Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
COME ESCONO LE PROTEINE DAL RER?

2
VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE, VESCICOLE,

3
Cambio di strategia Il segnale di localizzazione passa dalla singola proteina alla singola vescicola

4
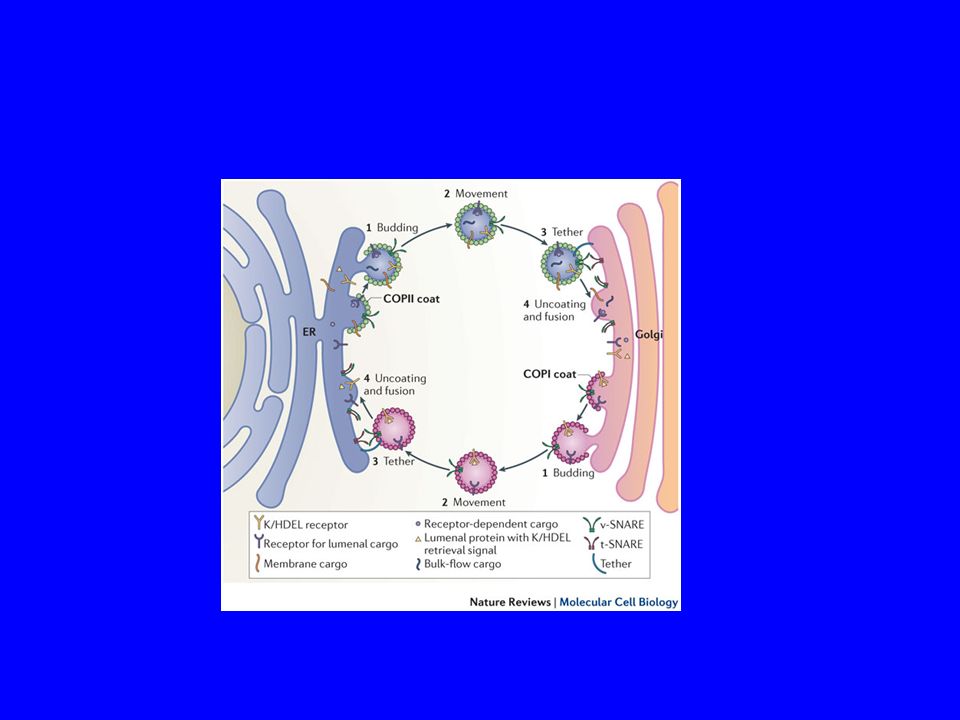
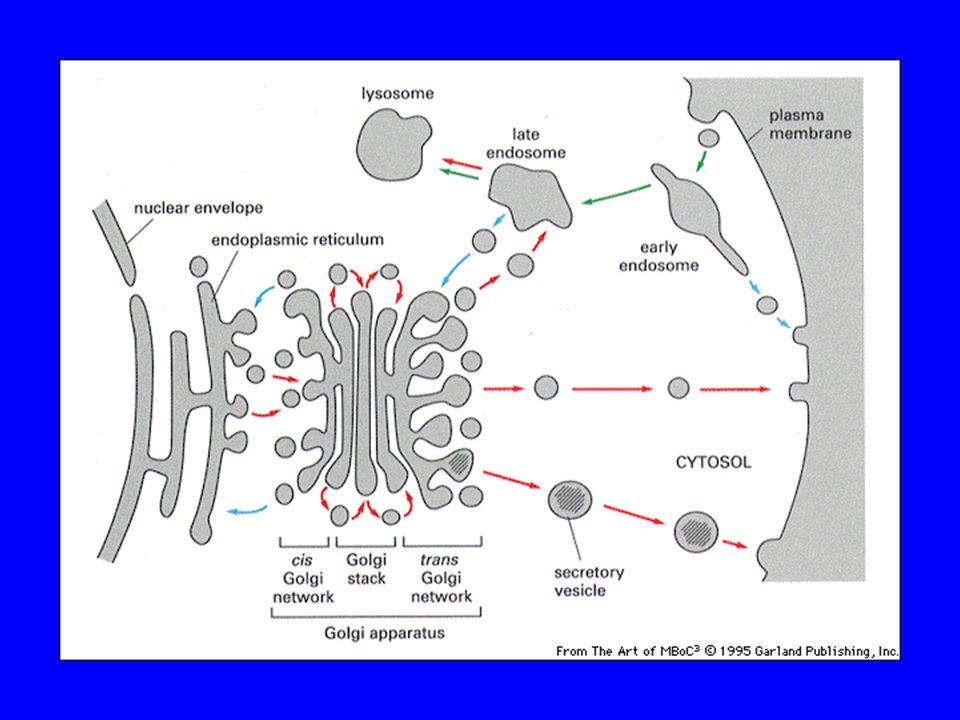
L’USCITA DAL RER AVVIENE MEDIANTE TRAFFICO VESCICOLARE
Figure Molecular Biology of the Cell (© Garland Science 2008)
")
5
COME SI FORMA UNA VESCICOLA?
QUANTI TIPI DI VESCICOLE VI SONO? COME SI DETERMINA SELETTIVITA’ DEL CONTENUTO? COME UNA VESCICOLA RICONOSCE IL BERSAGLIO? COME AVVIENE LA FUSIONE? COME VIENE REGOLATO L’INTERO PROCESSO?

