Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
farmaci antiinfiammatori
Infiammazione e farmaci antiinfiammatori

4
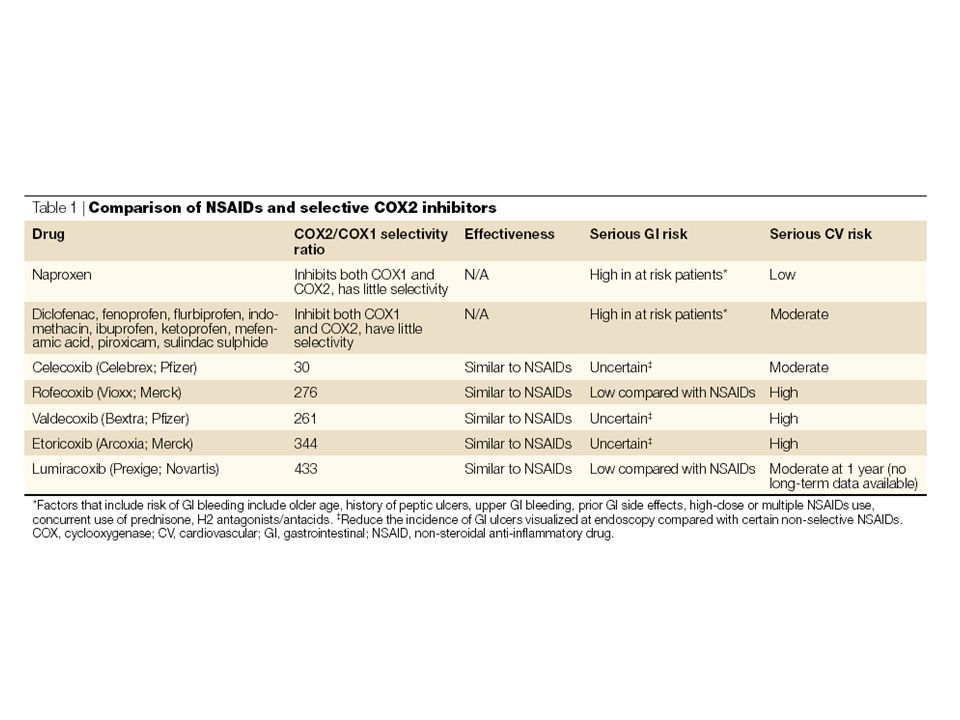
| Prostaglandin biosynthetic cascade
| Prostaglandin biosynthetic cascade. Nonspecific physical and chemical stimuli mobilize release of arachidonic acid from the sn-2 position of membrane phospholipids, permitting metabolism by PGG2/H2 synthases. These enzymes possess both cyclooxygenase (COX) and hydroperoxidase (HOX) activities and catalyze the sequential formation of prostaglandin (PG) endoperoxides. These are then further metabolized by isomerases and synthases (for example PGE synthases), which are expressed with some tissue specificity and generate distinct PGs. These activate distinct G protein coupled receptors which derive from an ancestral E prostanoid (EP) receptor with one exception. The DP2 (also known as CRTH2) belongs to the fMLP receptor superfamily. Adapted with permission from Ref. 16 © Massachusetts Medical Society (2001). COX, cyclooxygenase; DP, PGD2 receptor; EP, PGE2 receptor; FP, PGF2 receptor; HOX, hydroperoxidase; IP, PGI2 receptor; TP, TxA2 receptor.
 and hydroperoxidase (HOX) activities and catalyze the sequential formation of prostaglandin (PG) endoperoxides. These are then further metabolized by isomerases and synthases (for example PGE synthases), which are expressed with some tissue specificity and generate distinct PGs. These activate distinct G protein coupled receptors which derive from an ancestral E prostanoid (EP) receptor with one exception. The DP2 (also known as CRTH2) belongs to the fMLP receptor superfamily. Adapted with permission from Ref. 16 © Massachusetts Medical Society (2001). COX, cyclooxygenase; DP, PGD2 receptor; EP, PGE2 receptor; FP, PGF2 receptor; HOX, hydroperoxidase; IP, PGI2 receptor; TP, TxA2 receptor.")
5
Figure 1 Pathways of prostanoid formation.
Arachidonic acid, often supplied by the actions of phospholipase A2, is converted in a two-step process first to prostaglandin (PG) G2 and then PGH2. PGH2 is the substrate for a range of synthetic enzymes that form the prostanoids; relative amounts of these synthetic enzymes vary widely in different tissues; platelets, for example, contain particularly high amounts of thromboxane A2 (TxA2) synthase, and endothelial cells, prostacyclin (PGI2) synthase. COX, cyclooxygenase. Mitchell JA and Warner TD (2006) COX isoforms in the cardiovascular system: understanding the activities of non-steroidal anti-inflammatory drugs Nature Reviews Drug Discovery 5: 75–86 doi: /nrd1929
 G2 and then PGH2. PGH2 is the substrate for a range of synthetic enzymes that form the prostanoids; relative amounts of these synthetic enzymes vary widely in different tissues; platelets, for example, contain particularly high amounts of thromboxane A2 (TxA2) synthase, and endothelial cells, prostacyclin (PGI2) synthase. COX, cyclooxygenase. Mitchell JA and Warner TD (2006) COX isoforms in the cardiovascular system: understanding the activities of non-steroidal anti-inflammatory drugs Nature Reviews Drug Discovery 5: 75–86 doi: /nrd1929.")
