Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
GLI OSTEITTI

2
Osteitti:aspetti generali
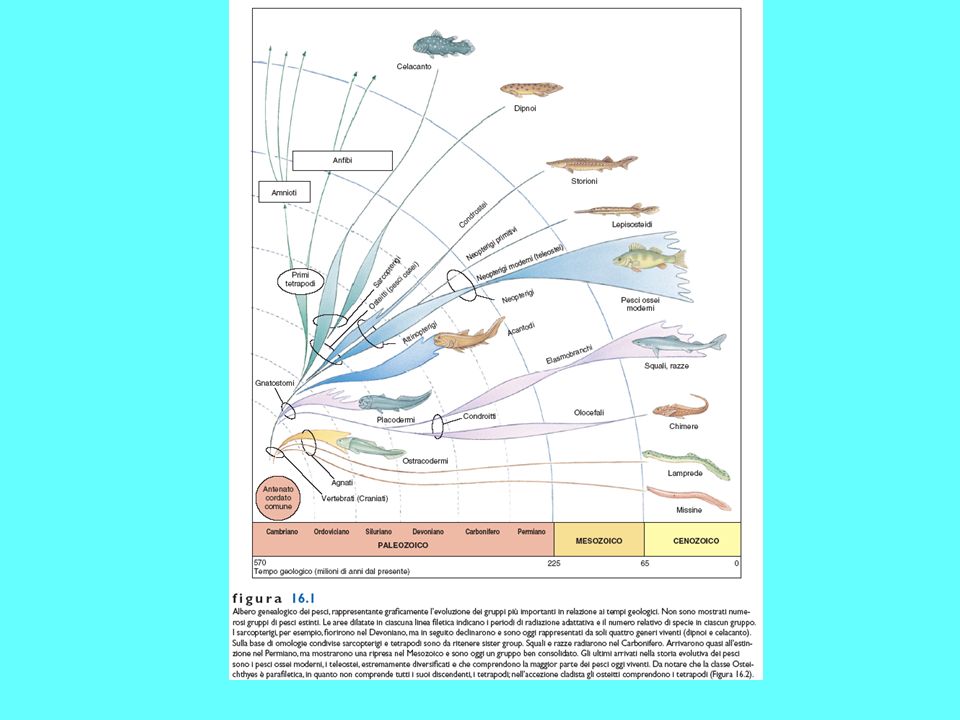
Gli osteitti o pesci ossei comparvero nel tardo Ordoviciano, inizio Siluriano probabilmente da pesci acantodi, pesci di piccole dimensioni presenti nel Siluriano e poi molto comuni per tutto il Paleozoico Gli osteitti, una volta comparsi, si diversificarono nei due taxa degli Attinotterigi e dei Sarcopterigi, un tempo indicati anche come Coanoitti. Fra gli attinotterigi (pesci a pinne raggiate), il maggior successo è arriso ai teleostei, che apparsi nel Triassico, sono oggi i vertebrati con il maggior numero di specie. Il loro successo evolutivo è dovuto alla grande prolificità, alla maggiore efficienza nel nuoto e allo sviluppo di un cranio cinetico che favorisce l’acquisizione di prede anche di grossa mole
, il maggior successo è arriso ai teleostei, che apparsi nel Triassico, sono oggi i vertebrati con il maggior numero di specie. Il loro successo evolutivo è dovuto alla grande prolificità, alla maggiore efficienza nel nuoto e allo sviluppo di un cranio cinetico che favorisce l’acquisizione di prede anche di grossa mole.")
6
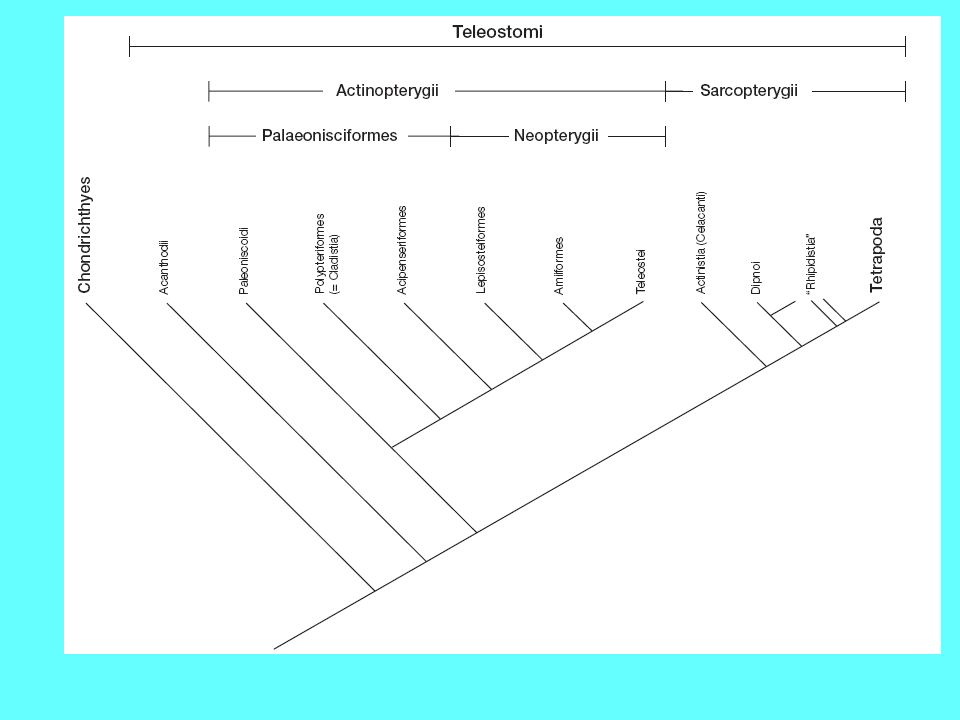
FIGURA 3.17 Filogenesi degli attinopterigi. Secondo J.Z.Young.

