Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
INGEGNERIA PROTEICA

2
L’ingegneria proteica è stata usata per due scopi:
La dissezione della struttura e dell’attività di una proteina esistente, attraverso alterazioni sistematiche e l’esame dei cambiamenti delle proprietà. (esempio: tirosil-tRNA sintetasi) 2) La produzione di “nuove” proteine per la medicina e per l’industria. (esempio: subtilisina)
 2) La produzione di nuove proteine per la medicina e. per l’industria. (esempio: subtilisina)")
3
Per comprendere il meccanismo catalitico di un enzima
non basta identificare tutti gli intermedi di reazione, ma bisogna conoscere la natura e l’energia delle interazioni tra proteina e substrati, intermedi, stati di transizione, prodotti. L’energia di queste interazioni è infatti usata per abbassare l’energia di attivazione e per determinare la specificità. L’ingegneria proteica è uno strumento di indagine eccellente, in quanto può essere usata per alterare sistematicamente le interazioni tra proteina e substrato e le interazioni all’interno della proteina, al fine di analizzarne le proprietà allosteriche e strutturali.

4
LA TIROSIL-tRNA SINTETASI
Questo fu il primo enzima ad essere studiato, nel 1982, tramite ingegneria proteica. Allora, il suo meccanismo catalitico era del tutto sconosciuto, nonostante fosse nota la struttura cristallografica. La tirosil-tRNA sintetasi di Bacillus stearotermophilus è un omodimero e catalizza l’aminoacilazione del tRNATyr in due passaggi: E + Tyr + ATP E•Tyr-AMP + PPi E•Tyr-AMP + tRNA Tyr-tRNA + E + AMP Nel primo la tirosina viene attivata formando un complesso enzima- tirosiladenilato molto stabile. Nel secondo la Tyr viene trasferita al tRNATyr.

5
Il meccanismo della reazione di attivazione, consiste in un attacco
nucleofilo del carbossilato della Tyr sul fosfato dell’ATP che genera un intermedio pentacovalente. Quest’ultimo elimina il pirofosfato (come sale del Mg2+). Il fosforo inverte la sua configurazione. E’ stata risolta la struttura cistallografica dei complessi E•Tyr e E•Tyr-AMP.
. Il fosforo inverte la sua. configurazione. E’ stata risolta la struttura cistallografica dei complessi E•Tyr e. E•Tyr-AMP.")
6
I REQUISITI CHE DEVONO ESSERE SODDISFATTI
PER CONDURRE DEGLI STUDI SISTEMATICI DI MUTAGENESI SITO-SPECIFICA SONO: La proteina deve essere disponibile in forma ricombinante La struttura cristallografica deve essere stata risolta con una risoluzione elevata. I valori assoluti delle costanti di velocità devono essere noti. Preparazioni diverse di enzima danno valori diversi, per via della presenza di quantità variabili di enzima inattivo. E’ necessario quindi normalizzare i valori ottenuti tramite titolazione dei siti attivi. Gli studi allo stato pre-stazionario non sono soggetti a queste variazioni e non soffrono della contaminazione di piccole quantità di enzima selvatico.

7
Regole per produrre mutanti che possano dare risultati
interpretabili: Le mutazioni devono essere isosteriche o comportare una delezione; Evitare di creare cariche interne non bilanciate; Eliminare il minor numero di interazioni; Non creare nuove interazioni che causino una riorganizzazione locale della struttura; 5) tutte queste regole possono essere infrante se necessario.
 tutte queste regole possono essere infrante se necessario.")
8
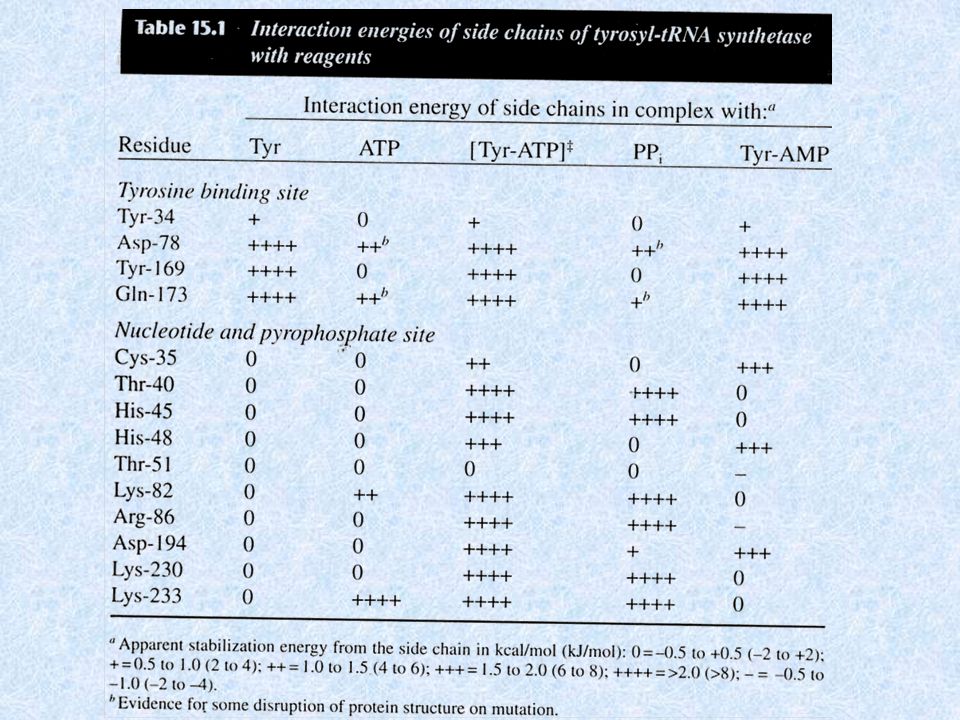
Residui della tirosil-tRNA sintetasi che formano legami idrogeno
con il tirosil adenilato.