6
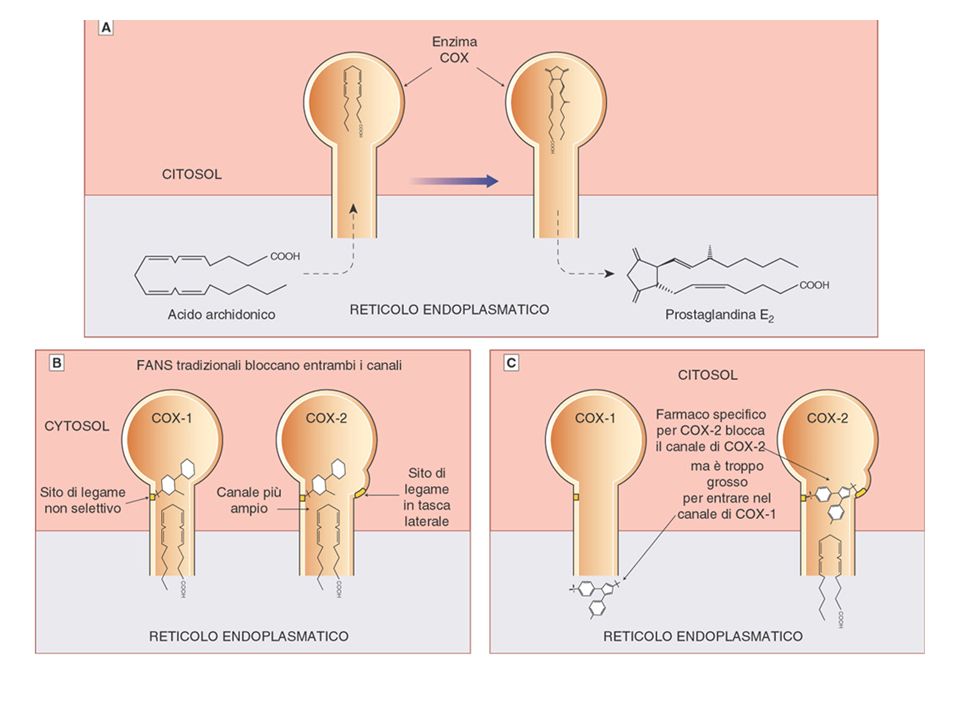
Structural distinction between hydrophobic channels in PGG/H synthase isozymes. Both COX-1 and COX-2 exist as dimers as depicted in a. The arachidonic acid substrate gains access to the cyclooxygenase (COX) catalytic site via a hydrophobic channel. Aspirin irreversibly acetylates a serine residue (Ser529 in human platelet COX-1), close to, but distinct from, the catalytic site. This prevents access of the substrate to the catalytic site of the enzyme. b | The surface of the COX-1 channel is coloured green to indicate the area of lipophilicity with the inhibitor bromo-aspirin bound. c |The specific inhibitor celecoxib is shown bound on the surface of the COX-2 channel. This channel is more bulky and accommodating than that in COX-1, as reflected by a broader substrate specificity, and includes a lateral extension that affords a basis for the design of specific inhibitors. The figures were developed based on the X-ray crystallography data Ref. 145 for COX-1 and from Ref. 146 for COX-2 using methods described in Ref. 147 to model in the inhibitors. Residues crucial to enzyme function as discussed in Ref. 1 are depicted. Part a adapted with permission from Ref. 79 © Massachusetts Medical Society (2001). Panels b and c kindly provided by Guenter Trummlitz (Boehringer Ingleheim, Germany).
 catalytic site via a hydrophobic channel. Aspirin irreversibly acetylates a serine residue (Ser529 in human platelet COX-1), close to, but distinct from, the catalytic site. This prevents access of the substrate to the catalytic site of the enzyme. b | The surface of the COX-1 channel is coloured green to indicate the area of lipophilicity with the inhibitor bromo-aspirin bound. c |The specific inhibitor celecoxib is shown bound on the surface of the COX-2 channel. This channel is more bulky and accommodating than that in COX-1, as reflected by a broader substrate specificity, and includes a lateral extension that affords a basis for the design of specific inhibitors. The figures were developed based on the X-ray crystallography data Ref. 145 for COX-1 and from Ref. 146 for COX-2 using methods described in Ref. 147 to model in the inhibitors. Residues crucial to enzyme function as discussed in Ref. 1 are depicted. Part a adapted with permission from Ref. 79 © Massachusetts Medical Society (2001). Panels b and c kindly provided by Guenter Trummlitz (Boehringer Ingleheim, Germany)..")