7
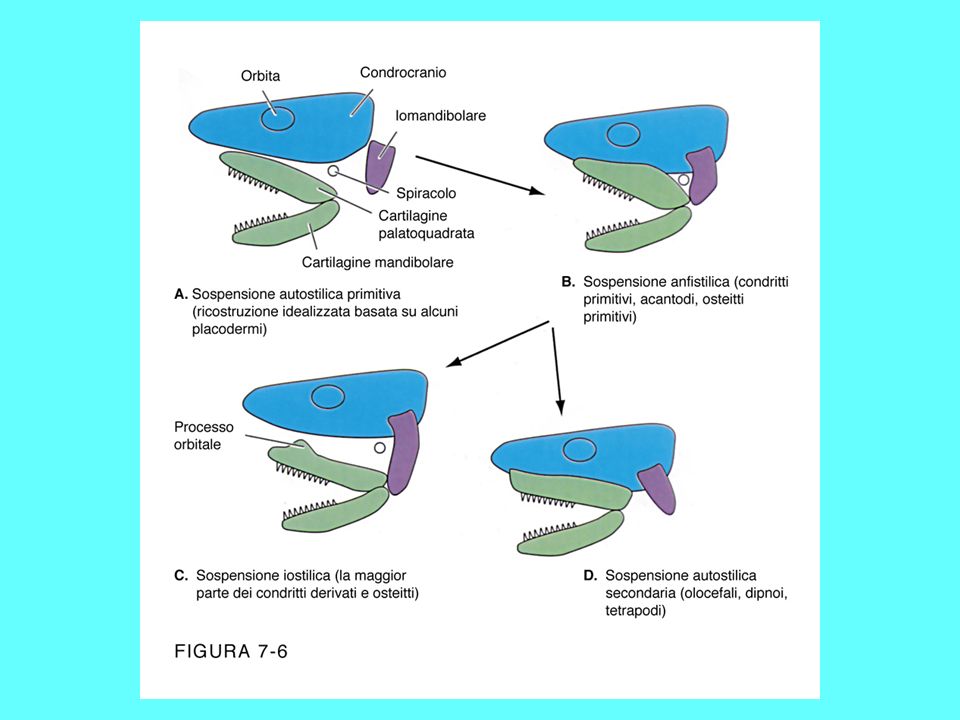
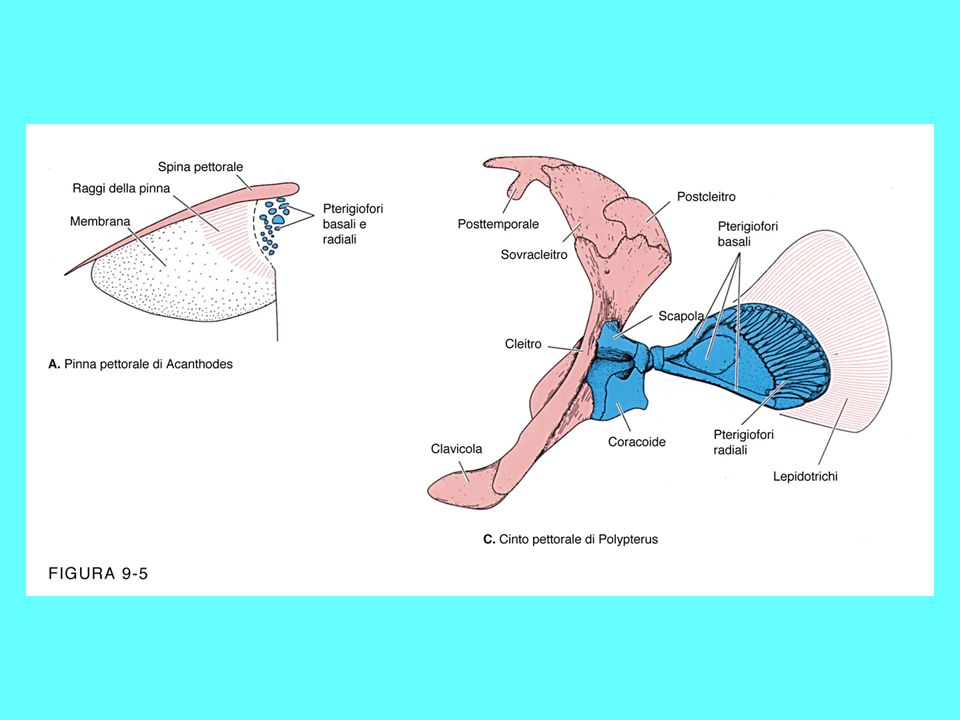
Attinopterigi più antichi
Gli attinopeterigi più antichi sono caratterizzati da una forma del corpo che ricorda quella dei condroitti, per la presenza di una bocca ventrale, uno spiracolo, la coda eterocerca ed un dermascheletro ancora ben strutturato Alcuni attinopeterigi, come lo storione, presentano, nella fase adulta, la corda dorsale, per cui le vertebre risultano prive di centro e costituite da numerosi pezzi che si dispongono introno alla corda stessa

8
Classificazione degli Osteitti

10
Lo storione e il pesce spatola: due Condrostei
Acipenser Polyodon

11
Amia e Lepidosteus: Olostei

13
Caratteri dei Neopterigi
Forma del corpo compressa in senso destra-sisistra Bocca terminale Scaglie cicloidi e/o ctenoidi Coda omocerca Appendici pari con stretta base di impianto e ravvicinate Fessure branchiali che si aprono nella camera branchiale, sormontata dall’opercolo Presenza di vescica natatoria e/o di polmoni Il tetto ottico mesencefalico come centro di associazione per eccellenza Ovipari

17
Correlare la velocità dei cambiamenti morfologici con la velocità dell’origine di nuove specie. Questo l’ambizioso obiettivo di una collaborazione multidisciplinare, finanziata dalla National Science Foundation e che ha coinvolto, oltre a diverse università americane – Berkeley, UCLA, Michigan, Idaho, Oregon State – anche quella di Torino. A questo scopo, i ricercatori hanno raccolto i dati esistenti da diverse fonti come GenBank e FishBase per creare una sorta di “albero della vita” di più di 7000 specie di Attinopterigi, che comprende la maggior parte dei pesci ossei viventi ed è caratterizzata dal possedere pinne sostenute da raggi. Hanno così ottenuto uno dei più grandi alberi filogenetici mai costruiti per uno specifico gruppo di animali, col quale hanno testato l’effettiva esistenza di una correlazione positiva fra tasso di speciazione e tasso di cambiamento morfologico, correlazione ipotizzata da diverse teorie, ma della quale erano state trovate ben poche prove in natura. I risultati del lavoro, pubblicati a giugno su Nature Communications, dimostrano che questa correlazione positiva in effetti esiste: gruppi di pesci che si separano rapidamente in nuove specie tendono a evolvere in fretta una certa varietà di tratti fisici, mentre in quelli che Darwin aveva descritto come “fossili viventi”, concetto che oggi sappiamo non essere valido, sia il numero di specie sia quello delle differenti morfologie è minore. Uno studio importante, poiché simili collegamenti fra speciazione e adattamento morfologico non erano mai stati osservati su una scala così grande. Benché focalizzato sugli Attinopterigi, l’approccio usato dagli autori è potenzialmente applicabile anche ad altri gruppi, dai mammiferi agli uccelli, dagli insetti alle piante.