6
COME ESCONO DAL RER LE PROTEINE
IN TRANSITO? o per selezioneBULK FLOW Secondo il modello «cattura del cargo», le proteine in uscita sono riconosciute da recettori specifici che le concentrano in siti specifici del ER Secondo il modello «flusso di massa» le proteine escono dal ER come parte del fluido o delle membrane delle vescicole in partenza con modalità indipendenti da recettore

7
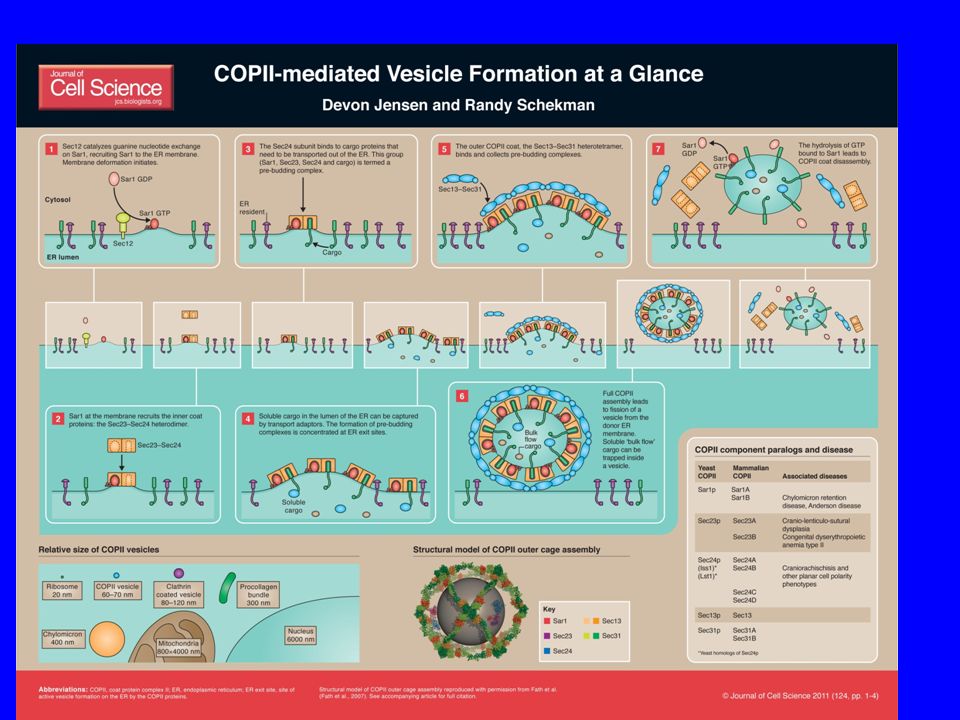
COME SI FORMA UNA VESCICOLA?