7
FUNZIONI FISIOLOGICHE DELLE PROSTAGLANDINE
Termoregolazione Tono della muscolatura liscia bronchiale Pressione sanguigna; Aggregazione piastrinica Azione citoprotettiva sulla mucosa gastrica Funzionalità renale Tono della muscolatura liscia uterina Motilità intestinale Vitalità degli spermatozoi

8
FUNZIONI PATOLOGICHE DELLE PROSTAGLANDINE
Febbre Infarto del miocardio Asma bronchiale Ipertensione Trombosi Ulcere Insufficienza renale Dismenorrea Diarrea Infiammazione Dolore Defenerazione ossea

12
Figure 2 Contrasting vascular effects of prostacyclin and thromboxane A2. b | Thromboxane A2 and prostacyclin have diametrically opposed roles in the cardiovascular system. Thromboxane A2 acts on TP receptors that promote platelet aggregation, adhesion, vasoconstriction and increased vascular smooth-muscle remodelling. Prostacyclin, by contrast, activates IP receptors, leading to reduced aggregation and adhesion of platelets, vasodilation, reduced smooth-muscle remodelling and reduced cholesterol uptake. COX, cyclooxygenase.

13
Figure 2 Contrasting vascular effects of prostacyclin and thromboxane A2. Thromboxane A2 versus prostacyclin balance. Platelets express thromboxane synthase, whereas endothelial cells express prostacyclin synthase. An anti-thrombotic environment can be achieved by exposure to low-dose aspirin, which selectively reduces platelet thromboxane A2 production.

14
Figure 2 | Schematic representation of the two NF-κB signalling pathways. The classical
pathway is depicted on the left. It is activated by TNF-α, IL-1, LPS, CD40 ligand (CD40L) and to a lesser extent by lymphotoxin α/β (LT α/β) and Blys/BAFF. Activation of this pathway depends on the three-subunit IKK holocomplex, which phosphorylates IκBs to induce their degradation. This pathway is crucial for the activation of innate immunity and inflammation, and for inhibition of apoptosis (increased cell survival). The alternative pathway is depicted on the right. It is activated by LT α/β, CD40L and Blys/BAFF, but not by TNF-α, IL-1 or LPS. Activation of this pathway depends on IKK-α homodimers, which induce processing of p100 and nuclear translocation of RelB–p52 dimers. This pathway is crucial for secondary lymphoid organ development, maturation of B cells and adaptive humoral immunity (that is, the production of high-affinity antibodies). IκB, inhibitor of NF-κB; IKK, IκB kinase; IL, interleukin; LPS, lipopolysaccharide; NF-κB, nuclear factor-κB; TNF, tumour-necrosis factor.
 and to. a lesser extent by lymphotoxin α/β (LT α/β) and Blys/BAFF. Activation of this pathway depends. on the three-subunit IKK holocomplex, which phosphorylates IκBs to induce their degradation. This pathway is crucial for the activation of innate immunity and inflammation, and for inhibition. of apoptosis (increased cell survival). The alternative pathway is depicted on the right. It is. activated by LT α/β, CD40L and Blys/BAFF, but not by TNF-α, IL-1 or LPS. Activation of this. pathway depends on IKK-α homodimers, which induce processing of p100 and nuclear. translocation of RelB–p52 dimers. This pathway is crucial for secondary lymphoid organ. development, maturation of B cells and adaptive humoral immunity (that is, the production. of high-affinity antibodies). IκB, inhibitor of NF-κB; IKK, IκB kinase; IL, interleukin; LPS, lipopolysaccharide; NF-κB, nuclear factor-κB; TNF, tumour-necrosis factor.")
