Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
I BIOCOMPOSTI Acqua H2O È il composto max abbondante in esseri viventi e costituisce praticamente il solvente universale del protoplasma (complesso di sostanze che compongono la cellula vivente). È un solvente polare. La sua molecola ha forma di triangolo.
. È un solvente polare. La sua molecola ha forma di triangolo.")
2
Caratteristiche generali Proprietà chimico-fisiche
Nome IUPAC monossido di diidrogeno, idrossido di idrogeno, ossano Caratteristiche generali Massa molecolare (uma) g/mol Aspetto liquido incolore, inodore Proprietà chimico-fisiche Densità liq (g/cm3, in c.s.) a K (4 °C) Temperatura di fusione (K) (0.00°C) Temperatura di ebollizione (K) ( °C) Punto triplo K (0.01 °C) Pa Punto critico 647 K (374 °C) × 107 Pa Tensione di vapore (Pa) a K Calore specifico Cp,m(J·K-1mol-1) 75.3
 g/mol. Aspetto. liquido incolore, inodore. Proprietà chimico-fisiche. Densità liq (g/cm3, in c.s.) a K (4 °C) Temperatura di fusione (K) (0.00°C) Temperatura di ebollizione (K) ( °C) Punto triplo K (0.01 °C) Pa. Punto critico. 647 K (374 °C) × 107 Pa. Tensione di vapore (Pa) a K Calore specifico. Cp,m(J·K-1mol-1)")
3
Dato che l'ossigeno ha una elettronegatività maggiore, il vertice della molecola ospita una parziale carica negativa, mentre le estremità recano una parziale carica elettrica positiva (dipolo elettrico). O H d- d+ Le cariche fanno sì che le molecole vengano attratte reciprocamente l'una dall'altra (legame idrogeno) e spiega molte delle proprietà fisiche tipiche dell'acqua come p. es. i relativamente alti punto di fusione e di ebollizione, è infatti richiesta una maggiore energia (rispetto a sostanze meno polari) per rompere i legami idrogeno che tengono unite le molecole le une alle altre (es. il metano CH4, simile per PM ma incapace di formare legami idrogeno, è un gas a temperatura ambiente).
 e spiega molte delle proprietà fisiche tipiche dell acqua come p. es. i relativamente alti punto di fusione e di ebollizione, è infatti richiesta una maggiore energia (rispetto a sostanze meno polari) per rompere i legami idrogeno che tengono unite le molecole le une alle altre (es. il metano CH4, simile per PM ma incapace di formare legami idrogeno, è un gas a temperatura ambiente).")
4
L’acqua è parzialmente dissociata
2H2O ⇄ H3O+ + OH- Kw= 10-14 pH=cologaritmo della attività degli ioni idrossonio Attività varia da zero ad 1 e quindi cologaritmo da infinito a zero In pratica, concentrazione minima ioni H+=10-14 e quindi pH varia da zero a 14 in acqua

5
Pressione osmotica La pressione osmotica si sviluppa quando una membrana semipermeabile separa due soluzioni una delle quali contiene un soluto che non può passare attraverso la membrana, come accade nel caso di una cellula. L'acqua può attraversare la membrana e lo farà passando nel compartimento dove è presente il soluto non permeabile nel tentativo di uguagliare le attività ad entrambi i lati della membrana. Questo causa un aumento della colonna d'acqua sul lato che contiene il soluto. Si definisce pressione osmotica (p) la pressione da esercitare per impedire che la colonna d'acqua si innalzi. h
 la pressione da esercitare per impedire che la colonna d acqua si innalzi. h.")
6
Integratori salini, in genere isotonici.
Per una cellula, se l'acqua entra nella cellula la soluzione è detta Ipotonica cioè la concentrazione delle sostanze disciolte è minore nella soluzione che nella cellula. La cellula tenderà ad espandersi ed eventualmente scoppiare. Se l'acqua esce dalla cellula, la soluzione è detta ipertonica cioè concentrazione delle sostanze disciolte è maggiore nella soluzione che nella cellula. La cellula si contrarrà. Integratori salini, in genere isotonici. Come mai naufrago in mare muore di sete?

8
Dove a1 è l’attività del solvente
Attraverso varie ipotesi semplificative, per P si può arrivare alla espressione di Vant’Hoff: Dove ci (i1) è è la concentrazione dell’i-esimo soluto esempio: acqua di mare (35 g/L di NaCl)
 è è la concentrazione dell’i-esimo soluto. esempio: acqua di mare (35 g/L di NaCl)")
9
Cationi: Na+, K+, Ca2+, Mg2+, …
Sali minerali I più comuni ioni di sali inorganici presenti in soluzione in forma dissociata sono: Cationi: Na+, K+, Ca2+, Mg2+, … Anioni: Cl-, NO3-, SO42-, PO43-, CO32-, …