8
The vesicle cocktail (Dose for 2 students) 1 ER A bunch of proteins (soluble or membran bound) Some cargo protein receptor Small GTPase qb A few adaptors 1 Coat stirred and not shaken 1 orange slice Ice qb

9
Molti componenti sono necessari per la formazione di una vescicola

10
Come inizia la formazione delle vescicole
Primo step: reclutamento ed attivazione di small GTPase dalla membrana donatrice

11
Come si forma una vescicola
3. 2. 1. 1. Reclutamento di una small GTPase nella donor membrane (mediato dalla sua GEF) la small GTPase è nella forma GTP-bound 2. La small GTPase attivata media il reclutamento di coat proteins 3. Le subunità del coat legano selettivamente e reclutano I cargo nelle vescicole L’assemblaggio del coat porta a - reclutamento del cargo - curvatura della membrana
 la small GTPase è nella forma GTP-bound. 2. La small GTPase attivata media il reclutamento di coat proteins. 3. Le subunità del coat legano selettivamente e reclutano I cargo nelle vescicole. L’assemblaggio del coat porta a - reclutamento del cargo. - curvatura della membrana.")
12
LE PICCOLE GTPasi

13
Come viene attivata una GTPasi?
Inattiva GDP GTPasi Pi GDP GTP GAP GEF Attiva GTP GTPasi

14
Tre tipi di proteine di rivestimento intervengono nel traffico vescicolare
COPII COPI CLATRINA

15
Due tipi di proteine di rivestimento intervengono nel traffico vescicolare
ER-GOLGI-ER

16
COPII Sec23p/Sec24p eterodimero
Sec13p/Sec31p eterotetradimero Sar1-GTPasi Sec12, a transmembrane protein found in the ER acts as a Guanine nucleotide exchange factor by stimulating the release of GDP to allow the binding of GTP.