15
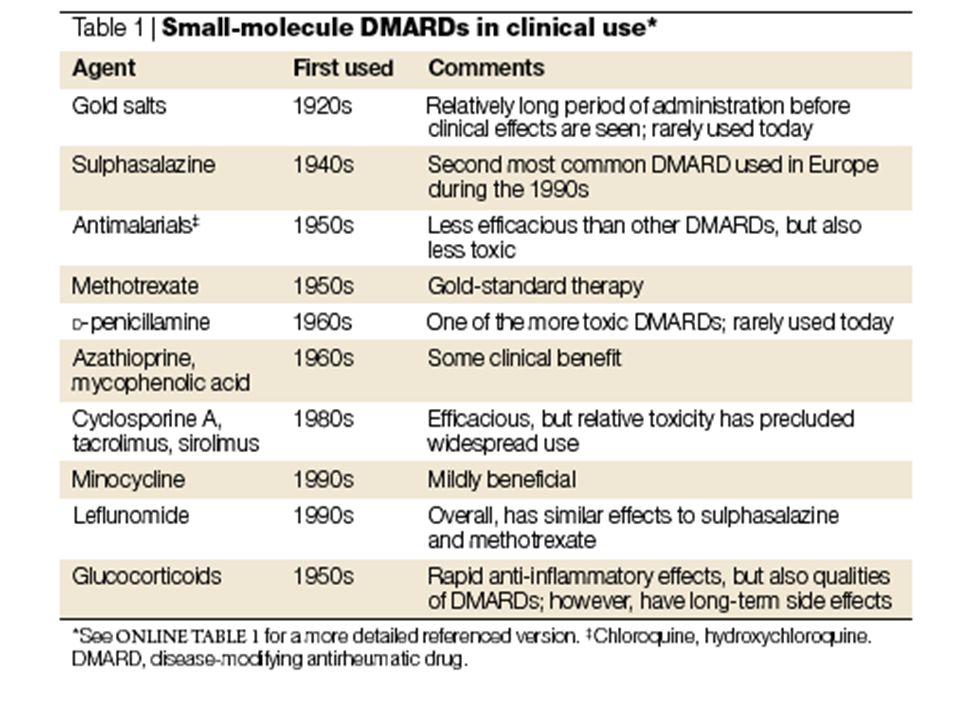
Box 1 | Other tools to interfere with the NF-κB pathway
Although major efforts to develop inhibitors of the nuclear factor-κB (NF-κB) pathway in the pharmaceutical industry have focused on selective IKK-β inhibitors, there are a number of other different potential approaches.As summarized in the figure, targets of these approaches can be categorized into seven different groups on the basis of unique events in regulation of NF-κB proteins: regulation of NF-κB protein expression and binding to DNA (RelA antisense,NF-κB decoy and RNAi); interference with the formation of the IKK complex (NBD peptide,which corresponds to the IKK-γ-binding domain of IKK-β); blockade of IKK-β activation process (TIRAP peptide,which corresponds to the Toll-interleukin-1 receptor (TIR) domain-containing adapter domain (TIRAP),Hsp90 inhibitor: Hsp90 stabilizes RIP proteins that are the components of the TNF-α receptor signalling complex and antioxidants); inhibitors of IKK-β kinase activity; proteasome inhibitors; NF-κB nuclear translocation inhibitors (IκB-αsuper-repressor and SN50: a peptide consisting of the nuclear localization sequence (NLS) of the p50 NF-κB subunit); and last, inhibitors of NF-κB transcriptional activity (glucocorticoids).Given the diverse processes involved in regulation of the NF-κB pathways, a better basic understanding of these crucial signalling pathways will ultimately lead to new avenues in the development of efficient and clinically useful inhibitors.
 pathway in the pharmaceutical industry. have focused on selective IKK-β inhibitors, there are a number of other different potential approaches.As summarized in the. figure, targets of these approaches can be categorized into seven different groups on the basis of unique events in regulation. of NF-κB proteins: regulation of NF-κB protein expression and binding to DNA (RelA antisense,NF-κB decoy and RNAi); interference with the formation of the IKK complex (NBD peptide,which corresponds to the IKK-γ-binding domain of. IKK-β); blockade of IKK-β activation process (TIRAP peptide,which corresponds to the Toll-interleukin-1 receptor (TIR) domain-containing adapter domain (TIRAP),Hsp90 inhibitor: Hsp90 stabilizes RIP proteins that are the components of the. TNF-α receptor signalling complex and antioxidants); inhibitors of IKK-β kinase activity; proteasome inhibitors; NF-κB nuclear translocation inhibitors (IκB-αsuper-repressor and SN50: a peptide consisting of the nuclear localization. sequence (NLS) of the p50 NF-κB subunit); and last, inhibitors of NF-κB transcriptional activity (glucocorticoids).Given. the diverse processes involved in regulation of the NF-κB pathways, a better basic understanding of these crucial signalling. pathways will ultimately lead to new avenues in the development of efficient and clinically useful inhibitors.")
17
EFFETTO DEI SALICILATI SULL’EQUILIBRIO ACIDO-BASE
DOSE TOSSICA DOSE TERAPEUTICA DISACCOPPIAMENTO FOSFORILAZIONE OSSIDATIVA DEPRESSIONE CENTRI RESPIRATORI STIMOLAZIONE CENTRI RESPIRATORI CONSUMO O2 PRODUZIONE CO2 FREQUENZA RESPIRATORIA FREQUENZA RESPIRATORIA COMPENSAZIONE ESCREZIONE RENALE HCO3- ACIDOSI RESPIRATORIA ALCALOSI RESPIRATORIA

25
Figure 1 | Structures of aspirin and NCX4016
Figure 1 | Structures of aspirin and NCX4016. NCX4016 consists of aspirin linked to a 'spacer' by an ester bond. A nitric oxide (NO)-releasing moiety (ONO2) is attached to the spacer.
-releasing moiety (ONO2) is attached to the spacer.")
