzano il suo trasferimento sul macrofago e il legame con CD14 Recettori Tlr presenti sui macrofagi in contatto con LPS legato a CD14, trasducono il segnale di membrana rilascio citochine Sin dagli inizi degli anni Novanta , sappiamo che l’interazione LPS-macrofagi è mediata dall’unione dell’LPS alla proteina macrofagica di membrana CD14 che è il recettore. Sappiamo poi che alcune proteine LBP (LPS-binding protein) prodotte nel fegato e presenti in circolo, sono in grado di combinarsi specificamente all’LPS e di veicolarlo sui macrofagi, catalizzandone il trasferimento sul CD14. Per molto tempo però non si è riusciti a comprendere quale fosse lo strumento di trasduzione del segnale di membrana in grado di attivare i macrofagi dato che la proteina CD14 è priva di una qualsiasi porzione transmembranaria in grado di raggiungere il citosol e di connettersi quindi a strutture (proteine-G simile) coinvolte nella trasduzione del signalling. Solo sul finire degli anni 1990 si è scoperto che sui macrofagi umani esistono alcuni recettori simili a cosiddetti Toll presenti in Drosophila. Questi recettori costituiscono una famiglia conservata detta Tlr Toll-like receptor. Sono questi recettori che, venendo in contatto con LPS ancorato al CD14, trasducono i segnali di attivazione macrofagica cui segue la liberazione di varie citochine. 94 94
 prodotte nel fegato e presenti in circolo, sono in grado di combinarsi specificamente all’LPS e di veicolarlo sui macrofagi, catalizzandone il trasferimento sul CD14. Per molto tempo però non si è riusciti a comprendere quale fosse lo strumento di trasduzione del segnale di membrana in grado di attivare i macrofagi dato che la proteina CD14 è priva di una qualsiasi porzione transmembranaria in grado di raggiungere il citosol e di connettersi quindi a strutture (proteine-G simile) coinvolte nella trasduzione del signalling. Solo sul finire degli anni 1990 si è scoperto che sui macrofagi umani esistono alcuni recettori simili a cosiddetti Toll presenti in Drosophila. Questi recettori costituiscono una famiglia conservata detta Tlr Toll-like receptor. Sono questi recettori che, venendo in contatto con LPS ancorato al CD14, trasducono i segnali di attivazione macrofagica cui segue la liberazione di varie citochine")
95
Endotossina In passato si riteneva che LPS agisse direttamente sulle membrane biologiche Oggi si sa che: stimola i macrofagi al rilascio di sostanze che sono le reali effettrici dell’attività tossica Attiva la cascata del complemento Attiva la cascata della coagulazione 95 95

96
I principali mediatori cellulari rilasciati dai macrofagi attivati in seguito al legame con l’LPS, sono rappresentati da TNF e IL-1. TNF è una citochina pleotropa con una serie di effetti su nemrosi elementi cellulari. Insieme a IL-1, responsabile primaria dell’azione pirogena, attiva la via metabolica dell’acido arachidonico con la produzione di prostaglandine che agiscono sugli endoteli (causando aumento della permeabilità vascolare, vasodilatazione, ipotensione e shock emodinamico) ha azione mitogena sui linfociti con incremento della produzione di interferone gamma, induce l’aumento del catabolismo proteico, rallenta il metabolismo del Fe, agisce sul fegato con conseguente ipoglicemia e stimola il fegato alla produzione di beta2-microglobulina a altre proteine cosiddette della fase acuta, svolgendo azione proinfiammatoria. Alle azioni indotte dal rilascio di TNF e IL-1 da parte dei macrofagi, vanno aggiunti gli effetti dell’attivazione del complemento per la via alternativa che, in presenza di massicce dosi di LPS, piò portare alla formazione di elevate quantità di C3a C5a che svolgono azione proinfiammatoria e sono in grado di danneggiare gli endoteli vascolari, favorendo la comparsa dello shock emodinamico, nonché gli effetti dell’attivazione dell’azione procoagulante dei linfomonociti, che facilita l’aggregazione piastrinica e l’attivazione della cascata coagulativa a partire dal fattore di Hageman, con conseguenze che possono giungere alla comparsa della CID. 96 96

97
Effetti dell'endotossina
- febbre (attivazione di IL-l) - neutropenia: adesivita' delle cellule ai vasi, granulocitosi - ipoglicemia: intensa attività metabolica cellulare glicolisi - ipotensione: vasodilatazione, permeabilità, stagnazione di liquidi - anormale perfusione di organi essenziali - attivazione del complemento - disseminata coagulazione intravascolare - reazioni di Schwartzman (locale e generalizzata) 97 97
 - neutropenia: adesivita delle cellule ai vasi, granulocitosi. - ipoglicemia: intensa attività metabolica cellulare glicolisi. - ipotensione: vasodilatazione, permeabilità, stagnazione di liquidi. - anormale perfusione di organi essenziali. - attivazione del complemento. - disseminata coagulazione intravascolare. - reazioni di Schwartzman (locale e generalizzata)")
98
Superantigene Alcune tossine particolari liberate da alcuni patogeni attivano le cellule T senza richiesta di antigene, si legano al recettore di queste cellule sia a quello del complesso maggiore di istocompatibilità che può scatenare una risposta immune nei confronti delle stesse cellule, quindi molto rischioso. Esempi di queste tossine sono quelle dello shock tossico di S.aureus e quelle eritrogeniche di S.pyogenes. 98 98

99
superantigeni Enterotossine stafilococciche
TSST (Tossina dello Shock Tossico) T. pirogeniche streptococciche Devono parte della loro tossicità a: Specifica attività enzimatica (es. attivazione interleuchina Sono fortemente pirogene Interagiscono con i LT, attivazione e moltiplicazione in % amplificata (2-20% vs % normale Ag) 99 99
 T. pirogeniche streptococciche. Devono parte della loro tossicità a: Specifica attività enzimatica (es. attivazione interleuchina. Sono fortemente pirogene. Interagiscono con i LT, attivazione e moltiplicazione in % amplificata (2-20% vs % normale Ag)")
100
L’antigene convenzionale, internalizzato ed elaborato dalla cellula viene esposto nel contesto del MHC II al riconoscimento del TCR con specifica capacità combinatoria Il superantigene lega direttamente, senza precedente internaliz. ed elaboraz., le proteine del MHC II e il TCR 100 100