10
Amminoacidi Gli amminoacidi (o aminoacidi) sono molecole organiche che nella loro struttura recano sia il gruppo funzionale dell'ammina (-NH2) sia quello dell'acido carbossilico (-COOH). In biochimica il termine amminoacidi si riferisce più spesso agli L-α-amminoacidi, di formula generica NH2CHRCOOH, cioè quelli il cui gruppo amminico ed il cui gruppo carbossilico sono legati allo stesso atomo di carbonio (chiamato carbonio α) in configurazione L (con l'unica esclusione della glicina achirale in cui -R = -H).
 sono molecole organiche che nella loro struttura recano sia il gruppo funzionale dell ammina (-NH2) sia quello dell acido carbossilico (-COOH). In biochimica il termine amminoacidi si riferisce più spesso agli L-α-amminoacidi, di formula generica NH2CHRCOOH, cioè quelli il cui gruppo amminico ed il cui gruppo carbossilico sono legati allo stesso atomo di carbonio (chiamato carbonio α) in configurazione L (con l unica esclusione della glicina achirale in cui -R = -H).")
11
Una molecola che ammette un'immagine speculare non sovrapponibile a sé è detta chirale. Al contrario, una molecola che invece è sovrapponibile alla propria immagine speculare è detta achirale. Esempi di chiralità sono una mano - destra o sinistra - o una vite, che può avere il filetto che ruota in senso orario o antiorario. Per distinguere due enantiomeri tali per la presenza di un atomo di carbonio con quattro sostituenti diversi, la IUPAC assegna agli isomeri i prefissi (R) (dal latino rectus, destro) ed (S) (dal latino sinister, sinistro); per stabilire quale dei due sia l'isomero (R) e quale l'isomero (S) si segue la regola ideata da Cahn, Ingold e Prelog: Esempio di applicazione della regola Cahn-Ingold-Prelog si ordinano i quattro gruppi legati all'atomo di carbonio "stereogenico" dando priorità maggiore al gruppo che ha direttamente legato all'atomo di carbonio "stereogenico" l'atomo dal peso atomico maggiore; se due gruppi sono legati all'atomo di carbonio "stereogenico" con atomi di uno stesso elemento si considerano i due successivi, se anche questi sono uguali si continua lungo le catene fino a quando viene individuata una differenza. A parità di atomi legati, legami tripli e doppi hanno priorità su quelli semplici. Prendendo ad esempio l'acido lattico (vedi figura sopra; gli atomi rappresentati in nero sono quelli di carbonio, quelli in rosso sono di ossigeno, quelli in bianco di idrogeno), i quattro gruppi sono, in ordine di priorità decrescente -OH, -COOH, -CH3, -H si osserva l'atomo di carbonio "stereogenico" mantenendo dietro di esso il gruppo a priorità più bassa; si osserva quindi se la sequenza delle priorità degli altri tre, in ordine decrescente, segue una rotazione oraria o antioraria se la rotazione è oraria si assegna il prefisso (R) se la rotazione è antioraria si assegna il prefisso (S) I due modelli riportati in figura sono quindi, rispettivamente, quelli dell'acido (R)-lattico e dell'acido (S)-lattico. La scritta "acido (RS)-lattico" ne indica sinteticamente il racemo. L'indicazione (R) o (S) deriva dall'applicazione della regola sopra indicata e non è in relazione con il senso della rotazione del piano della luce polarizzata, unica proprietà fisica che distingue i due enantiomeri. Quando il senso della rotazione del piano della luce polarizzata provocato da ciascun enantiomero è noto, questi può essere indicato nel nome del composto con i prefissi (+) (rotazione in senso orario, enantiomero destrogiro) o (—) (rotazione in senso antiorario, enantiomero levogiro). I nomi completi dei due modelli riportati in figura sono quindi, rispettivamente, acido (R)-(—)-lattico e acido (S)-(+)-lattico. Se vi sono due o più atomi di carbonio "stereogenici", ad ognuno di essi viene assegnato il prefisso (R) o (S) in base alle regole sopra esposte. Per gli zuccheri ed altre molecole di interesse biochimico, data la complessità di molte di queste, non si fa uso dei prefissi (R) ed (S). La distinzione in nomenclatura viene fatta assegnando i prefissi D ed L a seconda che i sostituenti legati all'atomo di carbonio asimmetrico più distante da quello che lega il gruppo carbonilico abbiano disposizione simile a quella della L-gliceraldeide o a quella della D-gliceraldeide.
 (dal latino rectus, destro) ed (S) (dal latino sinister, sinistro); per stabilire quale dei due sia l isomero (R) e quale l isomero (S) si segue la regola ideata da Cahn, Ingold e Prelog: Esempio di applicazione della regola Cahn-Ingold-Prelog. si ordinano i quattro gruppi legati all atomo di carbonio stereogenico dando priorità maggiore al gruppo che ha direttamente legato all atomo di carbonio stereogenico l atomo dal peso atomico maggiore; se due gruppi sono legati all atomo di carbonio stereogenico con atomi di uno stesso elemento si considerano i due successivi, se anche questi sono uguali si continua lungo le catene fino a quando viene individuata una differenza. A parità di atomi legati, legami tripli e doppi hanno priorità su quelli semplici. Prendendo ad esempio l acido lattico (vedi figura sopra; gli atomi rappresentati in nero sono quelli di carbonio, quelli in rosso sono di ossigeno, quelli in bianco di idrogeno), i quattro gruppi sono, in ordine di priorità decrescente -OH, -COOH, -CH3, -H si osserva l atomo di carbonio stereogenico mantenendo dietro di esso il gruppo a priorità più bassa; si osserva quindi se la sequenza delle priorità degli altri tre, in ordine decrescente, segue una rotazione oraria o antioraria. se la rotazione è oraria si assegna il prefisso (R) se la rotazione è antioraria si assegna il prefisso (S) I due modelli riportati in figura sono quindi, rispettivamente, quelli dell acido (R)-lattico e dell acido (S)-lattico. La scritta acido (RS)-lattico ne indica sinteticamente il racemo. L indicazione (R) o (S) deriva dall applicazione della regola sopra indicata e non è in relazione con il senso della rotazione del piano della luce polarizzata, unica proprietà fisica che distingue i due enantiomeri. Quando il senso della rotazione del piano della luce polarizzata provocato da ciascun enantiomero è noto, questi può essere indicato nel nome del composto con i prefissi (+) (rotazione in senso orario, enantiomero destrogiro) o (—) (rotazione in senso antiorario, enantiomero levogiro). I nomi completi dei due modelli riportati in figura sono quindi, rispettivamente, acido (R)-(—)-lattico e acido (S)-(+)-lattico. Se vi sono due o più atomi di carbonio stereogenici , ad ognuno di essi viene assegnato il prefisso (R) o (S) in base alle regole sopra esposte. Per gli zuccheri ed altre molecole di interesse biochimico, data la complessità di molte di queste, non si fa uso dei prefissi (R) ed (S). La distinzione in nomenclatura viene fatta assegnando i prefissi D ed L a seconda che i sostituenti legati all atomo di carbonio asimmetrico più distante da quello che lega il gruppo carbonilico abbiano disposizione simile a quella della L-gliceraldeide o a quella della D-gliceraldeide.")
12
GLICERALDEIDE L D

13
I gruppi NH2 e COOH possono ionizzarsi in soluzione acquosa
I gruppi NH2 e COOH possono ionizzarsi in soluzione acquosa. L’amminoacido è caricato positivamente (-NH3+) a bassi pH e negativamente (COO-) ad alti pH. Ad un pH intermedio (punto isoelettrico o pHi) l’amminoacido si comporta come uno ione dipolare, con carica risultante nulla.
 a bassi pH e negativamente (COO-) ad alti pH. Ad un pH intermedio (punto isoelettrico o pHi) l’amminoacido si comporta come uno ione dipolare, con carica risultante nulla.")
14
pH