18
Lo spinarello : un esempio di evoluzione per selezione naturale, sia in natura che in contesti sperimentali. Gli spinarelli (Gasterosteus aculeatus) sonopiccoli pesci distribuiti in quasi tutto l'emisfero settentrionale, dagli oceani fino ai torrenti. Gli spinarelli di mare si distinguono dagli spinarelli di acqua dolce per il numero di placche ossee che ricoprono il loro corpo e che fungono da corazza protettiva. Gli spinarelli di mare esibiscono una corazza composta da un maggior numero di placche, circa 30 dalla testa alla coda, mentre in quelli di acqua dolce si riscontra una corazza molto ridotta, a volte quasi assente. La diversificazione delle due forme ha avuto inizio alla fine dell'ultima era glaciale del Pleistocene,. L'ambiente di acqua dolce ha selezionato, nel tempo, popolazioni di pesci con corazze ossee formate da sempre minori placche, più piccole anche nelle dimensioni, in confronto ai membri delle popolazioni marine e oceaniche. Una placca ridotta significa un notevole risparmio di energia, una maturità sessuale più precoce e dimensioni corporee maggiori, un vantaggio, questo, nei confronti di possibili predatori come le larve di insetto. L'evoluzione degli spinarelli è stata osservata, si può dire, quasi in diretta nell'arco di poco più di un decennio, dal 1990 al Può sembrare sorprendente, ma questi pochi anni sono bastati perché la frequenza di individui con poche placche raggiungesse il 75%, in una popolazione di spinarelli marini che aveva colonizzato il lago Loberg, in Alaska. Il sequenziamento del genoma di diversi esemplari ha permesso di individuare alcuni geni responsabili della divergenza tra forme marine e forme di acqua dolce. In molti casi, si tratta di varianti non della parte codificante del gene, che contiene l'informazione per la sintesi della proteina, bensì delle sequenze regolatrici che, agendo come interruttori o modulatori, determinano la regolazione dell'espressione del gene stesso e dunque quanta proteina viene prodotta. Uno studio pubblicato su Proceedings of the National Academy of Sciences aggiunge, ora, un altro elemento alla comprensione della genetica alla base delle differenze morfologiche tra gli spinarelli. L'ipotesi degli autori era che l'adattamento all'ambiente di acqua dolce, dove questi pesci si nutrono di prede più grandi di quelle cacciate in mare, potesse avere modificato lo sviluppo dell'apparato mandibolare, in particolare della dentatura. Esaminando diversi esemplari, sia selvatici che allevati in laboratorio, gli autori hanno verificato che gli spinarelli di acqua dolce hanno un numero di denti quasi doppio rispetto a quelli marini, insieme a una maggiore area totale delle superfici dentali e a una maggiore densità di denti sulle mandibole. Il numero dei denti nelle due forme inizia a divergere solo verso la fine dello sviluppo embrionale