101
Infiammazione Si ha in risposta di infezione batterica, ma in particolare per i virus, si ha un eccesso di risposta immune in proporzione al processo infettivo. Chlamydia, Borrelia e sifilide, ma anche streptococchi (febbre reumatica) 101 101
")
102
102 102
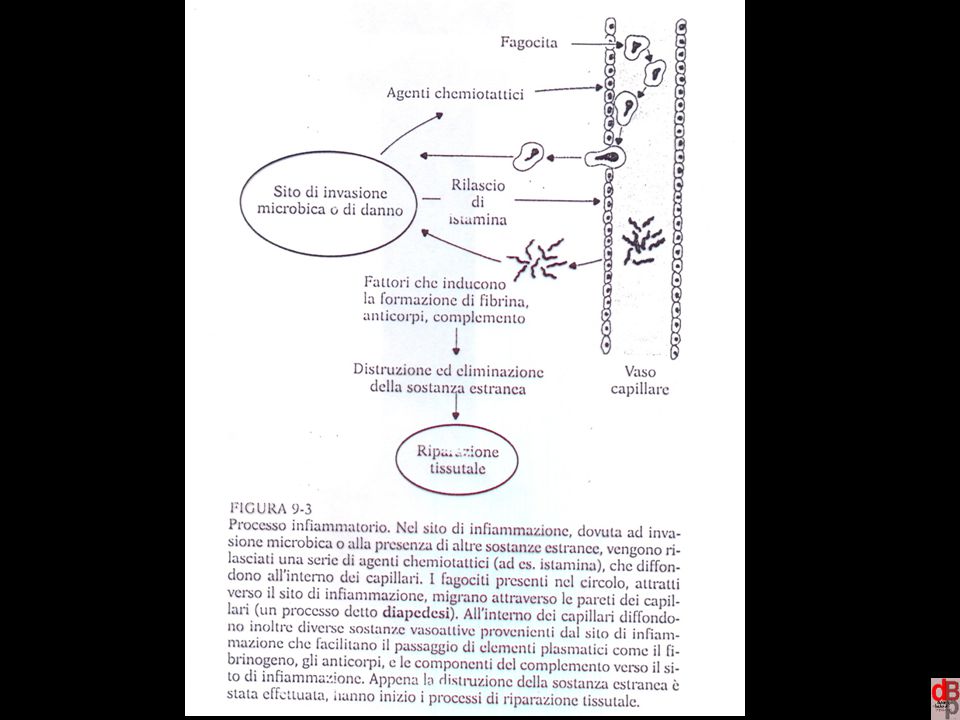
103
103 103
104
IgA Le IgA o immunoglobuline secretorie si trovano in tutti gli epiteli in contatto con l’esterno. Si formano, attraverso la cattura, da parte delle cellule M (microfold) che sono localizzate nei pressi delle placche del Peyer, di corpi estranei, virus, batteri, tossine, ecc. Queste cellule non hanno attività fagocitaria, presentano quindi l’antigene ai macrofagi sottostanti che lo digeriscono e lo presentano alle cellule T che elaborano le IgA specifiche, in questo modo l’organismo ha immunoglobuline per contrastare l’invasione dei microorganismi più frequentemente presenti. 104
 che sono localizzate nei pressi delle placche del Peyer, di corpi estranei, virus, batteri, tossine, ecc. Queste cellule non hanno attività fagocitaria, presentano quindi l’antigene ai macrofagi sottostanti che lo digeriscono e lo presentano alle cellule T che elaborano le IgA specifiche, in questo modo l’organismo ha immunoglobuline per contrastare l’invasione dei microorganismi più frequentemente presenti")
105
105 105
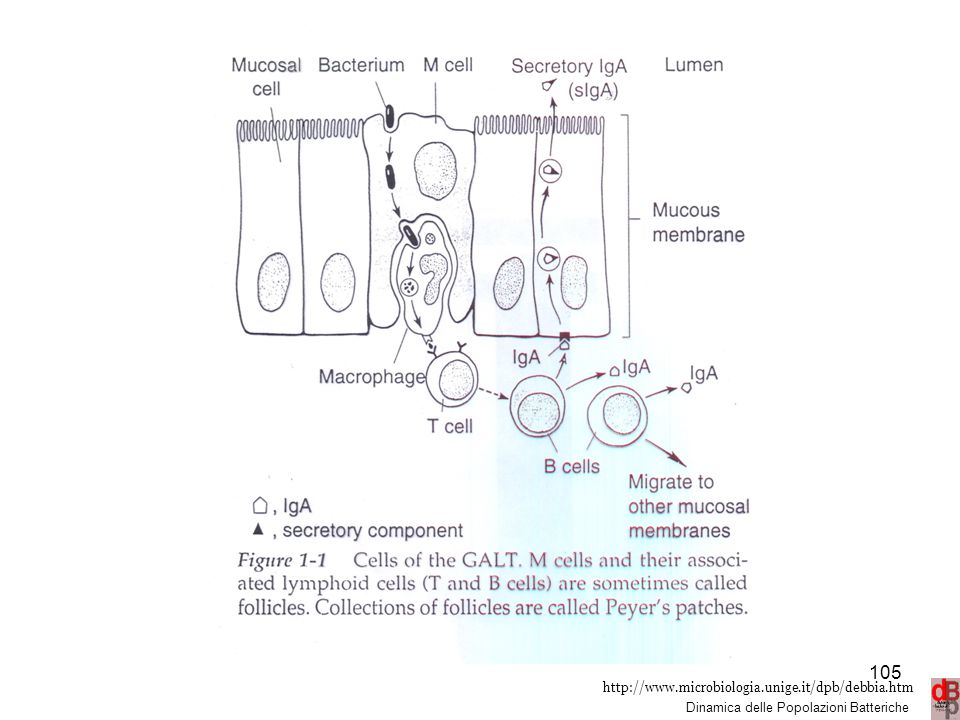
106
IgA La mucosa intestinale secerne Ig, in particolare IgA. Si calcola che ne siano prodotte dai 40 ai 60 mg al giorno per ogni kg di peso corporeo. Nelle placche del Peyer vengono prodotte anche IgM. Le IgA sono considerate l’elemento primario per la risposta immunitaria verso gli antigeni microbici. Le IgG fanno parte della risposta immunitaria innata della mucosa. La mucosa intestinale secerne Ig, in particolare IgA. Si calcola che ne siano prodotte dai 40 ai 60 mg al giorno per ogni kg di peso corporeo. Nelle placche del Peyer vengono prodotte anche IgM. Le IgA sono considerate l’elemento primario per la risposta immunitaria verso gli antigeni microbici. Le IgG fanno parte della risposta immunitaria innata della mucosa. Le IgA secretorie proteggono l’epitelio intestinale contro la colonizzazione e/o l’invasione che si attua attraverso il legame degli antigeni di patogeni o commensali, le IgA ricoprono il germe come uno scudo idrofilico che è repellente per l’epitelio intestinale, quindi è tenuto lontano dalle cellule dell’intestino. 106