9
Strategia: profili dell’energia libera e diagrammi di energia differenziali
La misura del profilo di energia libera (in realtà delle differenze di energia libera) della reazione catalizzata dalle forme selvatica e mutanti dell’enzima comporta la misura delle seguenti costanti di equilibrio e di velocità: Quindi i profili vengono calcolati utilizzando le teorie della termodinamica e dello stato di transizione. Dalla differenza dei profili dell’enzima selvatico e delle forme mutanti si ottengono dei diagrammi di energia differenziali:
 della reazione catalizzata dalle forme selvatica e. mutanti dell’enzima comporta la misura delle seguenti costanti. di equilibrio e di velocità: Quindi i profili vengono calcolati utilizzando le teorie della. termodinamica e dello stato di transizione. Dalla differenza dei. profili dell’enzima selvatico e delle forme mutanti si ottengono. dei diagrammi di energia differenziali:")
10
Diagramma di energia differenziale
Energia di legame apparente del gruppo eliminato dalla mutazione

11
Attivazione della tirosina:
Dimostrazione della complementarietà tra enzima e stato di transizione Thr40

12
His45

13
Thr40 e His45 formano un sito di legame per il fosfato
dell’ATP nello stato di transizione:

14
Attivazione della tirosina:
2) Scoperta della complementarietà tra enzima ed intermedi: bilanciamento delle costanti di equilibrio interne; sequestro degli intermedi instabili. Cys35 His48
 Scoperta della complementarietà tra enzima ed intermedi: bilanciamento delle costanti di equilibrio interne; sequestro degli intermedi instabili. Cys35. His48.")
15
Ci sono due buone ragioni per la complementarietà tra
enzima ed intermedi di reazione: La complementarietà cambia la costante di equilibrio di reazioni sfavorevoli. Tyr + ATP Tyr-AMP + PPi da 3,5•10-7 a 2,3 il tirosiladenilato deve essere presente ad elevate concentrazioni nell’enzima perché deve reagire con il tRNA. 2) La resa delle reazione viene ad essere aumentata perché il sequestro degli intermedi molto reattivi minimizza le reazioni collaterali. Il tirosiladenilato è molto reattivo ed idrolizza rapidamente.
 La resa delle reazione viene ad essere aumentata perché. il sequestro degli intermedi molto reattivi minimizza le. reazioni collaterali. Il tirosiladenilato è molto reattivo. ed idrolizza rapidamente.")
16
Attivazione della tirosina:
3) Rilevamento del processo di adattamento indotto. Residui presenti sul loop “KMSK” che legano lo stato di transizione
 Rilevamento del processo di adattamento indotto. Residui presenti sul. loop KMSK che. legano lo stato di. transizione.")
17
La flessibilità e l’adattamento indotto
rappresentano un compromesso tra complementarietà tra enzima e stato di transizione e libero accesso dei substrati al sito attivo.

18
IL MECCANISMO CATALITICO DELL’ATTIVAZIONE
DELLA TIROSINA Dai risultati dei diagrammi di energia differenziale si evince che la catalisi non è realizzata da alcun residuo in particolare ma dal numeroso insieme di residui che costituiscono il sito attivo. Essa non viene attuata tramite un classico meccanismo di catalisi acido-base o nucleofila, ma è il risultato della sola energia di legame.

20
I

Presentazioni simili













![AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.](/2/576433/big_thumb.jpg "AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.>")
![AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.](/2/576500/big_thumb.jpg "AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.>")
![AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.](/2/576501/big_thumb.jpg "AA + bB pP + qQ V = -1/a x d[A]/dt = -1/b x d[B]/dt =1/p x d[P]/dt =1/q x d[Q]/dt DEFINIZIONE di velocità di reazione.>")
>")