e gli spinarelli di acqua dolce continuano ad aggiungere denti anche durante lo stadio adulto, mentre in quelli marini lo sviluppo si ferma. Studiando ibridi generati da incroci tra esemplari marini e di acqua dolce, che generano prole fertile, gli autori sono andati alla ricerca di quelle regioni del DNA che mostravano la maggiore correlazione con lo sviluppo dei denti. Essi hanno individuato una regione di particolare interesse sul cromosoma 21, dove si trova un gene, Bmp6 (bone morphogenetic protein 6), già noto per il suo ruolo nella determinazione del numero, della forma e delle dimensioni dei denti durante l'embriogenesi. Durante lo sviluppo degli spinarelli ibridi le due copie del gene Bmp6 mostrano lo stesso livello di espressione nello stadio larvale. Tuttavia, all'inizio dello stadio giovanile, proprio quando lo sviluppo dei denti nelle due forme diverge, la copia del gene Bmp6 proveniente dal genitore di acqua dolce inizia ad aumentare il livello di trascrizione e quindi di produzione della proteina. Non c'è alcuna differenza nella sequenza della proteina prodotta dai due alleli, ciò che li distingue sono le sequenze delle regioni regolatrici che li affiancano sul DNA e che determinano una espressione genica differente nei due alleli. Gli adattamenti degli spinarelli agli ambienti di acqua dolce dimostrano che l'insorgere di mutazioni nelle sequenze regolative, soprattutto in geni che presiedono allo sviluppo embrionale, possono accelerare il corso dell'evoluzione di una specie, causando, rapidamente, la perdita o l'acquisizione di nuovi caratteri.
, già noto per il suo ruolo nella determinazione del numero, della forma e delle dimensioni dei denti durante l embriogenesi. Durante lo sviluppo degli spinarelli ibridi le due copie del gene Bmp6 mostrano lo stesso livello di espressione nello stadio larvale. Tuttavia, all inizio dello stadio giovanile, proprio quando lo sviluppo dei denti nelle due forme diverge, la copia del gene Bmp6 proveniente dal genitore di acqua dolce inizia ad aumentare il livello di trascrizione e quindi di produzione della proteina. Non c è alcuna differenza nella sequenza della proteina prodotta dai due alleli, ciò che li distingue sono le sequenze delle regioni regolatrici che li affiancano sul DNA e che determinano una espressione genica differente nei due alleli. Gli adattamenti degli spinarelli agli ambienti di acqua dolce dimostrano che l insorgere di mutazioni nelle sequenze regolative, soprattutto in geni che presiedono allo sviluppo embrionale, possono accelerare il corso dell evoluzione di una specie, causando, rapidamente, la perdita o l acquisizione di nuovi caratteri.")
23
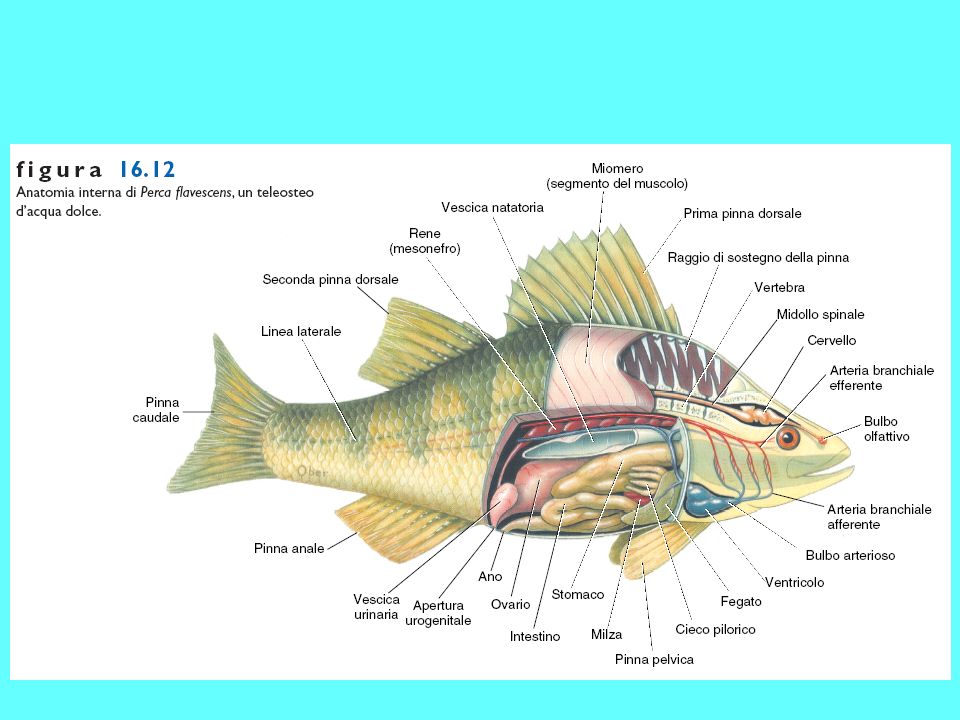
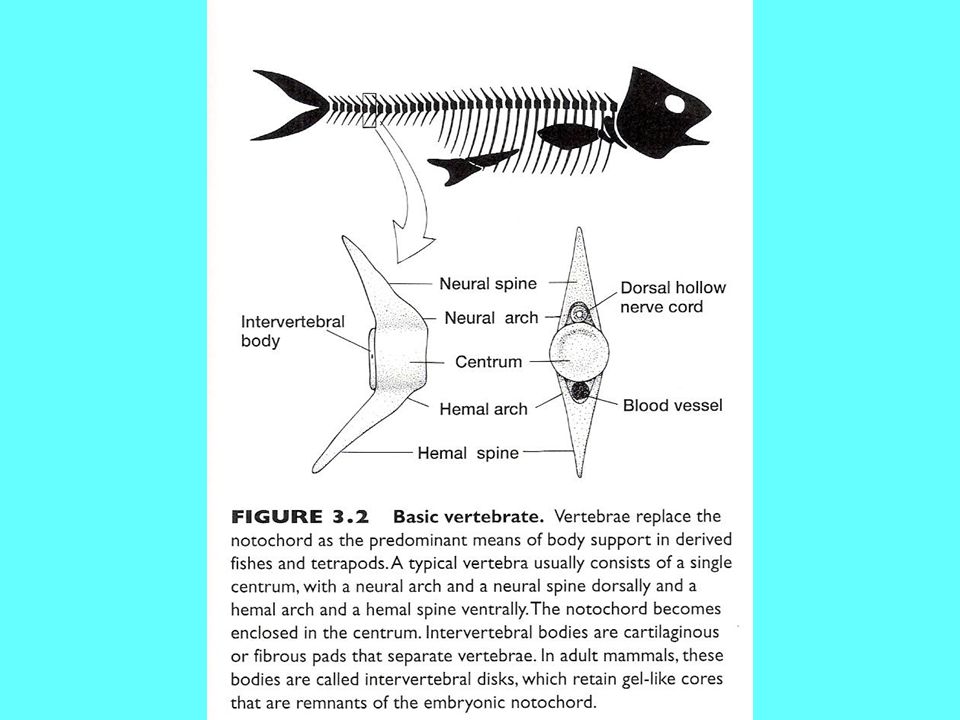
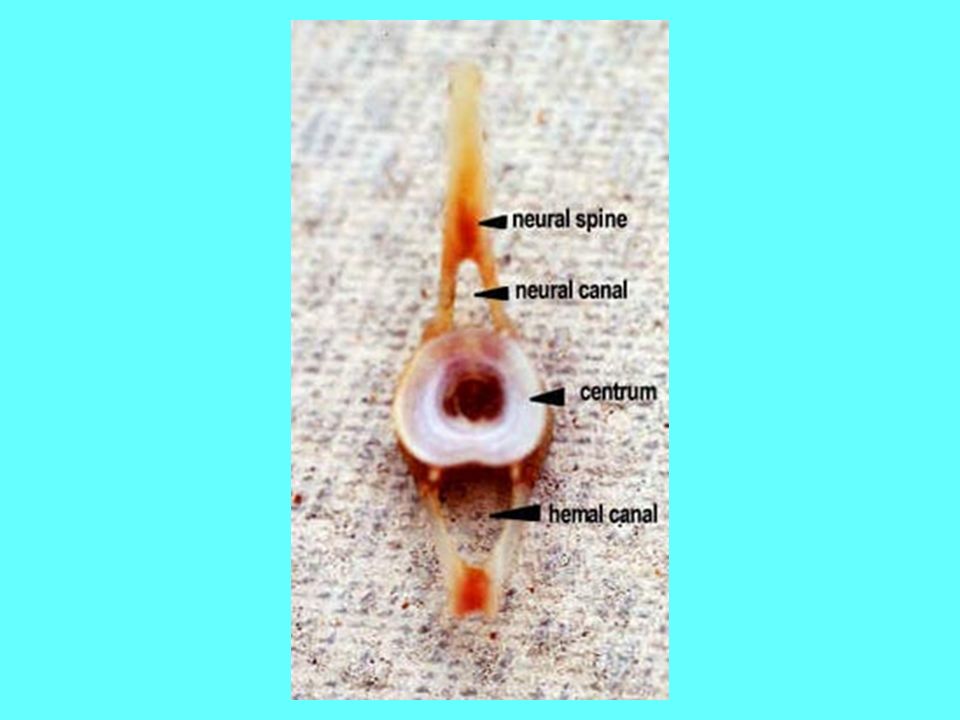
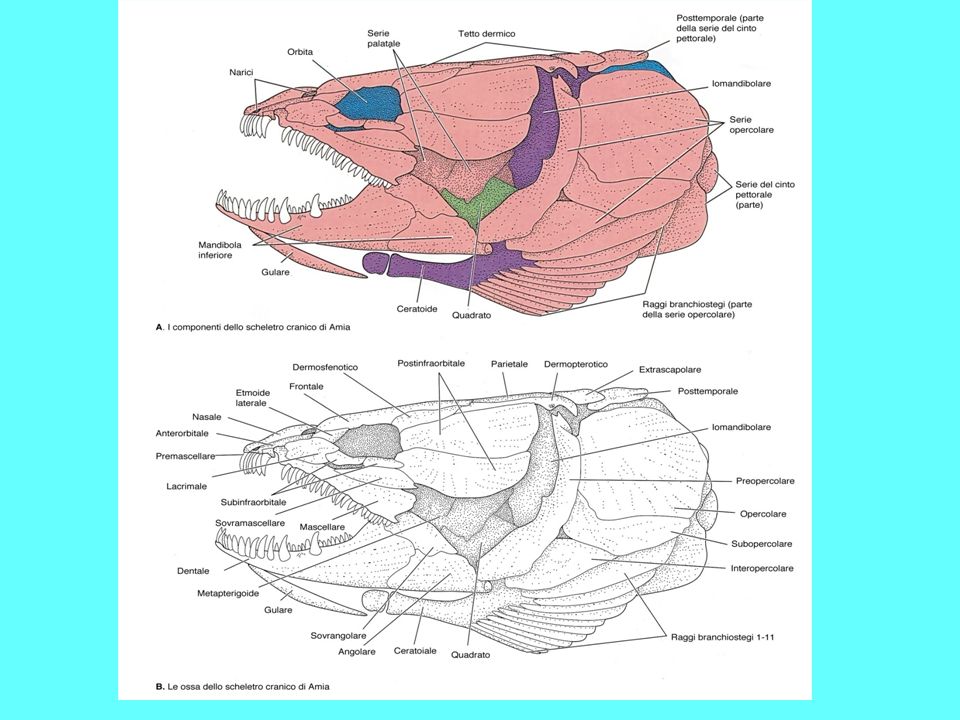
Scheletro di un Teleosteo