107
INTERAZIONI TRA BATTERI Infezioni polimicrobiche
Possono essere più gravi di quelle causate da un singolo patogeno quando le interazioni tra le singole specie trasformano un’infezione mista in una INFEZIONE SINERGICA 107 107

108
INTERAZIONI TRA BATTERI Sinergismo batterico
E’ un fenomeno comune e importante nella produzione di infezioni miste di aerobi ed anaerobi specie in tessuti a lento ricambio o drenaggio o dove l’eliminazione dei patogeni è rallentato 108 108

109
INTERAZIONI TRA BATTERI Sinergismo batterico
E’ un fenomeno ben documentato nelle infezioni sperimentali intraddominali tessuti molli setticemie dentali ginecologiche 109 109

110
Singola specie: innocua Coltura mista: palese infezione
INTERAZIONI TRA BATTERI Sinergismo batterico E’ un fenomeno ben documentato nelle infezioni sperimentali Da peritoniti sono stati isolati i vari microorganismi in coltura pura: Rinoculati in animali Singola specie: innocua Coltura mista: palese infezione 110 110

111
INTERAZIONI TRA BATTERI Sinergismo batterico (ipotesi)
Favorevoli modifiche dell’ambiente Provvedere a nutrienti essenziali Provvedere a fattori di virulenza Modulare la risposta immunitaria Protezione reciproca nei confronti degli agenti antibatterici 111 111

112
INTERAZIONI TRA BATTERI Sinergismo batterico Favorevoli modifiche all’ambiente
Il consumo di O2 da parte dei batteri aerobi nella popolazione mista produce condizioni favorevoli alla proliferazione degli anaerobi, i quali a loro volta proteggono i batteri aerobi dalla fagocitosi Il basso pH dovuto al metabolismo di molti aerobi e acidogeni crea situazioni favorevoli per lo sviluppo di anaerobi e acidofili 112 112

113
INTERAZIONI TRA BATTERI Sinergismo batterico Favorevoli modifiche all’ambiente
Molte specie aerobie che producono capsula (E.coli, K.pneumoniae, S.pneumoniae) raramente producono ascessi se agiscono come unico patogeno mentre il Bacteroides fragilis (anaerobio) produce una capsula che è in grado di promuovere la formazione di ascessi 113 113
 raramente producono ascessi se. agiscono come unico patogeno. mentre il Bacteroides fragilis (anaerobio) produce una capsula che è in grado di. promuovere la formazione di ascessi")
114
INTERAZIONI TRA BATTERI Sinergismo batterico Favorevoli modifiche all’ambiente
ASCESSO: UN AMBIENTE PROTETTO La formazione sinergistica di un ascesso provvede: una barriera alla diffusione degli agenti antibatterici una barriera ai fattori dell’immunità dell’ospite allo sviluppo di più specie batteriche che: >elaborano enzimi inattivanti gli antibiotici (β-lattamasi, cinasi anti aminoglicosidi ecc.) >causano strette condizioni di anaerobiosi che compromettono l’attività di di aminoglicosidi e chinoloni così come la fagocitosi 114 114
 >causano strette condizioni di anaerobiosi che compromettono l’attività di di aminoglicosidi e chinoloni così come la fagocitosi")
115
INTERAZIONI TRA BATTERI Sinergismo batterico Provvedere a nutrienti essenziali (commensalismo)
Veillonelle e difteroidi sintetizzano vit. K (naftaquinone) che è richiesto da B.melaninogenicus per crescere e per esprimere la virulenza S.mutans produce PAB utilizzato da S.sanguis I treponemi fanno uso di acidi grassi (butirrico e succinico) liberato da anaerobi (fusobatteri, batteroidi) Le Veillonelle metabolizzano ac. lattico liberato dalla popolazione microbica normale (PMN) Molti batteri della cavità orale usano ac. lattico liberato dagli altri commensali 115 115
 che è richiesto da B.melaninogenicus per crescere e per esprimere la virulenza. S.mutans produce PAB utilizzato da S.sanguis. I treponemi fanno uso di acidi grassi (butirrico e succinico) liberato da anaerobi (fusobatteri, batteroidi) Le Veillonelle metabolizzano ac. lattico liberato dalla popolazione microbica normale (PMN) Molti batteri della cavità orale usano ac. lattico liberato dagli altri commensali")
116
Fusobacterium necrophorum elabora una leucocidina che protegge
INTERAZIONI TRA BATTERI Sinergismo batterico Provvedere a fattori di virulenza Fusobacterium necrophorum elabora una leucocidina che protegge Corynebacterium pyogenes dall’attività battericida dei leucociti. (C.pyogenes produce un fattore di crescita per F.necrophorum) S.aureus produce ialuronidasi che esalta l’attività necrotica degli streptococchi anaerobi e conduce alla distruzione dei tessuti 116 116
 S.aureus produce ialuronidasi che esalta. l’attività necrotica degli streptococchi. anaerobi e conduce alla distruzione dei. tessuti")
117
INTERAZIONI TRA BATTERI Sinergismo batterico Modulazione della risposta immunitaria
Nelle infezioni miste gli anaerobi inibiscono la fagocitosi degli aerobi mediante disturbi o indebolimenti delle attività di: opsonizzazione chemiotassi dei neutrofili produzione di ossigeno batteriocidia Attraverso il meccanismo di: diminuzione delle opsonine del siero produzione di acidi grassi volatili (butirrico e succinico) tossici per la fagocitosi produzione di leucocidine 117 117
 tossici per la fagocitosi. produzione di leucocidine")
118
B.fragilis e altre specie anaerobie producono β- lattamasi
INTERAZIONI TRA BATTERI Sinergismo batterico Protezione reciproca nei confronti degli agenti antimicrobici B.fragilis e altre specie anaerobie producono β- lattamasi Nelle infezioni miste questi enzimi proteggono le specie sensibili ai β-lattamici inattivando questi antibiotici (patogenicità indiretta) B.fragilis e vari Clostridi possono aminofenilare il cloramfenicolo rendendolo inefficace sulla restante popolazione batterica mista 118 118
 B.fragilis e vari Clostridi possono aminofenilare il cloramfenicolo rendendolo inefficace sulla restante popolazione batterica mista")
119
INTERAZIONI TRA BATTERI Sinergismo batterico Strategie terapeutiche
L’eliminazione selettiva di un componente della popolazione mista fa fallire il sinergismo batterico Farmaci a spettro ristretto Eliminazione di tutti i patogeni in causa Antibiotici ad ampio spettro usualmente β-lattamici in caso di allergie: macrolidi o altri agenti 119 119