26
Figure 2 | Metabolism of NO-NSAIDs
Figure 2 | Metabolism of NO-NSAIDs. a | NO-NSAIDs, such as NO-aspirin, can be metabolized by esterases (primarily in the liver and in plasma) to yield the parent drug, plus the spacer attached to the NO-releasing moiety. Subsequent metabolism of the latter leads to the liberation of NO, which can occur many hours after oral administration of the drug. b | Plasma levels of nitrate and nitrite in the rat after oral administration of NCX4016 (50 mg kg-1). Liberation of salicylate from this drug preceded, in a parallel manner, the liberation of NO. NO, nitric oxide; NSAIDs, nonsteroidal anti-inflammatory drugs.
 to yield the parent drug, plus the spacer attached to the NO-releasing moiety. Subsequent metabolism of the latter leads to the liberation of NO, which can occur many hours after oral administration of the drug. b | Plasma levels of nitrate and nitrite in the rat after oral administration of NCX4016 (50 mg kg-1). Liberation of salicylate from this drug preceded, in a parallel manner, the liberation of NO. NO, nitric oxide; NSAIDs, nonsteroidal anti-inflammatory drugs.")
30
Comparison of the NSAID binding sites of COX1 and COX2 after Browner
Comparison of the NSAID binding sites of COX1 and COX2 after Browner. Schematic cartoon (modified from Ref. 142), showing the differences in the NSAID binding sites of COX1 and COX2. Note that the COX2 binding site is more accommodating and is characterized by a 'side pocket' that can accommodate bulky groups such as the methyl sulphonyl moiety of DuP697. COX, cyclooxygenase; NSAID, non-steroidal anti-inflammatory drug.
, showing the differences in the NSAID binding sites of COX1 and COX2. Note that the COX2 binding site is more accommodating and is characterized by a side pocket that can accommodate bulky groups such as the methyl sulphonyl moiety of DuP697. COX, cyclooxygenase; NSAID, non-steroidal anti-inflammatory drug.")
34
Schematic view of a normal joint and its changes in rheumatoid arthritis. a | The synovial joint is composed of two adjacent bony ends each covered with a layer of cartilage, separated by a joint space and surrounded by the synovial membrane and joint capsule. The synovial membrane is normally <100 m thick and the synovial lining (facing the cartilage and bone) consists of a thin (1–3 cells) layer of synoviocytes (type A are macrophage derived, and type B are fibroblast derived); there is no basement membrane. Only a few, if any, mononuclear cells are interspersed in the sublining connective tissue layer, which has considerable vascularity. The synovial membrane covers all intra-articular structures except for cartilage and small areas of exposed bone ('bare areas') and inserts near the cartilage–bone junction. The 'radiographic joint space' (seen in c, which shows a radiograph of the second, third and fourth metacarpophalangeal joints in a normal hand), in contrast to the usually minute 'anatomic joint space', consists of the latter, as well as of the two neighbouring, radiotranslucent portions of cartilage covering the non-translucent subchondral bone. b | Like many other forms of arthritis, rheumatoid arthritis (RA) is initially characterized by an inflammatory response of the synovial membrane ('synovitis') that is conveyed by a transendothelial influx and/or local activation of a variety of mononuclear cells, such as T cells, B cells, plasma cells, dendritic cells, macrophages, mast cells, as well as by new vessel formation. The lymphoid infiltrate can be diffuse or, commonly, form lymphoid-follicle-like structures. The lining layer becomes hyperplastic (it can have a thickness of >20 cells) and the synovial membrane expands and forms villi. However, in addition, the hallmark of RA is bone destruction (seen in d, which shows three metacarpophalangeal joints from a hand radiograph of a patient with established RA: the joint spaces have narrowed or disappeared as a sign of cartilage degradation and destructions of the adjacent bone, also termed 'erosions', have occurred). The destructive portion of the synovial membrane is termed 'pannus', and the destructive cellular element is the osteoclast; destruction mostly starts at the cartilage–bone–synovial membrane junction. Bone repair by osteoblasts usually does not occur in active RA. Polymorphonuclear leukocytes are found in high numbers in the joint fluid, but very rarely are seen in the synovial membrane, suggesting very rapid transgression from blood to the joint space. The neutrophils' enzymes, together with enzymes secreted by synoviocytes and chondrocytes, lead to cartilage degradation.