15
Proteine Rappresentano i composti organici più abbondanti nella cellula. Tutte le proteine contengono C (≈50%w), H (≈7%w), O (≈23%w) ed N (≈16%w). Possono contenere anche S (max 3% w). Il peso molecolare varia tra 6,000 ad oltre 1,000,000. Dal punto di vista chimico, una proteina è un polimero (o macromolecola) di residui amminoacidici, uniti mediante un legame peptidico (proteina semplice), spesso in associazione con altre molecole e/o ioni metallici (proteina coniugata). ‘ Per PM inferiore a 10,000 si parla in genere di peptidi o polipeptidi.
. Il peso molecolare varia tra 6,000 ad oltre 1,000,000. Dal punto di vista chimico, una proteina è un polimero (o macromolecola) di residui amminoacidici, uniti mediante un legame peptidico (proteina semplice), spesso in associazione con altre molecole e/o ioni metallici (proteina coniugata). ‘ Per PM inferiore a 10,000 si parla in genere di peptidi o polipeptidi.")
16
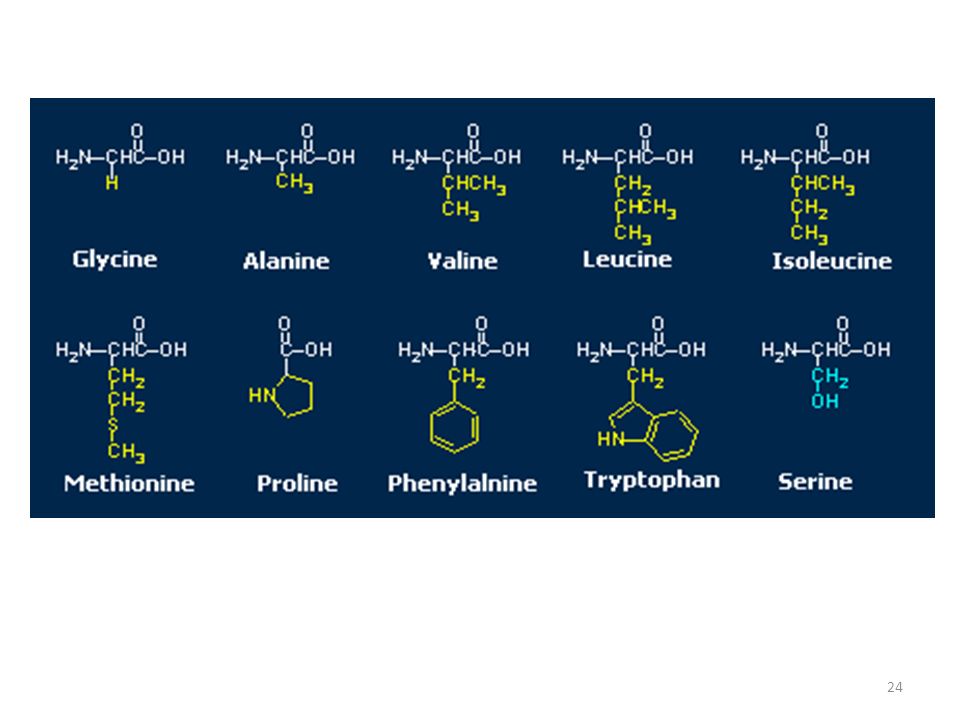
Gli amminoacidi che compaiono nelle proteine di tutti gli organismi viventi sono 20 (studi recenti suggeriscono che questo numero potrebbe aumentare fino a 23) e sono sotto controllo genetico (l'informazione del tipo e della posizione di un amminoacido in una proteina è codificata nel DNA). Talvolta, nelle proteine compaiono anche altri amminoacidi, più rari, detti occasionali. In natura sono stati finora scoperti oltre 500 amminoacidi diversi che non fanno parte di proteine e svolgono ruoli biologici diversi. Alcuni sono stati addirittura trovati nelle meteoriti.

17
Amminoacidi idrofobi Vediamo i 20 amminoacidi essenziali (ordinari):
Glicina (Gly, G) Alanina (Ala, A) Valina (Val, V) Leucina (Leu, L)
 Alanina (Ala, A) Valina (Val, V) Leucina (Leu, L)")
18
Triptofano (Trp, W) Metionina (Met, M) Fenilalanina (Phe, F) Isoleucina (Ile, I)
 Metionina (Met, M) Fenilalanina (Phe, F) Isoleucina (Ile, I)")
19
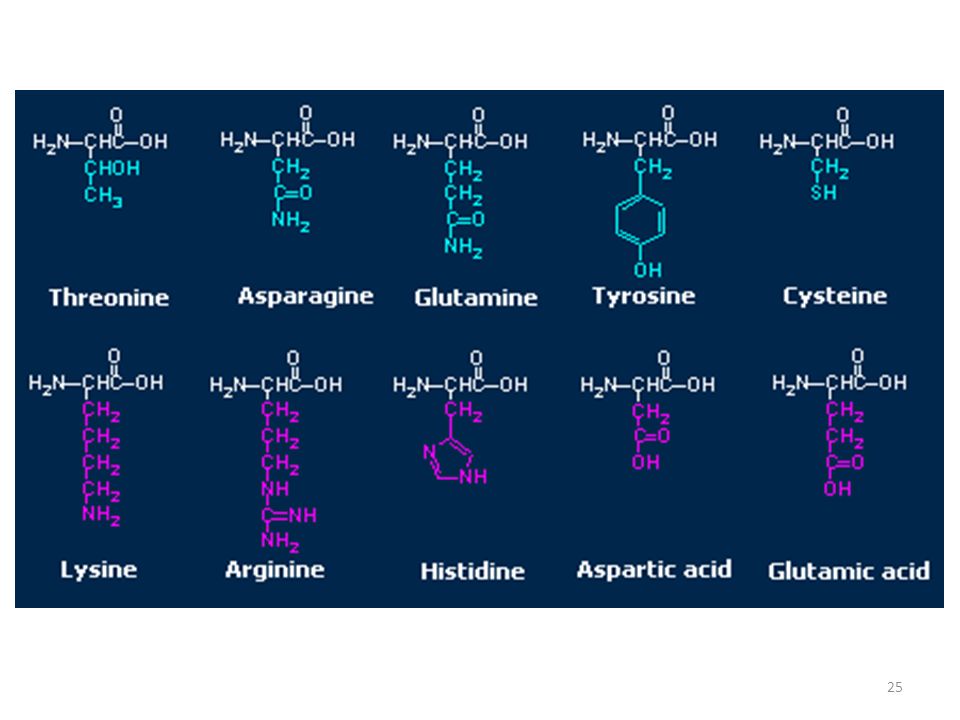
Amminoacidi idrofili idrossilici
Treonina (Thr, T) Serina (Ser, S) Tirosina (Tyr, Y)
 Serina (Ser, S) Tirosina (Tyr, Y)")
20
Amminoacidi idrofili ammidici
Glutammina (Gln, Q) Asparagina (Asn, N)
 Asparagina (Asn, N)")
21
Amminoacidi idrofili acidi
Acido aspartico (Asp, D) Acido glutammico (Glu, E)
 Acido glutammico (Glu, E)")
22
Amminoacidi idrofili basici
Arginina (Arg, R) Lisina (Lys, K) Istidina (His, H)
 Lisina (Lys, K) Istidina (His, H)")
23
Altri amminoacidi Cisteina (Cys, C) Prolina (Pro, P)
 Prolina (Pro, P)")
26
Per capire quante proteine si possono formare con 20 diversi tipi di amminoacidi, basta immaginare quante parole possiamo comporre con le 21 lettere dell'alfabeto, inoltre, proteine che contengono lo stesso tipo e numero di amminoacidi possono differire dall'ordine in cui questi sono situati nella struttura della molecola. Una minima variazione nella sequenza degli amminoacidi di una proteina (cioè nell'ordine con cui i vari tipi di amminoacidi si susseguono) può portare a variazioni nella struttura tridimensionale della macromolecola che possono rendere la proteina non funzionale.
 può portare a variazioni nella struttura tridimensionale della macromolecola che possono rendere la proteina non funzionale.")
28
NEL CSAO DLELE POINERTE QTEUSO NON E’ VREO!