120
Altre strategie batteriche per sfuggire alle difese dell’ospite
Capsula: per mascherare le cellule opsonizzate ostacola la fagocitosi (S.pneumoniae, K.pneumoniae, H.influenzae) Coagulasi: il germe si annida nel coagulo per proteggersi dalle difese in generale S.aureus Fibrinasi, jaluronidasi, collagenasi: distruggere l’integrità dei tessuti con penetrazione all’interno (molti opportunisti) Compromettere la funzionalità dei fagociti e delle cellule ciliate: Bordetelle Produzione di enzimi citotossici: S.aureus, S.pyogenes, Pseudomonas spp. A livello di fagocitosi: blocco dei processi ossidativi (Salmonella, Legionella) impedire la fusione tra fagosoma e lisozima (Gonococchi, Micobatteri) 120 120
 Coagulasi: il germe si annida nel coagulo per proteggersi dalle difese in generale S.aureus. Fibrinasi, jaluronidasi, collagenasi: distruggere l’integrità dei tessuti con penetrazione all’interno (molti opportunisti) Compromettere la funzionalità dei fagociti e delle cellule ciliate: Bordetelle. Produzione di enzimi citotossici: S.aureus, S.pyogenes, Pseudomonas spp. A livello di fagocitosi: blocco dei processi ossidativi (Salmonella, Legionella) impedire la fusione tra fagosoma e lisozima (Gonococchi, Micobatteri)")
121
Altre strategie batteriche per sfuggire alle difese dell’ospite
Cattura del ferro (siderofori) in competizione con lattoferrina Sfuggire alla risposta immunitaria: liberare la capsula (pneumococchi) Variazioni di antigenicità (Borrelia e gonococchi) E.coli KI nella meningite infantile Cattura di materiale dell’ospite e distribuirlo sulla superficie esterna per confondere la risposta immunitaria (Treponema pallidum) Liberare la proteina A che si lega alla porzione Fc delle Ig che provoca l’inversione dell’anticorpo (S.aureus) 121 121
 in competizione con lattoferrina. Sfuggire alla risposta immunitaria: liberare la capsula (pneumococchi) Variazioni di antigenicità (Borrelia e gonococchi) E.coli KI nella meningite infantile. Cattura di materiale dell’ospite e distribuirlo sulla superficie esterna per confondere la risposta immunitaria (Treponema pallidum) Liberare la proteina A che si lega alla porzione Fc delle Ig che provoca l’inversione dell’anticorpo (S.aureus)")
123
Altre strategie batteriche per sfuggire alle difese dell’ospite
Fase aerobia (aerobi, >>>ATP) Fase anaerobia (aerobi, <<<ATP) Fase planctonica Fase sessile Ridotta tossicità di LPS P.aeruginosa, Aggregatibacter actinomycetemcomitans Morte cellulare programmata (PCD) 123 123
 Fase anaerobia (aerobi, <<<ATP) Fase planctonica. Fase sessile. Ridotta tossicità di LPS P.aeruginosa, Aggregatibacter actinomycetemcomitans. Morte cellulare programmata (PCD)")
124
Altre strategie batteriche per sfuggire alle difese dell’ospite
Biofilm: Ig, macrofagi, antibiotici, replicazione lenta Fase intracellulare, per sfuggire ai macrofagi, Ig e a molti antibiotici, tasso di crescita Induzione di forme filamentose Quorum sensing, attivazione di alcune funzioni cellulari mediate da ferormoni (in funzione del numero) Attivazione di uno stato vitale ma non coltivabile 124 124
 Attivazione di uno stato vitale ma non coltivabile")
125
S. pneumoniae S. pneumoniae is known for its ability to enter into a state of genetic competence under conditions of high cell density in response to a secreted signaling peptide (QS). Only a fraction of the S. pneumoniae cells become competent in response to the peptide autoinducer and elaborate a bacteriocin that causes the lysis of noncompetent cells. The lysed cells release not only transforming DNA and nutrients but also pneumolysin and other factors important for virulence. S. pneumoniae sacrifices some its relatives for this purpose which facilitates invasion of its host. S. pneumoniae is known for its ability to enter into a state of genetic competence under conditions of high cell population density in response to a secreted signaling peptide. Only a fraction of the S. pneumoniae cells become competent in response to the peptide autoinducer. Those that do so elaborate a bacteriocin that causes the lysis of noncompetent cells in the population. The lysed cells release not only transforming DNA and nutrients but also pneumolysin and other factors important for virulence. Thus, rather than relying on self for secretion of virulence factors, S. pneumoniae sacrifices some its relatives for this purpose which facilitates invasion of its host.
. Only a fraction of the S. pneumoniae cells become competent in response to the peptide autoinducer and elaborate a bacteriocin that causes the lysis of noncompetent cells. The lysed cells release not only transforming DNA and nutrients but also pneumolysin and other factors important for virulence. S. pneumoniae sacrifices some its relatives for this purpose which facilitates invasion of its host. S. pneumoniae is known for its ability to enter into a state of genetic competence under conditions of high cell population density in response to a secreted signaling peptide. Only a fraction of the S. pneumoniae cells become competent in response to the peptide autoinducer. Those that do so elaborate a bacteriocin that causes the lysis of noncompetent cells in the population. The lysed cells release not only transforming DNA and nutrients but also pneumolysin and other factors important for virulence. Thus, rather than relying on self for secretion of virulence factors, S. pneumoniae sacrifices some its relatives for this purpose which facilitates invasion of its host.")
126
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
La vita dei microorganismi è prevalentemente sessile e solo una frazione minima (<0,1 %) mantiene uno stato planctonico 126 126
 mantiene. uno stato planctonico")
127
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
La semplice adesione ad una superficie, attiva nel microorganismo l’espressione di molti geni che codificano sia per tutti i prodotti deputati alla vita sessile sia quelli che sono coinvolti nei meccanismi di patogenicità e virulenza 127 127

128
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
Una volta che i batteri sono adesi, essi si organizzano in microcolonie. Questo fenomeno implica la presenza di un biofilm, ove i microorganismi sono immersi, costituito da polisaccaridi e prodotti metabolici che, come già detto, mediano adesività tra i microorganismi e gli epiteli e tra i batteri stessi. 128 128