 consists of a thin (1–3 cells) layer of synoviocytes (type A are macrophage derived, and type B are fibroblast derived); there is no basement membrane. Only a few, if any, mononuclear cells are interspersed in the sublining connective tissue layer, which has considerable vascularity. The synovial membrane covers all intra-articular structures except for cartilage and small areas of exposed bone ( bare areas ) and inserts near the cartilage–bone junction. The radiographic joint space (seen in c, which shows a radiograph of the second, third and fourth metacarpophalangeal joints in a normal hand), in contrast to the usually minute anatomic joint space , consists of the latter, as well as of the two neighbouring, radiotranslucent portions of cartilage covering the non-translucent subchondral bone. b | Like many other forms of arthritis, rheumatoid arthritis (RA) is initially characterized by an inflammatory response of the synovial membrane ( synovitis ) that is conveyed by a transendothelial influx and/or local activation of a variety of mononuclear cells, such as T cells, B cells, plasma cells, dendritic cells, macrophages, mast cells, as well as by new vessel formation. The lymphoid infiltrate can be diffuse or, commonly, form lymphoid-follicle-like structures. The lining layer becomes hyperplastic (it can have a thickness of >20 cells) and the synovial membrane expands and forms villi. However, in addition, the hallmark of RA is bone destruction (seen in d, which shows three metacarpophalangeal joints from a hand radiograph of a patient with established RA: the joint spaces have narrowed or disappeared as a sign of cartilage degradation and destructions of the adjacent bone, also termed erosions , have occurred). The destructive portion of the synovial membrane is termed pannus , and the destructive cellular element is the osteoclast; destruction mostly starts at the cartilage–bone–synovial membrane junction. Bone repair by osteoblasts usually does not occur in active RA. Polymorphonuclear leukocytes are found in high numbers in the joint fluid, but very rarely are seen in the synovial membrane, suggesting very rapid transgression from blood to the joint space. The neutrophils enzymes, together with enzymes secreted by synoviocytes and chondrocytes, lead to cartilage degradation..")
39
Antibodies are produced by plasma cells and some memory B cells
Antibodies are produced by plasma cells and some memory B cells. They are complex, multi-domain adapter molecules, used by the immune system to neutralize and/or destroy invading microorganisms and their products (antigens) by uniting the antigen with various effector molecules such as complement and leukocytes bearing fragment crystalizable- (Fc) receptors. An antibody comprises two heavy and two light chains (a). At one end of the Y-shaped antibody molecule, two identical variable (V) regions have a molecular structure that is highly complementary to the target antigen. The single constant (C) region at the other end determines the fate of the bound antigen. Within the V regions, three short polypeptide segments on the heavy chain and three on the light chain form the hypervariable or complementarity-determining regions (CDRs), which dictate the precise antigen-binding characteristics of the antibody. In the three-dimensional structure of an antibody molecule, the three heavy- and three light-chain CDRs lie close together and form the antigen-binding site. A technique for producing monoclonal antibodies (mAbs) was first published by Kohler and Milstein in Antibody-producing B cells from an immunized animal were 'fused' to a cancer cell line, resulting in 'hybridomas': immortal cells that secreted antibodies of a single specificity (that is, mAbs). It therefore became possible to produce and manufacture unlimited quantities of a mAb against a desired target antigen. Subsequent application of genetic engineering techniques to genes encoding antibody V and C regions have overcome many limitations associated with traditional hybridoma technology, yielding chimeric and, subsequently, humanized and fully human mAbs. Genetic engineering has also led to the development of Fc fusion proteins in which the extracellular domain of a non-antibody ligand-binding moiety (for example, a cell-surface receptor) is fused to the hinge, CH2 and CH3 domains of an immunoglobulin C region (b). 39
 by uniting the antigen with various effector molecules such as complement and leukocytes bearing fragment crystalizable- (Fc) receptors. An antibody comprises two heavy and two light chains (a). At one end of the Y-shaped antibody molecule, two identical variable (V) regions have a molecular structure that is highly complementary to the target antigen. The single constant (C) region at the other end determines the fate of the bound antigen. Within the V regions, three short polypeptide segments on the heavy chain and three on the light chain form the hypervariable or complementarity-determining regions (CDRs), which dictate the precise antigen-binding characteristics of the antibody. In the three-dimensional structure of an antibody molecule, the three heavy- and three light-chain CDRs lie close together and form the antigen-binding site. A technique for producing monoclonal antibodies (mAbs) was first published by Kohler and Milstein in Antibody-producing B cells from an immunized animal were fused to a cancer cell line, resulting in hybridomas : immortal cells that secreted antibodies of a single specificity (that is, mAbs). It therefore became possible to produce and manufacture unlimited quantities of a mAb against a desired target antigen. Subsequent application of genetic engineering techniques to genes encoding antibody V and C regions have overcome many limitations associated with traditional hybridoma technology, yielding chimeric and, subsequently, humanized and fully human mAbs. Genetic engineering has also led to the development of Fc fusion proteins in which the extracellular domain of a non-antibody ligand-binding moiety (for example, a cell-surface receptor) is fused to the hinge, CH2 and CH3 domains of an immunoglobulin C region (b). 39.")