29
Gli amminoacidi entrati a far parte di una catena vengono chiamati residui.
La sequenza di amminoacidi nella catena costituisce la struttura primaria. Dalle interazioni fra i vari gruppi presenti, la proteina assume una particolare struttura (conformazione nativa o struttura secondaria).
.")
30
La struttura secondaria consiste nella conformazione spaziale delle catene e può essere:
A spirale (alfa elica), mantenuta e consentita da legami a idrogeno. Le α eliche si formano quando un certo numero di residui amminoacidici consecutivi adottano coppie di angoli di legame compresi fra -60° e -45°. Lo scheletro del polipeptide risulta arrotolato attorno ad un asse centrale, mentre i gruppi laterali -R dei residui amminoacidici sporgono radialmente all'esterno dell'elica.
, mantenuta e consentita da legami a idrogeno. Le α eliche si formano quando un certo numero di residui amminoacidici consecutivi adottano coppie di angoli di legame compresi fra -60° e -45°. Lo scheletro del polipeptide risulta arrotolato attorno ad un asse centrale, mentre i gruppi laterali -R dei residui amminoacidici sporgono radialmente all esterno dell elica.")
31
Ogni giro completo dell'elica corrisponde ad una distanza di 5,4 Å lungo l'asse, il che implica che vengano coinvolti 3.6 amminoacidi ogni giro. Le α eliche finora trovate sono destrorse e rappresentano circa un quarto della struttura secondaria di tutte le proteine. All'interno dell'α elica ogni legame peptidico partecipa ad un legame idrogeno tra l'idrogeno attaccato all'azoto elettronpositivo e l'ossigeno carbonilico del quarto residuo amminoacidico successivo. Dunque ogni giro d'elica è unito a quelli adiacenti da tre o quattro legami idrogeno, il che rende particolarmente stabile la struttura.

32
Planare (o a foglietto beta)
È una struttura planare. I piani si dispongono, nella struttura β, a zig zag con un angolo di circa 120°. I residui amminoacidici -R sporgono al di sopra e al di sotto del foglietto alternativamente. Anche in questo caso si vengono a formare legami a idrogeno, perpendicolari all'asse, per stabilizzare la struttura, ma questo avviene tra foglietti β diversi, separati anche da molti aminoacidi.

34
A triplice elica (es collagene)
")
35
Le interazioni fra le catene laterali determinano una struttura compatta tridimensionale con ripiegamento dell’intera catena proteica (struttura terziaria). La catena proteica tende a disporsi in modo da orientare i gruppi R polari dei residui verso l’esterno, dove vengono solvatati dall’acqua e i gruppi R idrofobi verso l’interno.

36
La struttura terziaria può essere fibrosa (in genere elevato contenuto di alpha elica, molecole rigide a forma di bastoncello) o globulare (conformazione spaziale molto complessa, compatta, con cavità di dimensioni ridotte e forma ben definita). Le strutture secondaria e terziaria delle proteine possono essere distrutte per azione di vari agenti (denaturazione) sia fisici che chimici, ad esempio opera di acidi, alcooli, calore, solventi e agenti chimici, luce ultravioletta, agitazione meccanica etc. La denaturazione può essere reversibile o irreversibile.
 sia fisici che chimici, ad esempio opera di acidi, alcooli, calore, solventi e agenti chimici, luce ultravioletta, agitazione meccanica etc. La denaturazione può essere reversibile o irreversibile.")
37
Le proteine coniugate sono legate ad un gruppo di natura non proteica (gruppo prostetico).
Nucleoproteine: il gruppo prostetico è un acido nucleico. Glicoproteine: il gruppo prostetico contiene un carboidrato o un suo derivato. Fosfoproteine: il gruppo prostetico contiene fosforo. Lipoproteine: il gruppo prostetico è un lipide.

38
Carboidrati (o glucidi)
Sono composti da C, H ed O con formula Cx(H2O)y. La desinenza generale è –osio ed essendo chimicamente delle aldeidi o dei chetoni, possono essere suddivisi in aldosi e chetosi. aldeide chetone
y. La desinenza generale è –osio ed essendo chimicamente delle aldeidi o dei chetoni, possono essere suddivisi in aldosi e chetosi. aldeide. chetone.")
39
I glucidi sono i più importanti e veloci fornitori di energia biochimica.
Sono le principali molecole di riserva energetica della maggior parte degli esseri viventi (costituiscono anche molti componenti strutturali delle cellule vegetali, che sono formate per il 40% da cellulosa, il composto organico più abbondante della biosfera). I carboidrati si suddividono inoltre in monosaccaridi (o zuccheri semplici), disaccaridi e polisaccaridi.
. I carboidrati si suddividono inoltre in monosaccaridi (o zuccheri semplici), disaccaridi e polisaccaridi.")
40
Monosaccaridi

41
I pentosi, gli esosi e gli eptosi hanno un sufficiente numero di atomi tale da potersi chiudere ad anello per reazione tra il carbonile e l'ossidrile legato al penultimo atomo di carbonio della catena. L'anello può essere costituito da cinque atomi (quattro di carbonio e uno di ossigeno) o da sei atomi (cinque di carbonio e uno di ossigeno) .
 o da sei atomi (cinque di carbonio e uno di ossigeno) .")
42
Ognuna delle due forme esiste in due configurazioni diverse, dette α e β, a seconda che il nuovo gruppo -OH, ottenuto dall'apertura del doppio legame carbonilico, giaccia al di sotto o al di sopra del piano medio della molecola.

43
* * * * * * Interconversione tra le diverse forme del D-glucosio La forma aperta è al centro, a sinistra ci sono le forme cicliche furanosiche, a destra le forme cicliche piranosiche; in alto le forme cicliche α, in basso le β.