129
Biofilm : una resistenza fenotipica
Dunne WM Jr . Clin Microbiol Rev 2002;15: 129 129

130
Biofilm : una strategia batterica
Fasi del biofilm organizzazione rilascio adesione Salvatorelli G. et al.Biofilms: stato dell’arte l’INTERNISTA, 10 (4), 2002 Dunne WM Jr . Clin Microbiol Rev 2002;15: 130 130
, Dunne WM Jr . Clin Microbiol Rev 2002;15:")
131
Biofilm : i riflessi negativi !..
Protezione vs anticorpi Ridotta attività PMN nessuna fagocitosi Rilascio di enzimi inattivanti gli ATB I biofilm possono organizzarsi sulla superficie di differenti mucose o su materiali(protesi,valvole cardiache) iniziando una lenta fase moltiplicativa , dovuta all’acidità e alla bassa tensione di O2 presente nell’ambiente Solo gli elementi planctonici sono distrutti : normali processi di difesa antibiotici Lewis K et al : Antimicrob Agents Chemother 2001;45: 131 131
 iniziando una lenta fase moltiplicativa , dovuta all’acidità e alla bassa tensione di O2 presente nell’ambiente. Solo gli elementi planctonici sono distrutti : normali processi di difesa. antibiotici. Lewis K et al : Antimicrob Agents Chemother 2001;45:")
132
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
Il biofilm rappresenta un nido di microorganismi anche quando tutte le forme planctoniche sono state uccise. Questo spiega perché i cateteri colonizzati sono sorgenti di gravi infezioni con scarse possibilità di successo terapeutico. 132 132

133
Biofilm in patologia umana
Impianto su superfici mucose Center for Disease Control and Prevention stima che la formazione di biofilm sia implicata almeno nel 65-80% di tutte le infezioni croniche e/o recidivanti. Potera C. Forging a link between biofilms and disease. Science 283: , 1999 Davies D. Understanding biofilm resistance to antibacterial agents. Nature Rev. Drug Discov. 2: , 2003 133 133

134
Riacutizzazioni di BPCO
Biofilm batterici e Infezioni recidivanti Endocarditi: valvole naturali o artificiali Quadri legati a inserzione di cateteri, protesi, lenti,tubi endotracheali Otiti,sinusiti Prostatiti, uretriti ,cistiti infezioni vie biliari Fibrosi cistica Riacutizzazioni di BPCO Costerton JW e al :science 1999;284 : 134 134

135
“ Biofilm “ su superfici inerti
135 135

136
“ Biofilm “ su superfici inerti
I germi che vivono all’interno di questo microcosmo biologico, riescono a comunicare fra loro inviandosi molecole-messaggio non solo per la produzione del biofilm ,regolata geneticamente, ma anche per liberare all’esterno ceppi particolarmente virulenti destinati a colonizzare altre sedi complicando e cronicizzando l’infezione 136 136

137
Attacco al Biofilm Prevenire la formazione di biofilm
Disgregare biofilm, già esistenti Eradicare i microrganismi negli strati più profondi del biofilm E questa flossi infettiva della alte vie si ripercuote anche nelle basse vie favorita anche da fenomeni aspirativi Sethi et al. N Engl J Med 2002; 347: 137 137

138
Attacco al Biofilm La strategia terapeutica deve tener conto che gli antibiotici attualmente in uso sono stati sviluppati e studiati per agire contro batteri planctonici e non organizzati in biofilm E questa flossi infettiva della alte vie si ripercuote anche nelle basse vie favorita anche da fenomeni aspirativi Creson M. Focus: Bacterial Cities TimesDispatch.com 13 Gennaio 2003 138 138

139
Eradicare le cellule planctoniche ! responsabili dell’episodio acuto,
Attacco al Biofilm Eradicare le cellule planctoniche ! responsabili dell’episodio acuto, per ritardare : colonizzazione, potenziale produzione di biofilm recidivare dell’infezione Kobayashi H. Int J Antimicrob Agents 2001 ; 17:351-56 139 139

140
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
Nel loro insieme queste osservazioni indicano che la fase planctonica è quella che favorisce la disseminazione della vita microbica, mentre la fase sessile all’interno del biofilm è favorevole alla sopravvivenza. 140 140

141
Altre strategie batteriche Il biofilm come fattore di patogenicità e virulenza
Lo sviluppo del biofilm è condizionato dalla presenza nell’ambiente di un fattore prodotto dagli stessi microorganismi autoinducente che a sua volta dipende dal numero dei batteri, il fenomeno è noto come quorum sensing 141 141

142
Quorum Sensing Quando la popolazione batterica raggiunge una concentrazione critica, una molecola dà il segnale per lo sviluppo del biofilm Nel biofilm i batteri continuano a comunicare tra loro per controllare l’equilibrio del biofilm e la produzione di fattori di virulenza 142 142

143
Espressione di un gene in funzione della densità dei batteri Quorum sensing
E’ stato identificato nel 1970 su un batterio luminescente il Vibrio fischeri che colonizza un cefalopodo. Il microorganismo è presente in mare ad una concentrazione di circa 100 cell/ml. Mentre sull’organo luminoso del pesce raggiunge 1010 cell/ml e diviene luminescente. 143 143

144
Espressione di un gene in funzione della densità dei batteri Quorum sensing
I batteri elaborano un prodotto specie specifico (omoserinalattone) diffusibile nell’ambiente ove permane e si accumula. Quando la concentrazione raggiunge un limite condizionato dal numero dei batteri presenti si ha induzione del gene bersaglio. 144 144
 diffusibile nell’ambiente ove permane e si accumula. Quando la concentrazione raggiunge un limite condizionato dal numero dei batteri presenti si ha induzione del gene bersaglio")
145
Quorum sensing Il sistema detto luxI~luxR è stato identificato in molte specie ed è tutto da valorizzare. E’ un sistema molto economico per rivelare la propria presenza solo quando il numero è sufficientemente alto per avere tutti i vantaggi (nutrimenti, trasferimento di geni, fattori di patogenicità e virulenza, ecc.) 145 145
")
146
Quorum sensing Applicazioni
E’ stato dimostrato che se in P.aeruginosa si inattiva il gene las il microorganismo perde i suoi caratteri di patogenicità. Si possono rendere innocui i patogeni eliminando il loro sistema quorum sensing? 146 146

147
Quorum sensing Applicazioni
Trovare composti che funzionano come gli autoinduttori ma bloccano il gene Furanone è stato trovato in un’alga ha dimostrato di interferire con il quorum sensing di Serratia liquefaciens. 147 147

148
I Gram-positivi causano effetti simili ma usano mediatori diversi

149
La scoperta dei sistemi quorum sensing nei batteri gram-negativi ha non solo approfondito le nostre conoscenze sulla patogenesi delle infezioni causate da questi organismi ma potrebbe anche provvedere ai mezzi per il trattamento di queste comuni ed importanti infezioni Hartman and Wise 2001 149 149