40
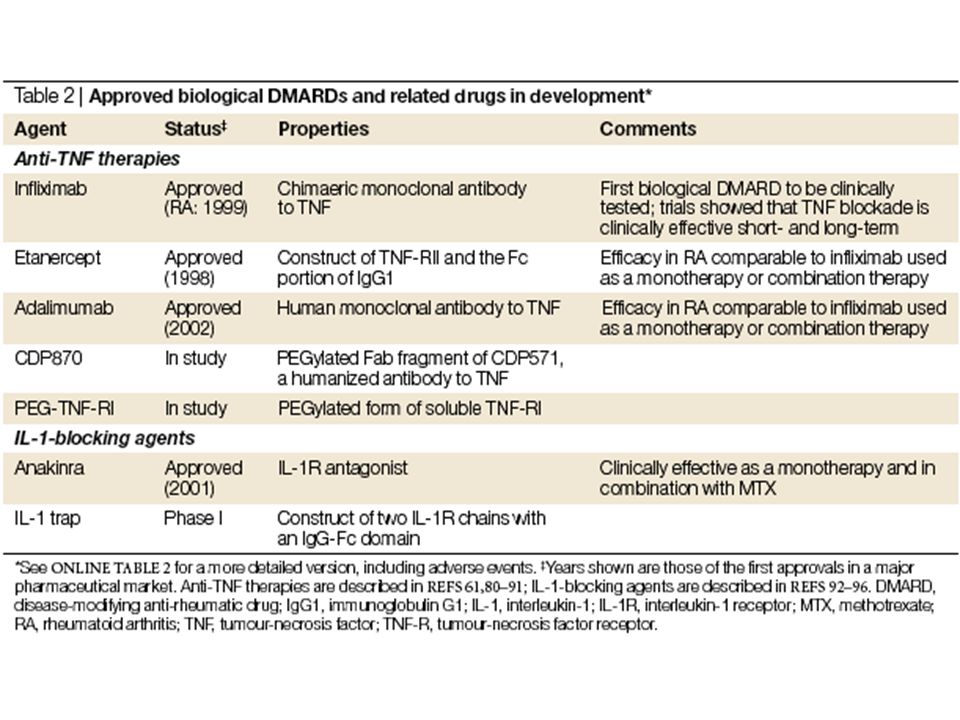
Figure 1 | Adalimumab (D2E7).
a | Simplified illustration of the cytokine cascade that leads to rheumatoid arthritis. Activated T cells invade the synovial membrane and release cytokines including interleukin (IL)-2, interferon (IFN)- , tumour-necrosis factor (TNF)- , TNF- and IL-3. This causes synovial macrophages and fibroblasts to produce the inflammatory cytokines TNF- , IL-1 and IL-6, which leads to complex signal-transduction cascades and to the production of molecules involved in tissue degradation and those resulting in the resorption and destruction of bone. b | Adalimumab is produced by phage-display technology, resulting in an antibody with human-derived heavy- and light-chain variable regions, and human immunoglobulin (Ig) G1: constant regions. Infliximab is chimeric: three-quarters human (constant region; Fc) and one quarter mouse (variable region; Fv). GM-CSF, granulocyte–macrophage colony-stimulating factor; IL-1Ra, IL-1 receptor antagonist; TNFR, TNF receptor. Adapted from Ref. 2 © Macmillan Magazines Ltd (2002).
-2, interferon (IFN)- , tumour-necrosis factor (TNF)- , TNF- and IL-3. This causes synovial macrophages and fibroblasts to produce the inflammatory cytokines TNF- , IL-1 and IL-6, which leads to complex signal-transduction cascades and to the production of molecules involved in tissue degradation and those resulting in the resorption and destruction of bone. b | Adalimumab is produced by phage-display technology, resulting in an antibody with human-derived heavy- and light-chain variable regions, and human immunoglobulin (Ig) G1: constant regions. Infliximab is chimeric: three-quarters human (constant region; Fc) and one quarter mouse (variable region; Fv). GM-CSF, granulocyte–macrophage colony-stimulating factor; IL-1Ra, IL-1 receptor antagonist; TNFR, TNF receptor. Adapted from Ref. 2 © Macmillan Magazines Ltd (2002).")
42
a | Potential physiological effects of tumour-necrosis factor- (TNF)
a | Potential physiological effects of tumour-necrosis factor- (TNF). Cell I produces soluble TNF and also expresses membrane-bound TNF (from which soluble TNF is cleaved). Soluble TNF binds to type I and type II TNF receptors on TNF-responsive cell types (cell III). The net effect of TNF depends on the relative proportion of type I and type II receptors, and can be activating or pro-apoptotic. Cell-surface TNF can also bind and signal to adjacent cells (cell II), and soluble TNF can also act in an autocrine manner if cell I also expresses TNF receptors (not illustrated). 42
. Cell I produces soluble TNF and also expresses membrane-bound TNF (from which soluble TNF is cleaved). Soluble TNF binds to type I and type II TNF receptors on TNF-responsive cell types (cell III). The net effect of TNF depends on the relative proportion of type I and type II receptors, and can be activating or pro-apoptotic. Cell-surface TNF can also bind and signal to adjacent cells (cell II), and soluble TNF can also act in an autocrine manner if cell I also expresses TNF receptors (not illustrated). 42.")