36
Branchie, polmoni e vescica natatoria negli Osteitti
Celacantidi Branchie Dipnoi polmoni branchie Condrostei Olostei Teleostei Ripidisti polmoni Branchie Crossopterigi Branchie + polmoni Sarcopterigi Branchie + polmoni Attinopterigi Branchie+vescica natatoria Primi Teleostomi Acantodi (?) Branchie + polmoni
 Branchie + polmoni.")
37
Sarcopterigi:caratteristiche generali
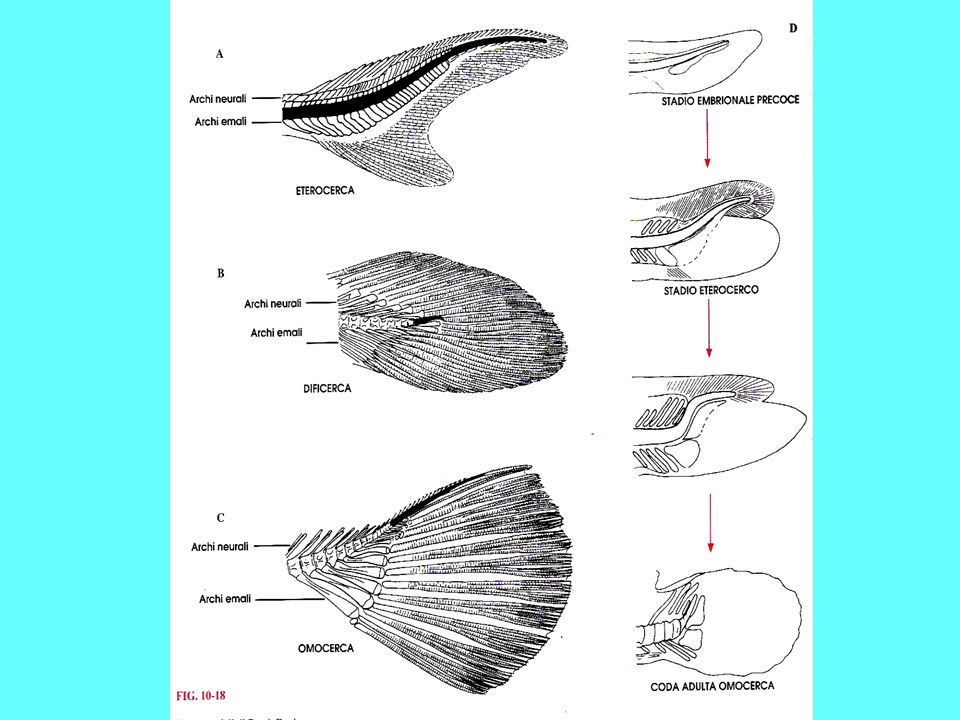
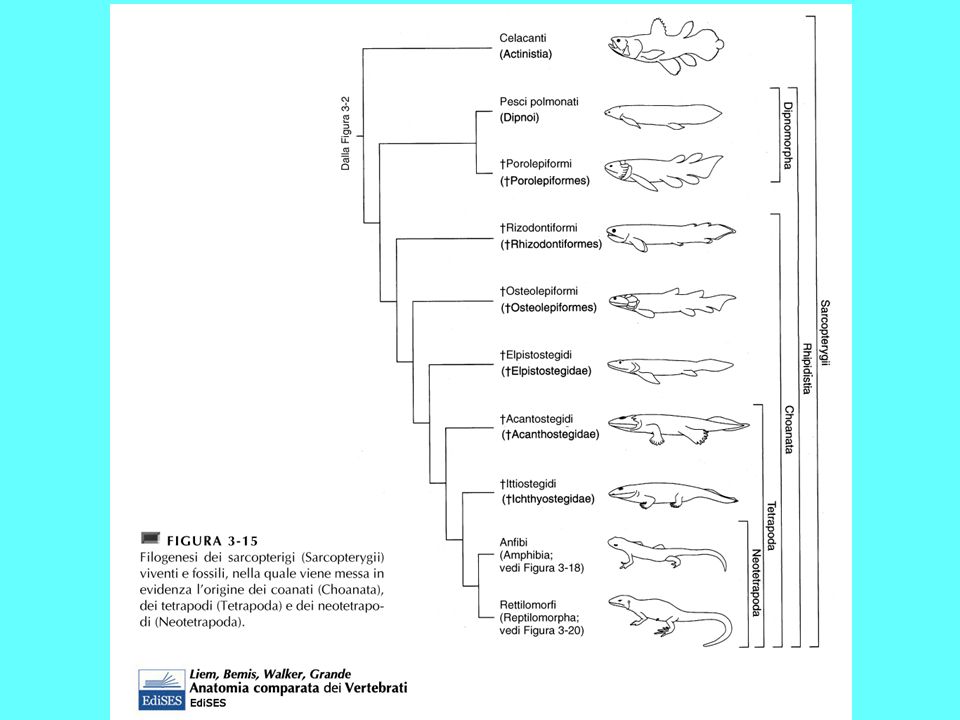
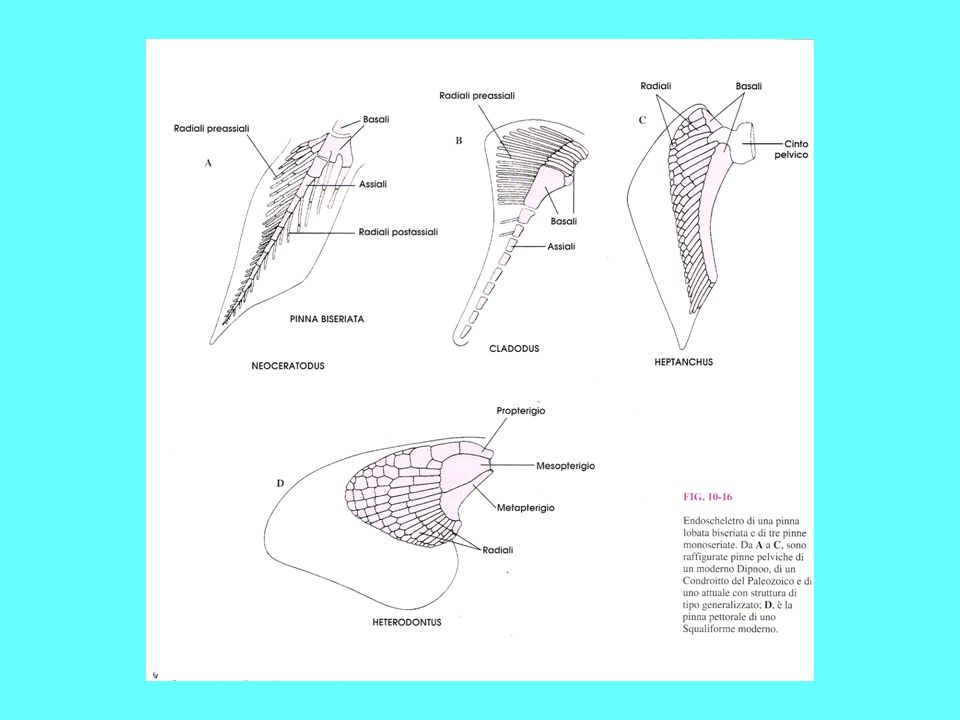
I sarcopetrigi comparvero nel Devoniano e decaddero nel Mesozoico I sarcopterigi comprendono: i Celacanti, i “Ripidisti”, collettivamente indicati in precedenza come Crossopterigi, e i Dipnoi Sono il sister group degli attinopterigi Sono pesci dotati di pinne carnose con endoscheletro, i cui pezzi sono disposti come gli elementi in una foglia, con una serie di elementi centrali disposti in fila da cui si dipartono una doppia serie di raggi. Tali pinne, robuste, potevano essere utilizzate per muoversi sul fondo La pinna caudale è di tipo dificerca I sarcoperigi sono pesci dotati di polmoni e/o vescica natatoria e di narici interne