150
E’ noto che in natura i batteri si presentano con caratteristiche molto diverse da quelle che mostrano in coltura in laboratorio L’ambiente naturale può essere sfavorevole alla crescita e alla sopravvivenza di molti batteri, specialmente di quelli adattati a moltiplicarsi nell’uomo o negli animali Questi microorganismi hanno sviluppato risposte sofisticate alle variazioni ambientali 150

151
Aumento della resistenza agli agenti antimicrobici
Quando le condizioni ambientali non sono idonee alla crescita, i batteri entrano in una fase programmata di sviluppo che risulta in uno stato metabolico meno attivo e rende il batterio più resistente. Aumento della resistenza agli agenti antimicrobici Cambiamenti nell’idrofobicità di superficie e nelle capacità adesive Cambiamenti a carico degli acidi grassi di membrana, degli aa della parete Cambiamento di forma Topologia del cromosoma 151

152
152

153
I BATTERI SI ADATTANO ALLE VARIAZIONI AMBIENTALI MEDIANTE
Induzione della sintesi di sistemi di cattura dei nutrienti presenti in concentrazioni ridotte Modificazione della sintesi di alcuni enzimi allo scopo di utilizzare efficientemente i nutrienti presenti in concentrazioni ridotte Modulazione del tasso di incorporazione dei nutrienti presenti in eccesso Riorganizzazione di alcune vie metaboliche allo scopo di evitare possibili blocchi Coordinamento dei tassi di sintesi per mantenere la crescita bilanciata 153

154
VBNC viable but non culturable (Xu et al. 1982)
I batteri in stato VBNC sono metabolicamente attivi ma non sono in grado di attuare i processi divisionali che portano alla formazione di colonia sui terreni di coltura normalmente utilizzati per il loro rilevamento 154

155
FATTORI CHE INFLUENZANO L’ATTIVAZIONE DELLO STATO VBNC
Scarsità di nutrienti Temperatura Luce Salinità Pressione idrostatica Ossigeno 155

156
ELENCO DI ALCUNI BATTERI CHE POSSONO ATTIVARE LO STATO VBNC
Aeromonas Agrobacterium Alcaligenes Campylobacter Enterobacter Enterococcus Escherichia Helicobacter Klebsiella Legionella Micrococcus Pseudomonas Rhizobium Salmonella Shigella Vibrio Yersinia 156

157
SAGGI UTILIZZATI PER LA DIMOSTRAZIONE DELLA VITALITA’ BATTERICA
Conta Vitale Diretta (DVC) Immunofluorescenza - Conta Vitale Diretta (DFA-DVC) Colorazione con cloruro di tetrazolio (CTC) Colorazione con “Live/Dead bacLight Viability kit” Ricerca di RNA messaggero (RT-PCR) Ricerca di antigeni specifici dello stato VBNC 157
 Immunofluorescenza - Conta Vitale Diretta (DFA-DVC) Colorazione con cloruro di tetrazolio (CTC) Colorazione con Live/Dead bacLight Viability kit Ricerca di RNA messaggero (RT-PCR) Ricerca di antigeni specifici dello stato VBNC")
158
Le cellule non coltivabili (VBNC, stressate) possono rappresentare un problema per la salute pubblica se non rilevate con saggi adeguati 158

159
159

160
160

161
161

162
162

163
163

164
Filamentation of bacteria in response to environmental cues.
a | Innate immune cues. Uropathogenic Escherichia coli (UPEC) reside intracellularly within bladder epithelial cells. A small number of intracellular bacteria respond to the activation of host immune effectors by filamentation. Epithelial-cell death accompanies bacterial growth, thereby resulting in the exposure of filamentous and bacillary organisms on the surface. The filamentous form is resistant to neutrophil phagocytosis. The recovery from filamentation results in the invasion of naive epithelial cells to begin the process again. 164
 reside intracellularly within bladder epithelial cells. A small number of intracellular bacteria respond to the activation of host immune effectors by filamentation. Epithelial-cell death accompanies bacterial growth, thereby resulting in the exposure of filamentous and bacillary organisms on the surface. The filamentous form is resistant to neutrophil phagocytosis. The recovery from filamentation results in the invasion of naive epithelial cells to begin the process again")
165
Filamentation of bacteria in response to environmental cues.
b | Predator sensing cues. Marine bacterial populations are made up of multiple species, including the prototypical Flectobacillus spp. Protist grazing on all species stimulates filamentation in Flectobacillus spp. Filamentous forms cannot be grazed by marine protists. Recovery from filamentation results in an alteration in the diversity of bacteria in the environment and the depletion of non-Flectobacillus species 165

166
Filamentation of bacteria in response to environmental cues.
c | Quorum sensing cues. Proteus mirabilis grows as a bacillary form that can count the number of similar species in the vicinity by quorum sensing. If a quorum of organisms is verified, the bacteria respond by initiating the differential gene expression that leads to filamentation. Although a mechanistic reason for filamentation in Proteus spp. Is under debate, the evidence suggests that this morphology leads to enhanced invasion of the urothelium, thereby providing protection from the host immune response. Dissemination into the tissue disperses the swarming filaments. Cell division is restored owing to low levels of quorumsensing molecules in the tissue. 166

167
Filamentation of bacteria in response to environmental cues.
d | Antimicrobial cues. Filamentation occurs if bacteria are exposed to certain β‑lactam antibiotics in vitro and in vivo. Filamentation allows for survival until the antibiotic is diluted or becomes inactive. The restoration of cell division occurs once the antibiotic activity is lost. In some cases, for example, Burkholderia pseudomallei, cell-division capacity is maintained even in the presence of antibiotics of similar and dissimilar classes, which indicates that protection has been conferred to daughter cells. The mechanisms and consequences of this response have not yet been explored. 167

168
168

169
INVASIONE CELLULARE da parte di batteri enteropatogeni
169 169

170
Cos‘è un batterio enteropatogeno invasivo?
I batteri invasivi sono capaci di promuovere il loro ingresso in cellule che non sono fagociti professionisti. I batteri enteroinvasivi raggiungono il lume intestinale, aderiscono alle cellule dell’ospite e superano la barriera naturale, estremamente impermeabile, rappresentata dalle cellule epiteliali Esempi: Yersinia enterocolitica e Yersinia pseudotuberculosis Shigella spp. Salmonella spp. Listeria monocytogenes 170 170