18
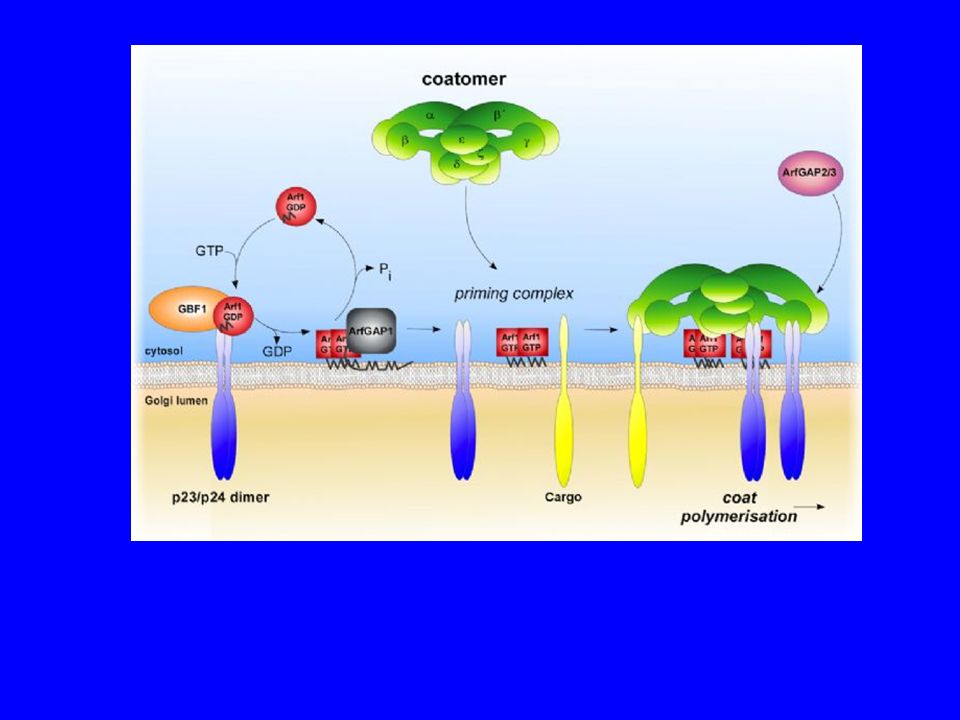
COPICoatomer

20
COPI

22
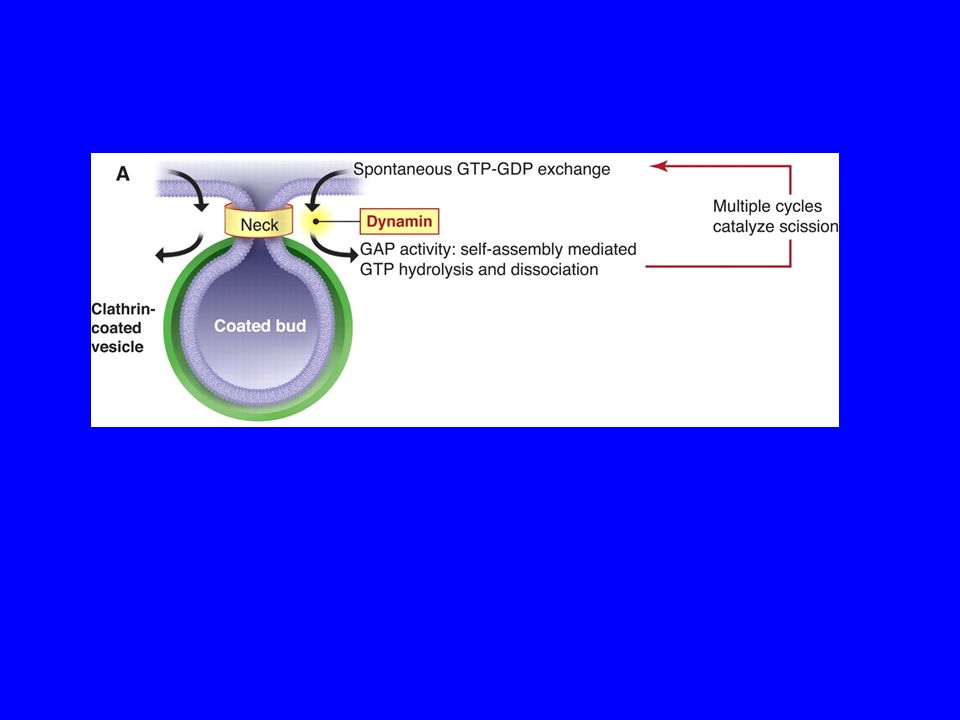
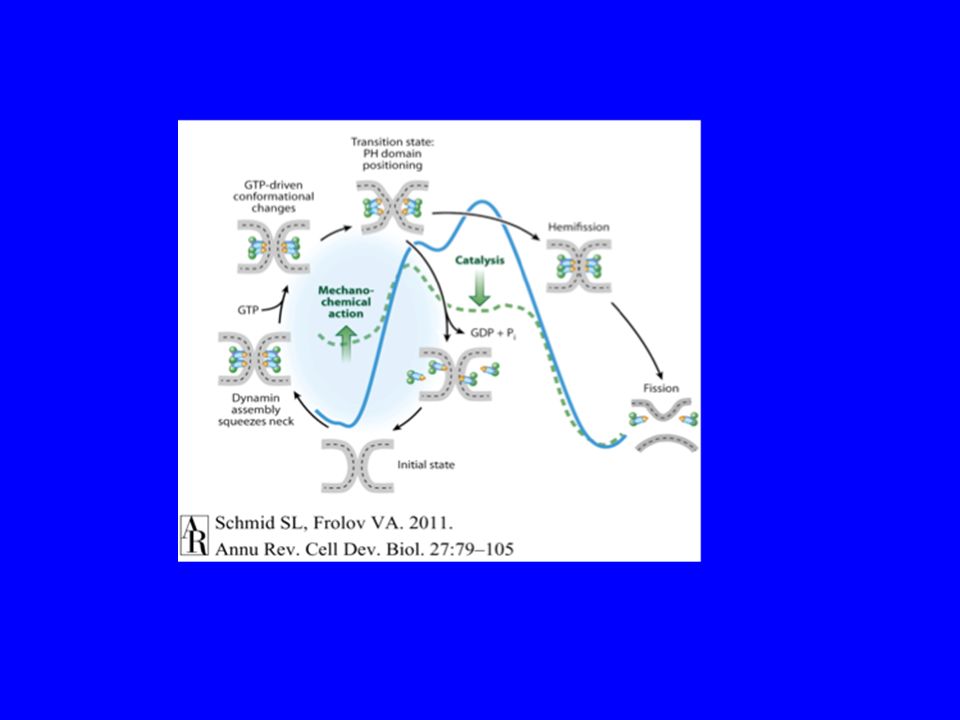
La fissione delle vescicole COPI e COPII