44
Disaccaridi Costituiti dall’unione di due monosaccaridi con perdita di una molecola d’acqua. Presente nell’orzo ed alcuni cereali germinati.

45
Il comune zucchero da tavola, presente in barbabietola e canna da zucchero.
Lo zucchero del latte.

46
Polisaccaridi

48
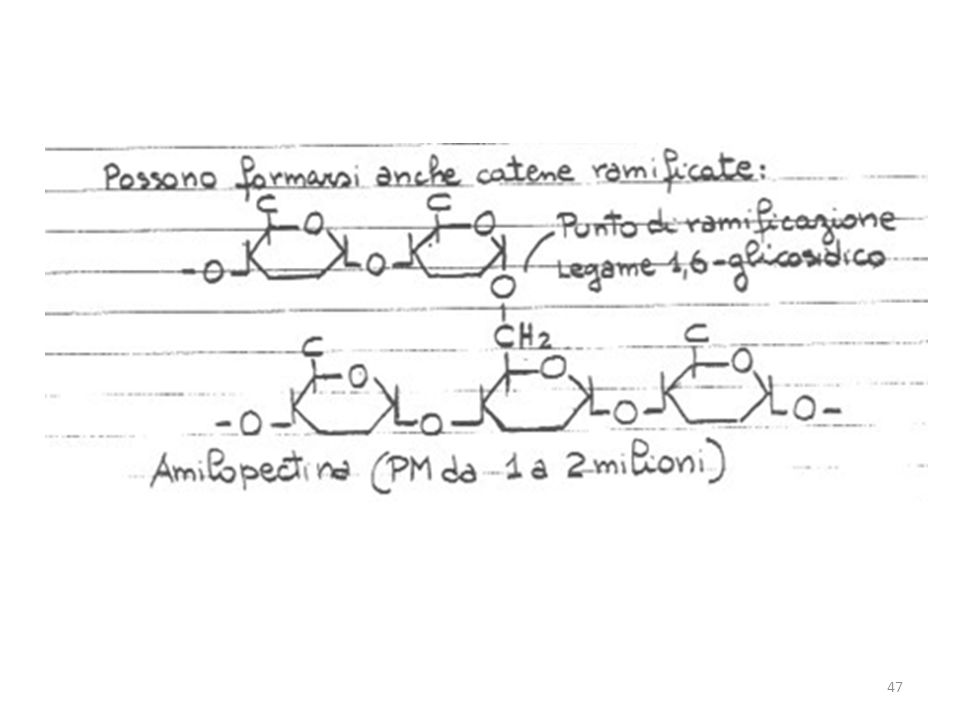
La molecola della cellulosa (componente strutturale principale delle piante) è una lunga catena non ramificata di sub-unità del glucosio con un PM da 50,000 a più di un milione. Sebbene il legame glicosidico nella cellulosa avvenga fra gli atomi di C 1 e 4, le sub-unità sono legate diversamente che nell’amilosio:

49
Lipidi I lipidi (detti anche grassi, dal greco lypos, grasso) sono molecole organiche, raggruppate sulla base delle loro proprietà comuni di solubilità: sono insolubili in acqua (per questo si definiscono idrofobi), mentre sono solubili in solventi organici non polari (nelle molecole lipidiche i gruppi non polari idrofobi sono prevalenti su quelli idrofili). A seconda del grado di complessità, i lipidi si suddividono in 3 categorie: lipidi semplici (trigliceridi, cere, terpeni); lipidi complessi (fosfolipidi); lipidi derivati (steroidi).
 sono molecole organiche, raggruppate sulla base delle loro proprietà comuni di solubilità: sono insolubili in acqua (per questo si definiscono idrofobi), mentre sono solubili in solventi organici non polari (nelle molecole lipidiche i gruppi non polari idrofobi sono prevalenti su quelli idrofili). A seconda del grado di complessità, i lipidi si suddividono in 3 categorie: lipidi semplici (trigliceridi, cere, terpeni); lipidi complessi (fosfolipidi); lipidi derivati (steroidi).")
50
Gliceridi e trigliceridi
Lipidi semplici Gliceridi e trigliceridi I gliceridi sono esteri del glicerolo con acidi grassi. glicerolo (1,2,3 propantriolo) L'acido grasso è formato da un gruppo carbossilico e da una catena idrocarburica.
 L acido grasso è formato da un gruppo carbossilico e da una catena idrocarburica.")
51
Un trigliceride è un lipide costituito da una molecola di glicerolo a cui sono legati 3 acidi grassi.

52
I triglicèridi sono i lipidi più abbondanti di origine naturale, e costituiscono i grassi animali e gli oli vegetali. Servono soprattutto come deposito per l'energia prodotta e immagazzinata a livello di tessuto adiposo. Spesso i 3 acidi grassi sono differenti. Gli acidi grassi sono i lipidi più semplici e comuni, e possono differire per la lunghezza e/o il tipo di legame tra gli atomi di carbonio, legami che possono essere tutti singoli, e allora si parla di acidi grassi saturi, oppure no, e in questo caso si parla di acidi grassi insaturi (monoinsaturi se c'è un solo doppio legame, polinsaturi altrimenti).
.")
53
Sono stati isolati, da varie cellule e tessuti, più di 500 tipi di acidi grassi, e si può notare che quasi sempre questi hanno un numero pari di atomi di carbonio, solitamente compreso tra 12 e 20. Le insaturazioni impediscono alle molecole di compattarsi saldamente e di solidificare a temperatura ambiente. La maggior parte dei grassi vegetali è composta da oli insaturi, mentre i grassi animali si dividono: nei pesci prevalgono i grassi insaturi, negli animali terrestri quelli saturi (burro). La margarina è solida perché i grassi sono parzialmente saturati tramite processo di idrogenazione.
. La margarina è solida perché i grassi sono parzialmente saturati tramite processo di idrogenazione.")
54
Cere Le cere (in biochimica) sono esteri di alcoli superiori monovalenti (cioè a lunga catena e con una sola funzione alcolica) con acidi grassi anch’essi a lunga catena (C30-C40).
 sono esteri di alcoli superiori monovalenti (cioè a lunga catena e con una sola funzione alcolica) con acidi grassi anch’essi a lunga catena (C30-C40).")
55
Terpeni I terpeni sono costituiti da multipli dell'unità isoprenica (isoprenoidi). La loro struttura contiene cioè uno scheletro idrocarburico rispondente alla formula (C5H10)n. sulla catena si trovano poi funzioni ossigenate come –OH, -CHO etc).
n. sulla catena si trovano poi funzioni ossigenate come –OH, -CHO etc).")
56
trans cis