171
Barriere all‘infezione batterica nel tratto gastro-intestinale
171

172
La barriera dell‘epitelio intestinale
172

173
Cellule M: il punto debole, il tallone d‘Achille
Le placche del Peyer (formate da un centro germinativo di linfociti B circondato da linfociti T) si trovano nella sottomucosa dell’intestino tenue. L’epitelio sovrastante le placche presenta cellule M, cellule epiteliali specializzate nel consentire il passaggio di particelle nel tessuto linfoide sottostante. La superficie basolaterale invaginata forma una superficie dove migrano linfociti e macrofagi. Molecole e particelle vangono trasportate nella tasca. 173
 si trovano nella sottomucosa dell’intestino tenue. L’epitelio sovrastante le placche presenta cellule M, cellule epiteliali specializzate nel consentire il passaggio di particelle nel tessuto linfoide sottostante. La superficie basolaterale invaginata forma una superficie dove migrano linfociti e macrofagi. Molecole e particelle vangono trasportate nella tasca")
174
Fattori che determinano se il batterio sarà internalizzato o no:
Natura del recettore sulla cellula ospite Forza dell’interazione Capacità del batterio di inviare segnali che stimolano o antagonizzano l’internalizzazione 174

175
STUDI IN VITRO E IN VIVO Colture cellulari
Diverse linee cellulari sono state usate per studiare l’invasione batterica: sistema eccellente, riproducibile e non complicato, ma con diverse limitazioni Modelli animali Dopo che i dettagli molecolari sono stati determinati in vitro, possono essere poi verificati in modelli animali 175

176
Meccanismi di invasione cellulare
Zipper Trigger Yersinia, Listeria Salmonella, Shigella 176 176

177
Ingresso di Listeria monocytogenes in fagociti non professionisti in vitro
Sono coinvolte almeno due proteine di superficie: Internalina (InlA): proteina di superficie che è ancorata covalentemente alla parete InlB: proteina di superficie legata debolmente alla parete 177
: proteina di superficie che è ancorata covalentemente alla parete. InlB: proteina di superficie legata debolmente alla parete")
178
in vivo I topi non sviluppano una malattia letale dopo infezione per os con Listeria: scarsa traslocazione dei batteri attraverso l’intestino Risultati simili con Listeria wt o mutanti inlA Cellule che esprimono E-cadherin di topo, al contrario di cellule che esprimono E-cadherina umana, non permettono ingresso di Listeria via inlA la specie specificità è dovuta a differenze in un singolo aa Listeria wt causa una malattia sistemica in topi transgenici che esprimono caderina E umana negli enterociti, mentre mutanti InlA sono incapaci di attraversare la barriera intestinale efficientemente 178

179
Model of invasion of the intestinal epithelium by Listeria
179

180
180

181
Motilità dipendente dalla polimerizzazione di actina
Polimerizzazione dell’actina con formazione di code Si muove nel citoplasma Invade le cellule adiacenti ActA: proteina di 90 kDa, determina il reclutamento e l’assemblaggio dei filamenti Localizzata ad una estremità del batterio Velocità: 1um/sec 181

182
Per la fuoriuscita dai vacuoli:
Ingresso nella cellula adiacente e fuoriuscita dal vacuolo secondario I batteri a contatto con la membrana plasmatica continuano a muoversi Producono protusioni che si estendono nella cellula adiacente: listeriopodi Si forma una vescicola con due membrane e la cellula si libera nel citoplasma Listeriolisina PC- PLC: proteina extracellulare 28 kDa fosfolipasi a largo spettro, idrolizza la fosfatidil colina (PC) Metallo proteasi: trasforma il precursore di PC- PLC (33 kDa) in forma attiva (28 kDa) Le due fosfolipasi determinano la lisi di entrambe le membrane cellulari. Non è noto se svolgano ruoli diversi nei due compartimenti che formano il vacuolo Per la fuoriuscita dai vacuoli: Pseudopodi: 40 um 182
 Metallo proteasi: trasforma il precursore di PC- PLC (33 kDa) in forma attiva (28 kDa) Le due fosfolipasi determinano la lisi di entrambe le membrane cellulari. Non è noto se svolgano ruoli diversi nei due compartimenti che formano il vacuolo. Per la fuoriuscita dai vacuoli: Pseudopodi: 40 um")
183
inlA/B plcB hlyA hlyA plcA actA
inlA: internalina; hly: listeriolisina; plcA: PI- PLC (una fosfolipasi); actA: polimerizzazione actina; plcB: PC- PLC (una fosfolipasi) 183
; actA: polimerizzazione actina; plcB: PC- PLC (una fosfolipasi) 183.")
184
184