23
LE VESCICOLE RICOPERTE DI CLATRINA

24
Struttura di una vescicola rivestita di clatrina

25
Clathrin “pits” in the membrane where budding is initiated.
The triskelion structure of clathrin

26
Formazione di una vescicola rivestita di clatrina
Proteine di membrana Vescicola di clatrina

27
LE ADAPTINE mediano la formazione delle clathrin-coated pits, interagendo con receptors membrane-bound Vi sono molti diversi tipi di adattine ognuno relato ad uno specifico gruppo di recettori di membrana. La somiglianza nelle sequenze di adattine e COP 1 fanno pensare ad una origine evolutiva comune. Le adattine si aggregano a formare il complesso AP2 . Un eterotetramero di due adattine grandi (alpha or beta), una media (mu), ed una piccola (sigma) complex 1 AP1B1 AP1G1 AP1G2 AP1M1 AP1M2 AP1S1 AP1S2 AP1S3 complex 3 AP3B1 AP3B2 AP3M1 AP3M2 AP3S1 AP3S2 complex 2 AP2A1 AP2A2 AP2B1 AP2M1 AP2S1 complex 4 AP4B1 AP4E1 AP4M1 AP4S1
, una media (mu), ed una piccola (sigma) complex 1. AP1B1. AP1G1. AP1G2. AP1M1. AP1M2. AP1S1. AP1S2. AP1S3. complex 3. AP3B1. AP3B2. AP3M1. AP3M2. AP3S1. AP3S2. complex 2. AP2A1. AP2A2. AP2B1. AP2M1. AP2S1. complex 4. AP4B1. AP4E1. AP4M1. AP4S1.")
28
PROTEINE ACCESSORIE

29
Il rivestimento serve anche a selezionare il carico della vescicola
infatti le proteine di rivestimento interagiscono con il recettore specifico mediante le adattine

32
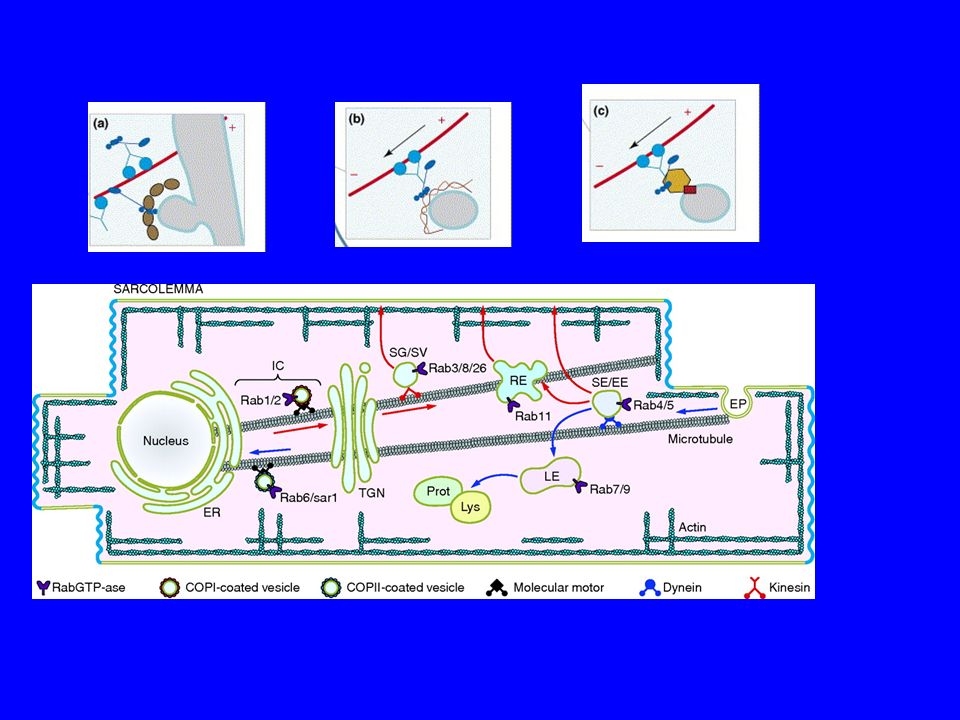
Dopo il distacco dalla sito donatore la vescicola si dirige lungo i
Microtubuli verso il sito accettore e intanto perde il coat L’uncoating è mediato da specifiche RAB La perdita del coat scopre le proteine chiave per la fusione