41
La Latimeria: un Sarcopterigio Celacantide

42
Il termine “fossile vivente”, sebbene fosse già stato utilizzato da Darwin per indicare specie che hanno subito pochi cambiamenti durante il corso della loro evoluzione, tornò in auge con la scoperta della specie Latimeria calumnae (1938) fino ad allora conosciuta solo attraverso i reperti fossili. Da quel momento in poi, la Latimeria è stata inserita nella categoria degli organismi ritenuti “primitivi” (lamprede, squali, dipnoi, tuatara, solo per citare alcuni vertebrati), cioè che hanno subito pochi o nessun cambiamento nel tempo. Recentemente, Casane D. e Laurenti P. hanno cercato di dimostrare che il termine “fossile vivente” o altri suoi analoghi come “primitivo”, “poco evoluto”, “basale”,”ancestrale”, etc… non siano corretti. . La verità però è che come mostrato nel dettaglio dagli studiosi, non c’è alcuna prova paleontologica che dimostri che i celacanti attuali del genere Latimeria siano effettivamente uguali a qualche fossile di celacanto estinto; tanto diversi morfologicamente invece, che il genere Latimeria è stato costituito apposta per le due specie attualmente esistenti.. Non è vero dunque che esistano prove a favore del fatto che i celacanti siano rimasti in stasi morfologica dal Devoniano. Questa assunzione però è stata fatta, per esempio, nel lavoro comparso su Nature dedicato al sequenziamento del genoma di Latimeria, in cui si afferma che il lento tasso di evoluzione molecolare (tasso di sostituzione) riscontrato attraverso i dati genomici, spiega in qualche modo la lenta evoluzione morfologica di questa specie Tuttavia il concetto di “fossile vivente” ed affini sono più adatti ad una concezione progressionista dell’evoluzione, in cui la selezione, guidata da una sorta di forza finalistica, determina un incremento di complessità e perfezione di cui l’uomo è l’apice. Questa versione (errata) della teoria evoluzionistica è purtroppo molto più popolare di quella corretta, e vicina all’interpretazione di Haeckel, che concepisce la biodiversità come una scala evolutiva simile all’aristotelica “scala naturae”. Tuttavia molti passi avanti sono stati fatti, soprattutto dopo la nascita della cladistica che grazie a W. Hennig ha chiarito il processo di speciazione (cladogenesi), mostrando come una specie ancestrale cessi di esistere una volta formatesi le due specie “sorelle”. Inoltre molti importanti autori tra i quali vi sono Kimura e Gould, hanno rinforzato l’idea che l’evoluzione sia un processo continuo che non si arresta in nessuna linea. Purtroppo però questo paradigma filogenetico noto anche come tree-thinking, ha trovato molte difficoltà nel diffondersi non solo nell’opinione comune, ma perfino nella cerchia degli addetti ai lavori (Pikaia ne ha parlato qui). Numerosi infatti sono gli studi che usano organismi ritenuti “primitivi” o “meno evoluti”, come taxa attraverso i quali ricostruire i caratteri ancestrali, o che giustificano la scelta di un certa specie come modello di una certa classe di organismi, in base ad una presunta “basalità”. Non c’è alcuna ragione valida per cui la collocazione nell’albero filgenetico o la povertà di specie esistenti di un clade possa fornire informazioni sullo stato di primitività di un organismo in sé. Tutte le linee evolutive una volta staccatesi dall’antenato comune evolvono in modo indipendente e tutte le specie attualmente viventi sono un mosaico di caratteri primitivi (plesiomorfie) e derivati (apomorfie). Non esistono specie più o meno evolute, più o meno primitive, più o meno fossili. Siamo tutti egualmente evoluti, ogni specie con le proprie peculiarità
, cioè che hanno subito pochi o nessun cambiamento nel tempo. Recentemente, Casane D. e Laurenti P. hanno cercato di dimostrare che il termine fossile vivente o altri suoi analoghi come primitivo , poco evoluto , basale , ancestrale , etc… non siano corretti. . La verità però è che come mostrato nel dettaglio dagli studiosi, non c’è alcuna prova paleontologica che dimostri che i celacanti attuali del genere Latimeria siano effettivamente uguali a qualche fossile di celacanto estinto; tanto diversi morfologicamente invece, che il genere Latimeria è stato costituito apposta per le due specie attualmente esistenti.. Non è vero dunque che esistano prove a favore del fatto che i celacanti siano rimasti in stasi morfologica dal Devoniano. Questa assunzione però è stata fatta, per esempio, nel lavoro comparso su Nature dedicato al sequenziamento del genoma di Latimeria, in cui si afferma che il lento tasso di evoluzione molecolare (tasso di sostituzione) riscontrato attraverso i dati genomici, spiega in qualche modo la lenta evoluzione morfologica di questa specie Tuttavia il concetto di fossile vivente ed affini sono più adatti ad una concezione progressionista dell’evoluzione, in cui la selezione, guidata da una sorta di forza finalistica, determina un incremento di complessità e perfezione di cui l’uomo è l’apice. Questa versione (errata) della teoria evoluzionistica è purtroppo molto più popolare di quella corretta, e vicina all’interpretazione di Haeckel, che concepisce la biodiversità come una scala evolutiva simile all’aristotelica scala naturae . Tuttavia molti passi avanti sono stati fatti, soprattutto dopo la nascita della cladistica che grazie a W. Hennig ha chiarito il processo di speciazione (cladogenesi), mostrando come una specie ancestrale cessi di esistere una volta formatesi le due specie sorelle . Inoltre molti importanti autori tra i quali vi sono Kimura e Gould, hanno rinforzato l’idea che l’evoluzione sia un processo continuo che non si arresta in nessuna linea. Purtroppo però questo paradigma filogenetico noto anche come tree-thinking, ha trovato molte difficoltà nel diffondersi non solo nell’opinione comune, ma perfino nella cerchia degli addetti ai lavori (Pikaia ne ha parlato qui). Numerosi infatti sono gli studi che usano organismi ritenuti primitivi o meno evoluti , come taxa attraverso i quali ricostruire i caratteri ancestrali, o che giustificano la scelta di un certa specie come modello di una certa classe di organismi, in base ad una presunta basalità . Non c’è alcuna ragione valida per cui la collocazione nell’albero filgenetico o la povertà di specie esistenti di un clade possa fornire informazioni sullo stato di primitività di un organismo in sé. Tutte le linee evolutive una volta staccatesi dall’antenato comune evolvono in modo indipendente e tutte le specie attualmente viventi sono un mosaico di caratteri primitivi (plesiomorfie) e derivati (apomorfie). Non esistono specie più o meno evolute, più o meno primitive, più o meno fossili. Siamo tutti egualmente evoluti, ogni specie con le proprie peculiarità.")
43
Esempi di “ripidisti”