57
Appartengono ai terpeni alcune vitamine (A, E, etc).
Mircene (uno dei composti aromatici principali del luppolo) Lo squalene è un terpene presente nell'olio di fegato degli squali (elasmobranchi) ma anche nei vegetali come i semi di amaranto, crusca di riso, germe di grano e olive. Tutti gli organismi superiori producono squalene, inclusi gli esseri umani. Lo squalene è il precursore biochimico degli steroidi. Il farnesolo è un terpene con un gruppo alcolico che ha la fragranza di mughetto e spesso viene usato nelle profumazioni con questa nota. È un liquido incolore e insolubile in acqua, presente in molti olii essenziali (es. citronella, neroli, cyclamen, rosa, muschio ecc.). Il farnesolo rappresenta anche un pesticida naturale contro gli acari e un feromone per diversi altri tipi di insetti.
 Lo squalene è un terpene presente nell olio di fegato degli squali (elasmobranchi) ma anche nei vegetali come i semi di amaranto, crusca di riso, germe di grano e olive. Tutti gli organismi superiori producono squalene, inclusi gli esseri umani. Lo squalene è il precursore biochimico degli steroidi. Il farnesolo è un terpene con un gruppo alcolico che ha la fragranza di mughetto e spesso viene usato nelle profumazioni con questa nota. È un liquido incolore e insolubile in acqua, presente in molti olii essenziali (es. citronella, neroli, cyclamen, rosa, muschio ecc.). Il farnesolo rappresenta anche un pesticida naturale contro gli acari e un feromone per diversi altri tipi di insetti.")
58
Lipidi complessi Fosfolipidi Sono simili ai triglicèridi dal punto di vista strutturale, ma contengono un gruppo fosfato. Il risultato finale è che ogni fosfolipide ha una testa idrofila e una coda idrofoba. Il gruppo X può essere uno zucchero

59
Questa particolare struttura li rende idonei a formare le membrane biologiche che avvolgono le cellule e gli organuli cellulari. In un ambiente liquido le molecole di fosfolipidi si dispongono con i gruppi idrofili rivolti sia verso la soluzione acquosa interna alle cellule, sia verso quella esterna, relativa all’ambiente circostante. Invece le code idrofobe si attraggono tra loro occupando una posizione mediana. I fosfolipidi sono i principali componenti della frazione lipidica delle membrane cellulari.

60
Lipidi derivati steroidi Sono lipidi policiclici derivati del ciclopentano peridrofenantrene. I loro scheletri carboniosi sono piegati per formare quattro anelli uniti tra loro. Sono sempre costituiti da tre anelli a sei lati e un anello a cinque lati. Comprendono il colesterolo, calciferolo (vitamina D2), testosterone, progesterone, cortisone.
, testosterone, progesterone, cortisone.")
61
Esempio: Colesterolo

62
Nucleotidi I nucleotidi sono degli esteri fosforici dei nucleosidi, costituiti da tre sub-unità: Una base azotata (purina o pirimidina); uno zucchero a cinque atomi di carbonio (pentosio) (base + zucchero costituiscono un nucleoside); un gruppo fosfato.
 (base + zucchero costituiscono un nucleoside); un gruppo fosfato.")
63
I nucleotidi sono i monomeri degli acidi nucleici (DNA e RNA).
La presenza del residuo fosforico conferisce carattere fortemente acido ai nucleotidi (per questo noti anche come acido adenilico o acido guanilico). L'aggiunta di uno o di due altri residui fosforici nella catena (fosforilazione ossidativa) produce i nucleosidi di- e trifosfato (NDP e NTP), fondamentali nel metabolismo energetico della cellula.
. L aggiunta di uno o di due altri residui fosforici nella catena (fosforilazione ossidativa) produce i nucleosidi di- e trifosfato (NDP e NTP), fondamentali nel metabolismo energetico della cellula.")
64
basi Le basi puriniche adenina e guanina si ritrovano sia nel DNA sia nell’RNA.

65
Le basi pirimidiniche Citosina e Timina si ritrovano nel DNA, mentre nell'RNA troviamo citosina e Uracile.

66
pentosi Il DNA contiene il D-2-deossiribosio, l’RNA contiene il D-ribosio.