43
b | The 'simple' scenario in which the effects of soluble and membrane-bound TNF are prevented by neutralization with monoclonal antibody (mAb) or soluble receptor, and receptor-expressing cells II and III consequently remain in a basal state. 43

44
c | Inhibition of TNF results in cell death, either through cytokine deprivation (cells II and III); through antibody-facilitated cell death (complement-mediated cytotoxicity (CDC) or antibody dependent cell-mediated cytotoxicity (ADCC)); or through reverse signalling. Both of the latter mechanisms affect cells expressing membrane TNF. 44

46
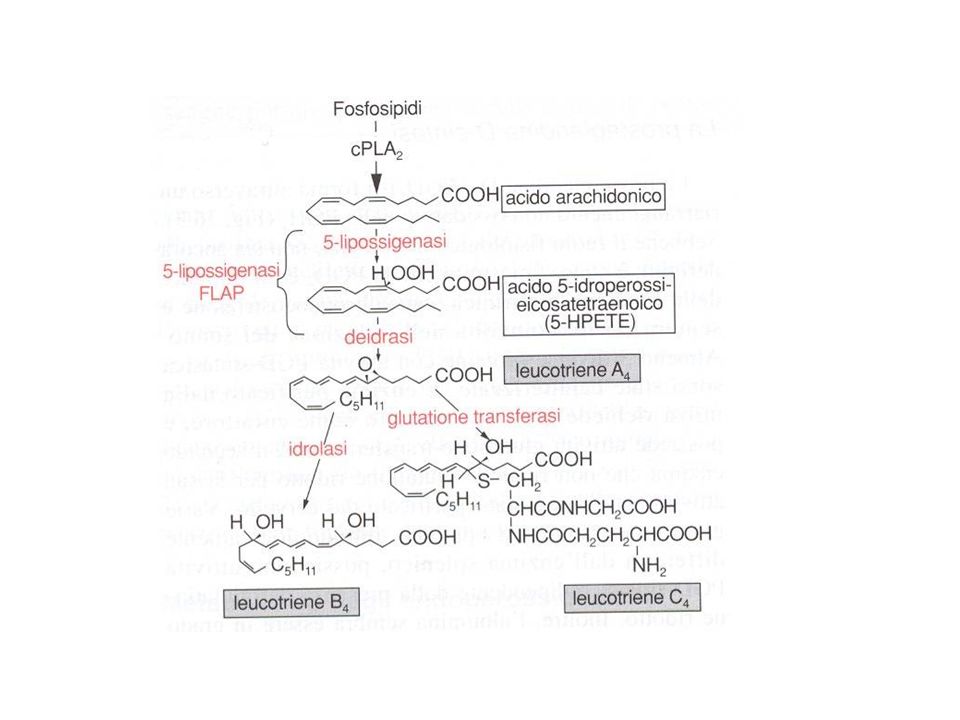
Figure 1 | Leukotriene biosynthetic cascade
Figure 1 | Leukotriene biosynthetic cascade. Various stimuli mobilize release of arachidonic acid from the sn-2 position of membrane glycerophospholipids via cytosolic phospholipase A2(cPLA2), allowing metabolism by 5-lipoxygenase (5-LO). 5-LO catalyses the conversion from arachidonic acid to 5-hydroperoxyeicosatetraenoic acid, while 5-LO-activating protein (FLAP) acts as an apparent chaperone and facilitates the transfer of substrate to the enzyme. A complex of 5-LO, FLAP, leukotriene C4 (LTC4) synthase and possibly LTA4 hydrolase forms at the nuclear envelope with only the synthase and FLAP integrally associated in the membrane. Tissue specificity of expression yield either LTB4 or the cysteinyl leukotriene LTC4. Specific transporters carry these molecules outside the cell where metabolism to LTD4 and LTE4 can occur and activation of cell-surface GPROTEINCOUPLED RECEPTORS takes place. 5-HPETE, hydroperoxyeicosatetraenoic acid; BLT, leukotriene B receptor; CysLT, cysteinyl leukotriene receptor.
, allowing metabolism by 5-lipoxygenase (5-LO). 5-LO catalyses the conversion from. arachidonic acid to 5-hydroperoxyeicosatetraenoic acid, while 5-LO-activating protein (FLAP) acts as an apparent chaperone and facilitates the transfer of substrate to the enzyme. A complex. of 5-LO, FLAP, leukotriene C4 (LTC4) synthase and possibly LTA4 hydrolase forms at the nuclear. envelope with only the synthase and FLAP integrally associated in the membrane. Tissue. specificity of expression yield either LTB4 or the cysteinyl leukotriene LTC4. Specific transporters. carry these molecules outside the cell where metabolism to LTD4 and LTE4 can occur and. activation of cell-surface GPROTEINCOUPLED RECEPTORS takes place. 5-HPETE, hydroperoxyeicosatetraenoic. acid; BLT, leukotriene B receptor; CysLT, cysteinyl leukotriene receptor.")
Presentazioni simili



























 è una malattia respiratoria cronica prevenibile e trattabile variabilmente associata con significativi.>")