34
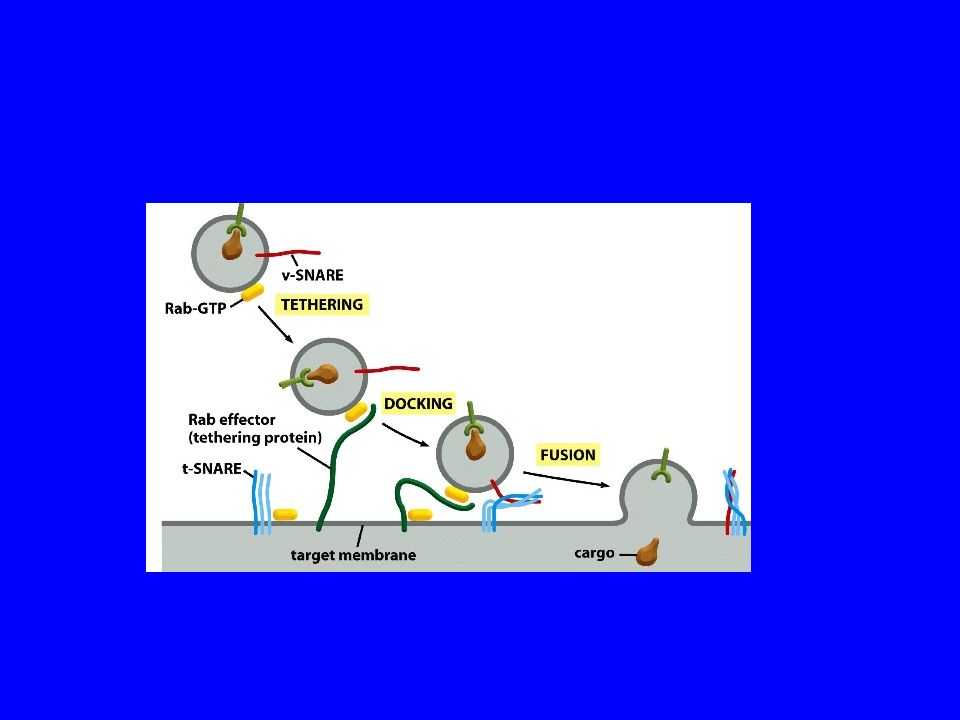
Come le vescicole riconoscono e legano il sito accettore?