67
I nomi dei nucleotidi sono abbreviati in codici standard di tre o quattro lettere.
Una d iniziale sta per "deossi-", cioè indica che il nucleotide in questione è un deossiribonucleotide. Nel caso di un ribonucleotide semplicemente non c'è la d. La seconda lettera indica il nucleoside corrispondente alla nucleobase (G, A, T, C, U). La terza e la quarta lettera indicano la lunghezza della catena di gruppi fosfato attaccata (mono-, di- e tri-, abbreviati in M, D,T) e la presenza del gruppo fosfato (P).
. La terza e la quarta lettera indicano la lunghezza della catena di gruppi fosfato attaccata (mono-, di- e tri-, abbreviati in M, D,T) e la presenza del gruppo fosfato (P).")
68
Ad esempio, la deossicitosina trifosfato è abbreviata con dCTP (la d perché è un deossiribonucleotide, la C perché è una citosina, TP perché è un tri-fosfato).
.")
70
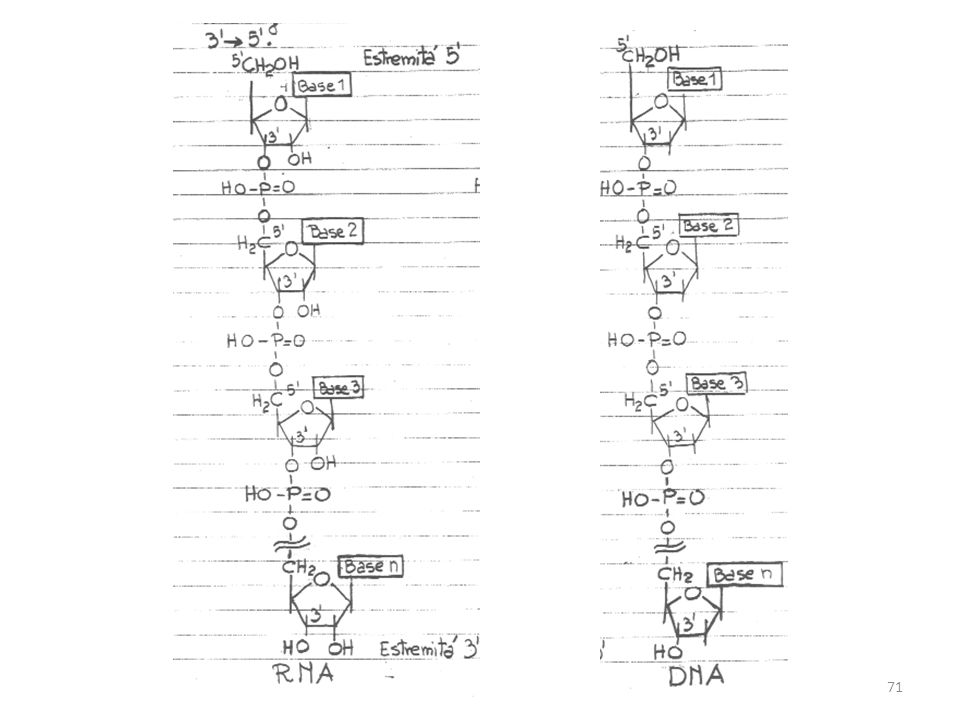
polinucleotidi Due sono i polinucleotidi importanti: gli acidi ribonucleici (RNA) e gli acidi deossiribonucleici (DNA). In entrambi, le unità mononucleotidiche sono legate allo stesso modo da ponti fosfo-diesterei 3’-5’. Dal punto di vista chimico, DNA ed RNA sono polimeri organici le cui unità ripetitive sono nucleotidi.
 e gli acidi deossiribonucleici (DNA). In entrambi, le unità mononucleotidiche sono legate allo stesso modo da ponti fosfo-diesterei 3’-5’. Dal punto di vista chimico, DNA ed RNA sono polimeri organici le cui unità ripetitive sono nucleotidi.")
72
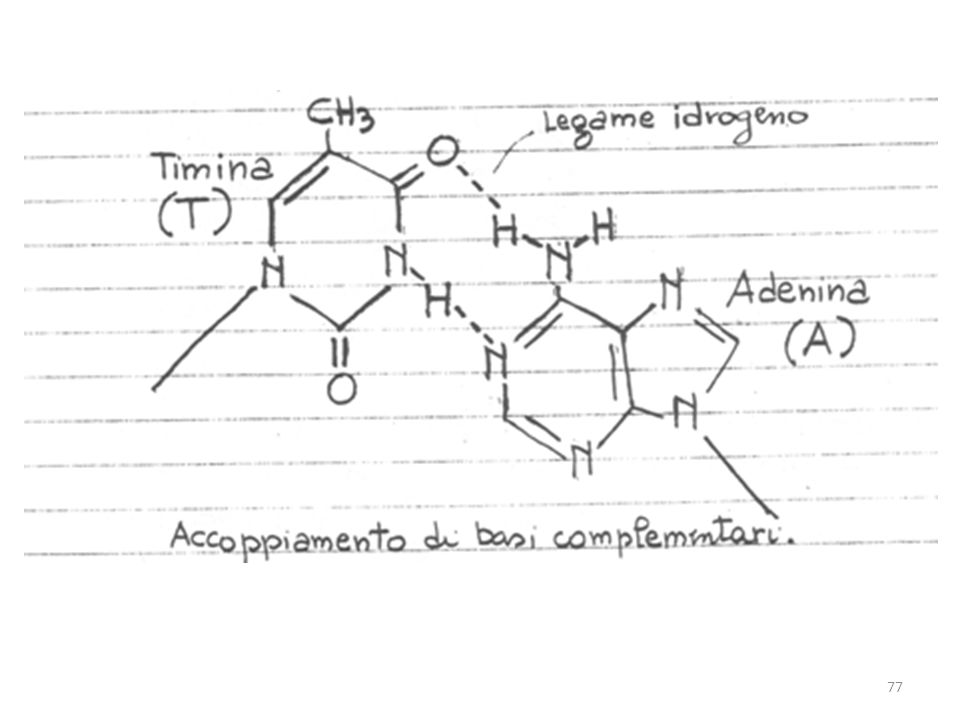
Il DNA contiene le basi azotate A, C, G e T ed è il portatore dell’informazione genetica.
Il DNA di quasi tutti gli esseri viventi, ad eccezione di alcuni virus, è a doppia elica, cioè costituito da 2 catene elicoidali (cromosomi) avvolte l’una sull’altra intorno ad uno stesso asse. Le due catene hanno polarità opposte: esse vanno dall’estremità 5’ a quella 3’ in direzioni opposte.
 avvolte l’una sull’altra intorno ad uno stesso asse. Le due catene hanno polarità opposte: esse vanno dall’estremità 5’ a quella 3’ in direzioni opposte.")
73
Watson e Crick