185
The invasive phenotype of Shigella is dependent on a type III secretion system encoded by a 30 kb region of a 200 kb virulence plasmid VIRF: quando il batterio cresce a 37°C, la proteina plasmidica virF si lega al promotore di virB inducendo l’espressione della proteina virB, anch’essa plasmidica VirB: attiva i promotori ipa, mxi, e spa. IpaA iniettata nel citoplasma dell’ospite favorisce la depolimerizzazione dell’actina accumulata intorno al sito d’ingresso del batterio. Le proteine IpaA servono anche per la lisi del vacuolo Ipa B e C, si integrano nella membrana dell’ospite e promuovono la formazione degli pseudopodi IcsB lisa le membrane dei vacuoli; esposta sulla superficie o secreta IcsA (VirG): una ATPasi localizzata sulla membrana esterna della cellula batterica, in posizione polare IcsA richiama sulla superficie della cellula batterica diverse proteine actino-associate, provocando la polimerizzazione dell’actina dell’ospite e lo spostamento della cellula batterica. mxi membrane excretion of Ipa, and virulence vir genes spa surface presentation of invasion plasmid antigens spa e mxi operon: la loro espressione porta all’assemblaggio del complesso di traslocazione inattivo Mxi-Spa; l’attivazione avviene dopo il contatto con la cellula 185
: una ATPasi localizzata sulla membrana esterna della cellula batterica, in posizione polare IcsA richiama sulla superficie della cellula batterica diverse proteine actino-associate, provocando la polimerizzazione dell’actina dell’ospite e lo spostamento della cellula batterica. mxi membrane excretion of Ipa, and virulence vir genes. spa surface presentation of invasion plasmid antigens. spa e mxi operon: la loro espressione porta all’assemblaggio del complesso di traslocazione inattivo Mxi-Spa; l’attivazione avviene dopo il contatto con la cellula")
186
FATTORI DI VIRULENZA plasmidici
IPa (Invasion-Plasmid-Antigens)-ABCD IpaA (30 kDa) IpaB (62 kDa), IpaC (42 kDa), e IpaD (35 kDa) in condizioni non secretorie sono accumulate nel citoplasma legate ad una proteina di 18 kDa (IpgC) che ne impedisce l’aggregazione e la digestione Ipa B e C, si integrano nella membrana dell’ospite e promuovono la formazione degli pseudopodi IpaA iniettata nel citoplasma dell’ospite favorisce la depolimerizzazione dell’actina accumulata intorno al sito d’ingresso del batterio. Le proteine IpaA servono anche per la lisi del vacuolo VIRF: quando il batterio cresce a 37°C, la proteina plasmidica virF si lega al promotore di virB inducendo l’espressione della proteina virB, anch’essa plasmidica VirB: attiva i promotori ipa, mxi, e spa. 186
-ABCD. IpaA (30 kDa) IpaB (62 kDa), IpaC (42 kDa), e IpaD (35 kDa) in condizioni non secretorie sono accumulate nel citoplasma legate ad una proteina di 18 kDa (IpgC) che ne impedisce l’aggregazione e la digestione. Ipa B e C, si integrano nella membrana dell’ospite e promuovono la formazione degli pseudopodi. IpaA iniettata nel citoplasma dell’ospite favorisce la depolimerizzazione dell’actina accumulata intorno al sito d’ingresso del batterio. Le proteine IpaA servono anche per la lisi del vacuolo. VIRF: quando il batterio cresce a 37°C, la proteina plasmidica virF si lega al promotore di virB inducendo l’espressione della proteina virB, anch’essa plasmidica. VirB: attiva i promotori ipa, mxi, e spa")
187
IcsB lisa le membrane dei vacuoli; esposta sulla superficie o secreta
FATTORI DI VIRULENZA spa surface presentation of invasion plasmid antigens mxi membrane excretion of Ipa, and virulence vir genes. spa e mxi operon: la loro espressione porta all’assemblaggio del complesso di traslocazione inattiva Mxi-Spa; l’attivazione avviene dopo il contatto con la cellula IcsA (VirG): una ATPasi localizzata sulla membrana esterna della cellula batterica, in posizione polare IcsA richiama sulla superficie della cellula batterica diverse proteine actino-associate, provocando la polimerizzazione dell’actina dell’ospite e lo spostamento della cellula batterica. IcsB lisa le membrane dei vacuoli; esposta sulla superficie o secreta 187
: una ATPasi localizzata sulla membrana esterna della cellula batterica, in posizione polare IcsA richiama sulla superficie della cellula batterica diverse proteine actino-associate, provocando la polimerizzazione dell’actina dell’ospite e lo spostamento della cellula batterica. IcsB lisa le membrane dei vacuoli; esposta sulla superficie o secreta")
188
invasion plasmid antigens ipa,
surface presentation of invasion plasmid antigens spa, membrane excretion of Ipa mxi, virulence vir genes. A 37°C, VIRF e VIRB attivano operon spa, mxi e ipa Preinvasion step in the pathogenesis of Shigella spp. Ipa: invasion plasmid antigens; Mxi-Spa: membrane expression of Ipa proteins and surface presentation of antigens 188

189
Early steps in the invasion of Shigella spp into M cells
Early steps in the invasion of Shigella spp into M cells. Ipa: invasion plasmid antigen; Spa: surface presentation of antigens 189

190
190

191
191

192
Cytoskeletal rearrangements induced during Shigella invasion of epithelial cells. Shigella secretes the effector proteins, such as IpaB, IpaC, IpaA and VirA, into host cells. IpaB and IpaC are integrated into the host membrane, and IpaC modulates Cdc42-dependent filopodial formation which, in turn, may cause activation of Rac1 and lamellipodial formation. IpaA binds vinculin, and the resulting IpaA−vinculin complex promotes depolymerization of actin filaments, which is thought to be required for modulation of lamellipodial formation. VirA binds / -tubulin heterodimers and induces MT destabilization. VirA-induced MT destabilization would in turn lead to MT growth and stimulation of the Rac1 activity, and thus evoke the local membrane ruffling. 192

Presentazioni simili
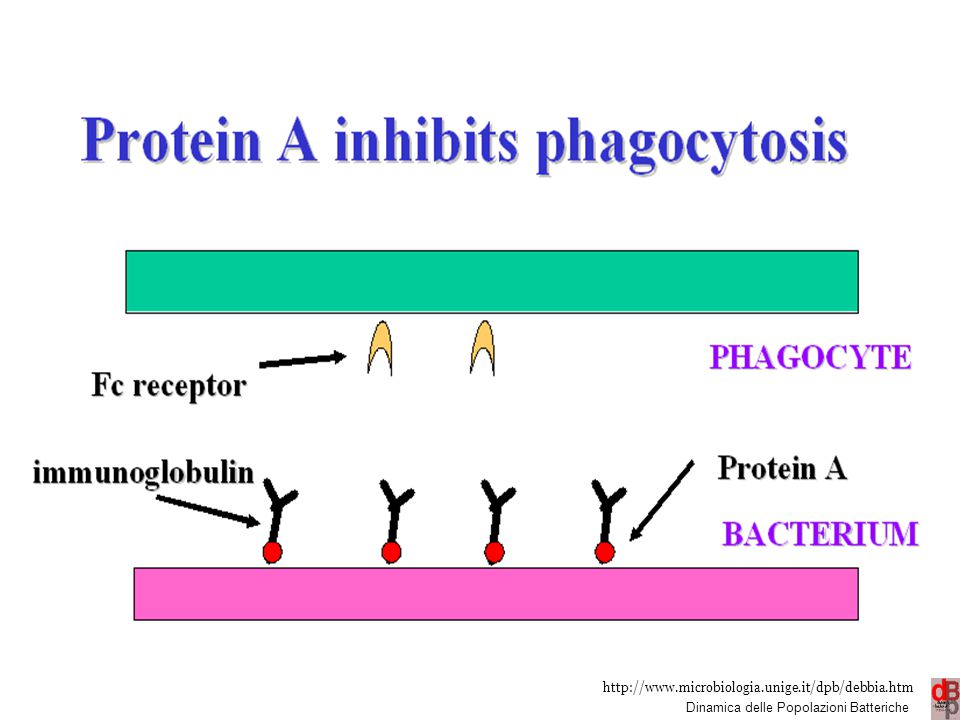




 Costituito da una parte liquida: plasma e.>")



 e loro genomi: rapporto con organismo umano cellule batteriche molto più numerose delle cellule dell’organismo umano.>")





 Pontieri - Elementi di Patologia Generale (ed. Piccin) Majno-Joris - Cellule,>")