44
Eusthenopteron: un ripidista di 380 milioni di anni fa

45
Neoceratodus, Protopterus e Lepidosiren: 3 generi di Dipnoi (pesci polmonati)
Neoceratodus (Australia) vive nelle pozze d’acqua e sopravvive ingoiando aria Protopterus (Africa) e Lepidosiren (sud America) vivono in acque che vanno incontro a prosciugamento. Durante la stagione secca si ricoprono di fango e trascorrono il periodo secco in letargo.Il Protopterus è quello meglio adattato dei tre a vivere dentro un bozzolo,respirando l’aria durante i periodi di siccità
 vive nelle pozze d’acqua e sopravvive ingoiando aria. Protopterus (Africa) e Lepidosiren (sud America) vivono in acque che vanno incontro a prosciugamento. Durante la stagione secca si ricoprono di fango e trascorrono il periodo secco in letargo.Il Protopterus è quello meglio adattato dei tre a vivere dentro un bozzolo,respirando l’aria durante i periodi di siccità.")
46
Dipnoi Protopterus Neoceratodus

47
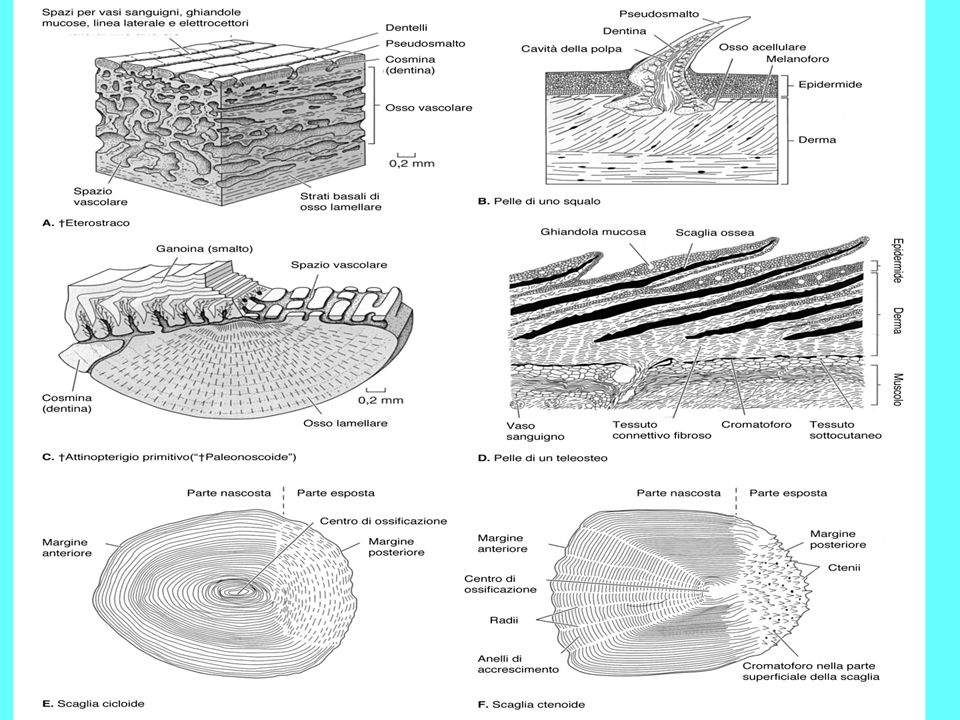
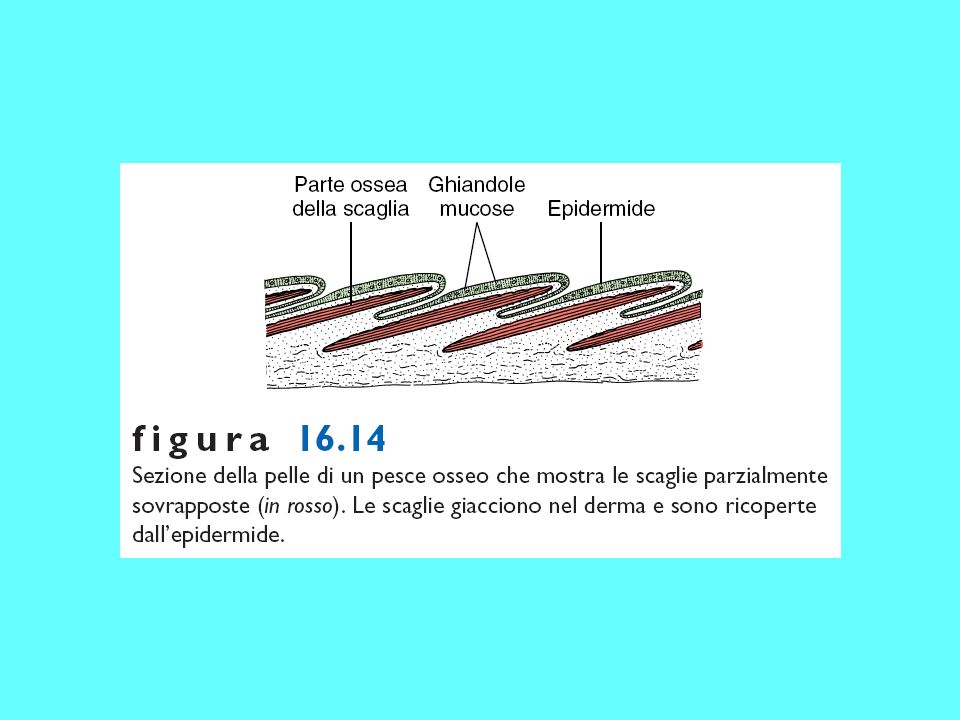
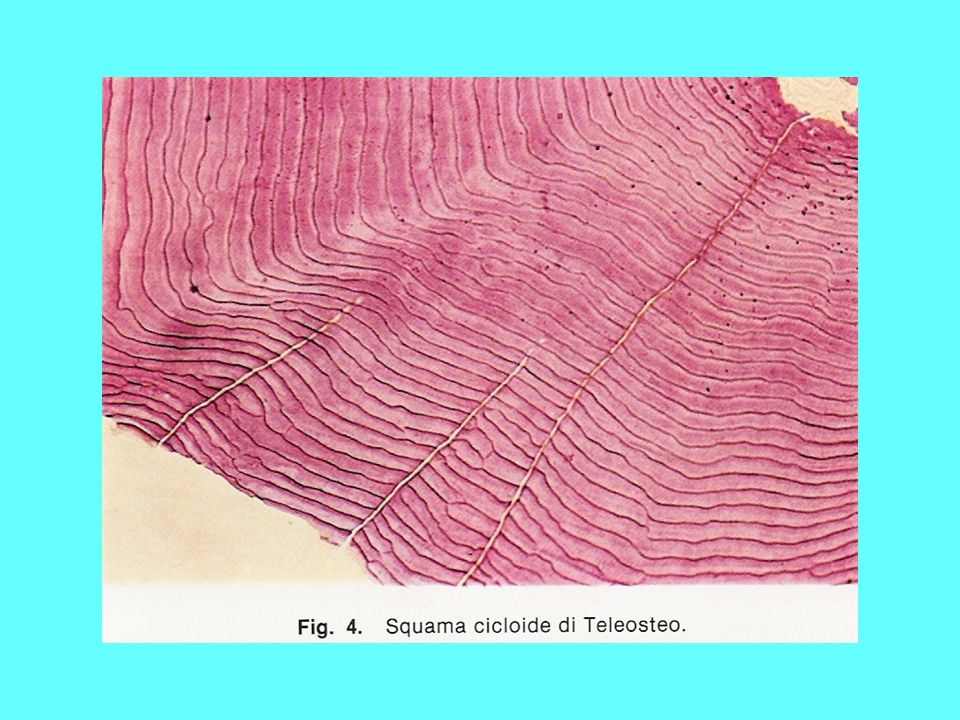
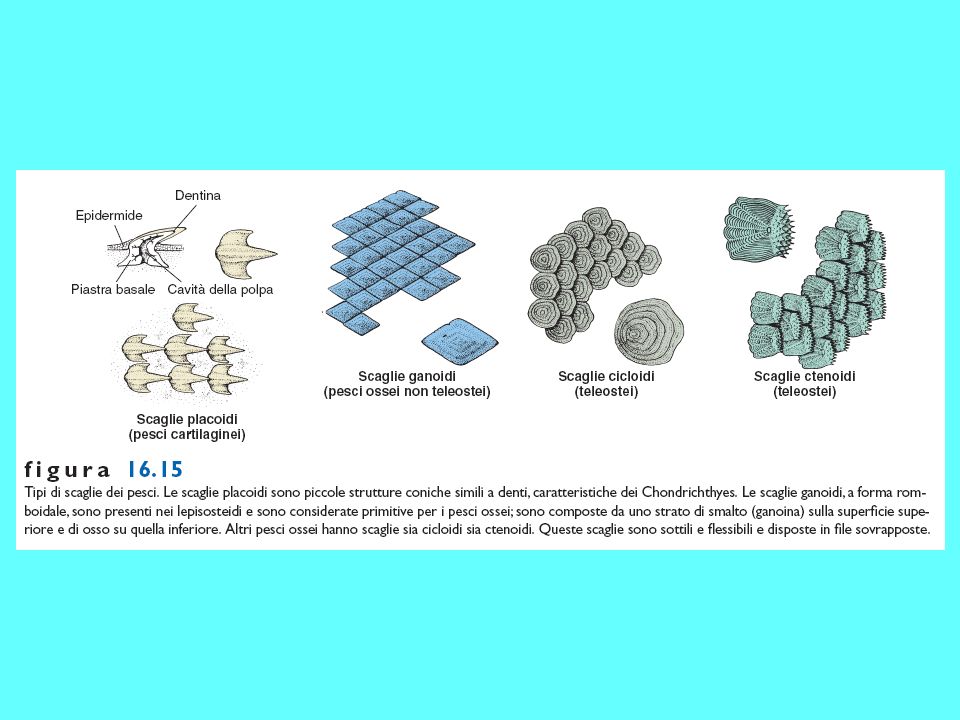
Scaglie degli osteitti sarcopterigi

49
Schema differenze tra Condroitti e Osteitti

Presentazioni simili













 mettere in discussione l’idea del progetto 2) rivalutare il.>")









>")




>")