75
Nel gennaio 1951, Franklin iniziò a lavorare come ricercatrice associata al King's College di Londra e nell'Unità di Biofisica del Medical Research Council (MRC), diretta da John Randall. Anche se in origine avrebbe dovuto lavorare sulla diffrazione a raggi x di proteine in soluzione, il suo lavoro fu cambiato nell'analisi di fibre di DNA prima ancora che iniziasse a lavorare al King's College. Maurice Wilkins e Raymond Gosling avevano analizzato il DNA con il metodo della diffrazione a raggi x dal 1950. Franklin, lavorando con il suo studente Raymond Gosling, iniziò ad applicare le sue competenze a proposito delle tecniche di diffrazione a raggi x per la struttura del DNA. Scoprirono che vi sono due forme di DNA: ad alta umidità (quando bagnato), il DNA è diventa una fibra lunga e sottile, quando si è asciutto, è corto e grosso. Questi DNA sono stati definiti rispettivamente 'B' e 'A'. Il lavoro sul DNA fu successivamente diviso tra Franklin e Wilkins: la prima analizzò la forma 'A', mentre il secondo studiò la forma 'B'. Le foto scattate da Franklin con la diffrazione a raggi x furono definite da John Desmond Bernal "tra le più belle fotografie a raggi x di qualsiasi sostanza mai scattate". Entro la fine del 1951 era stata generalmente accettata al King's College che la forma 'B' del DNA era una spirale, ma si dubitava fortemente che la forma 'A' del DNA avesse una struttura ad elica. Nel corso del 1952 Rosalind Franklin e Raymond Gosling applicarono la funzione di Patterson alle foto a raggi x di DNA che avevano realizzato: fu un lavoro lungo e difficile, ma diede una panoramica della struttura delle molecole. Tra le altre immagini ottenute da Rosalind Franklin, ci fu la Photograph 51, che rappresentava il DNA nella sua forma B. Su questo fatto, sull'analisi del riscoperto epistolario della scienziata e sulla base di interviste ai protagonisti della storia della scoperta della struttura del DNA, si è venuta a formare l'ipotesi che fu proprio Franklin la vera scopritrice della morfologia ad elica del DNA. Nel febbraio 1953 Francis Crick e James Watson del Cavendish Laboratory dell'Università di Cambridge avevano iniziato a costruire un modello della forma 'B' del DNA utilizzando dati simili a quelli disponibili al team del King's College. La costruzione di un modello era stata utilizzata con successo nella spiegazione della struttura dell'alfa elica da Linus Pauling nel 1951, ma Rosalind Franklin si opponeva alla costruzione di modelli teorici, ritenendo che la costruzione di un modello doveva essere intrapresa soltanto dopo che la struttura fosse stata individuata esattamente. Maurice Wilkins mostrò a Watson all'insaputa della ricercatrice la foto 51 e questo avvenimento fornì lo spunto fondamentale a Francis Crick ed a James Watson per elaborare quello che venne chiamato “Il modello della doppia elica”. Si ottenne un modello coerente con le conoscenze teoriche e sperimentali dell'epoca, risultato, in ogni caso, di speculazioni teoriche sul lavoro sperimentale svolto soprattutto nel laboratorio di Wilkins e della sua collaboratrice. L'articolo venne pubblicato il 25 aprile del 1953, sulla rivista Nature. Sulla stessa rivista furono pubblicati due lavori, uno di Rosalind Franklin e uno di Maurice Wilkins. Nell'autobiografia di Watson “La doppia elica”, pubblicata nel 1968 viene descritta come “la terribile Rosy”, una donna non attraente, e dal carattere pessimo, molto gelosa del proprio lavoro. Questo fatto evidenzia l'esistenza di screzi tra i due scienziati. Nel 2003 la rivista on-line BioMedNet News ha adombrato il fatto che fu Raymond Gosling, allora suo studente di dottorato al King’s College di Londra e non la Franklin il vero autore della foto 51. Non ottenne il Nobel anche perché morì prematuramente, a soli 37 anni, a causa di un tumore probabilmente generato dalla forte esposizione ai raggi X, ma è tuttavia opinione condivisa che il suo contributo in tali ricerche fu sostanziale. Rosalind Franklin
, il DNA è diventa una fibra lunga e sottile, quando si è asciutto, è corto e grosso. Questi DNA sono stati definiti rispettivamente B e A . Il lavoro sul DNA fu successivamente diviso tra Franklin e Wilkins: la prima analizzò la forma A , mentre il secondo studiò la forma B . Le foto scattate da Franklin con la diffrazione a raggi x furono definite da John Desmond Bernal tra le più belle fotografie a raggi x di qualsiasi sostanza mai scattate . Entro la fine del 1951 era stata generalmente accettata al King s College che la forma B del DNA era una spirale, ma si dubitava fortemente che la forma A del DNA avesse una struttura ad elica. Nel corso del 1952 Rosalind Franklin e Raymond Gosling applicarono la funzione di Patterson alle foto a raggi x di DNA che avevano realizzato: fu un lavoro lungo e difficile, ma diede una panoramica della struttura delle molecole. Tra le altre immagini ottenute da Rosalind Franklin, ci fu la Photograph 51, che rappresentava il DNA nella sua forma B. Su questo fatto, sull analisi del riscoperto epistolario della scienziata e sulla base di interviste ai protagonisti della storia della scoperta della struttura del DNA, si è venuta a formare l ipotesi che fu proprio Franklin la vera scopritrice della morfologia ad elica del DNA. Nel febbraio 1953 Francis Crick e James Watson del Cavendish Laboratory dell Università di Cambridge avevano iniziato a costruire un modello della forma B del DNA utilizzando dati simili a quelli disponibili al team del King s College. La costruzione di un modello era stata utilizzata con successo nella spiegazione della struttura dell alfa elica da Linus Pauling nel 1951, ma Rosalind Franklin si opponeva alla costruzione di modelli teorici, ritenendo che la costruzione di un modello doveva essere intrapresa soltanto dopo che la struttura fosse stata individuata esattamente. Maurice Wilkins mostrò a Watson all insaputa della ricercatrice la foto 51 e questo avvenimento fornì lo spunto fondamentale a Francis Crick ed a James Watson per elaborare quello che venne chiamato Il modello della doppia elica . Si ottenne un modello coerente con le conoscenze teoriche e sperimentali dell epoca, risultato, in ogni caso, di speculazioni teoriche sul lavoro sperimentale svolto soprattutto nel laboratorio di Wilkins e della sua collaboratrice. L articolo venne pubblicato il 25 aprile del 1953, sulla rivista Nature. Sulla stessa rivista furono pubblicati due lavori, uno di Rosalind Franklin e uno di Maurice Wilkins. Nell autobiografia di Watson La doppia elica , pubblicata nel 1968 viene descritta come la terribile Rosy , una donna non attraente, e dal carattere pessimo, molto gelosa del proprio lavoro. Questo fatto evidenzia l esistenza di screzi tra i due scienziati. Nel 2003 la rivista on-line BioMedNet News ha adombrato il fatto che fu Raymond Gosling, allora suo studente di dottorato al King’s College di Londra e non la Franklin il vero autore della foto 51. Non ottenne il Nobel anche perché morì prematuramente, a soli 37 anni, a causa di un tumore probabilmente generato dalla forte esposizione ai raggi X, ma è tuttavia opinione condivisa che il suo contributo in tali ricerche fu sostanziale. Rosalind Franklin.")
76
Le due ossature zucchero-fosfato, fortemente idrofile, si dispongono all’esterno della struttura e le coppie di basi idrofobe sono impilate all’interno e disposte perpendicolarmente rispetto all’asse longitudinale. Le basi di un filamento sono appaiate alle basi dell’altro in maniera selettiva attraverso legami a idrogeno. La doppia elica consente il massimo numero possibile di legami ad idrogeno, rendendo la struttura molto stabile.

78
La sequenza delle basi contiene l’informazione genetica, leggibile attraverso il codice genetico che ne permette la traduzione in amminoacidi. Ogni tipo di base presente su un filamento forma un legame con la base posta sul filamento opposto (appaiamento complementare). Le basi puriniche formano legami idrogeno con le basi pirimidiniche: A può legare solo T G può legare solo C.
. Le basi puriniche formano legami idrogeno con le basi pirimidiniche: A può legare solo T. G può legare solo C.")
81
Replicazione del DNA Il DNA è in grado di autoreplicarsi se si trova nelle adatte condizioni: la doppia elica si svolge e ciascun filamento funziona da stampo per la sintesi di un filamento a lui complementare.

82
In un eucariote la replicazione del DNA inizia contemporaneamente in migliaia di punti di origine:

83
Nei batteri (e in molti virus) il DNA è circolare e pare che come tale venga replicato. Due forcelle di replicazione prendono origine nello stesso punto e si allontanano da esso in entrambe le direzioni contemporaneamente, fino a quando si incontrano.

Presentazioni simili








>")
















>")