35
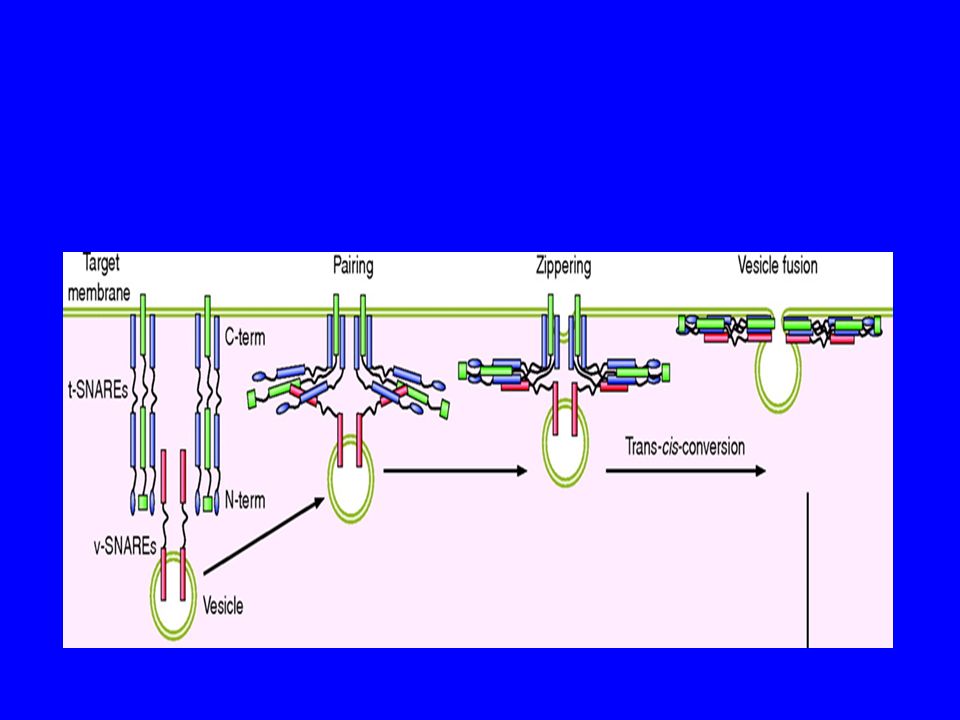
Ci sono proteine specifiche sia sulla vescicola che sul sito accettore che giocano un ruolo chiave nella fusione le SNARE

36
SNARES SNARE acronimo da "SNAP (Soluble NSFAttachment Protein) REceptor") NSF: enzima N-ethylmaleimide-sensitive factor, anche NSF or N-ethylmaleimide sensitive fusion proteins

37
V-SNARE T-SNARE Rab

39
TETHERING Attracco ancoraggio 3-Proteine coleid-coil
Il termine tethering factors indica un gruppo eterogeneo di proteine cellulari responsabile del legame iniziale delle vescicole di trasporto con le membrane bersaglio prima della loro fusione. Fino a poco tempo fa i tethering factors venivano divisi in due classi: complessi multi-subunità e coleid-coil proteine. Attualmente sono state individuate tre differenti classi funzionali: 1-Complessi oligomerici che legano le SNARE e che tipicamente agiscono come effettori RAB ( DCGE ) 2-Complessi oligomerici che funzionano come fattori di scambio guanina-nucleotide per le RAB (GEF) 3-Proteine coleid-coil Attracco ancoraggio
 2-Complessi oligomerici che funzionano come fattori di scambio guanina-nucleotide per le RAB (GEF) 3-Proteine coleid-coil. Attracco ancoraggio.")
41
Rab6 (blue) and Arl1 (gray) cooperate to bind GCC185 (green), a vesicle- transporting tether.
GM130 and/or Giantin binding may trigger a conformation change in p115, revealing a Rab1 binding site.

42
FUSIONE

43
2) The protein pore model: to suggest that
SNARE protein pairs interact and form “pore” or channel-like structure that initiates that fusion process so that the vesicle contents are not “spilled”. Summary: Coat is to specify cargo and SNARE is to match the cargo with the location.

44
Rab 27 Rab 3 Rab 8 Regolano la secrezione

45
A cycle of vesicle budding and fusion Sorting information:
Membrane proteins sorting signal is contained in the protein Soluble proteins Soluble cargo protein binds to a transmembrane receptor that contains a sorting signal.

46
DOVE SONO LOCALIZZATE LE Rab ?

48
TRA RER E APPARATO DI GOLGI ESISTE
UN COMPARTIMENTO INTERMEDIO IC ERGIC SALVAGE COMPARTMENT CGN EXOSOME

49
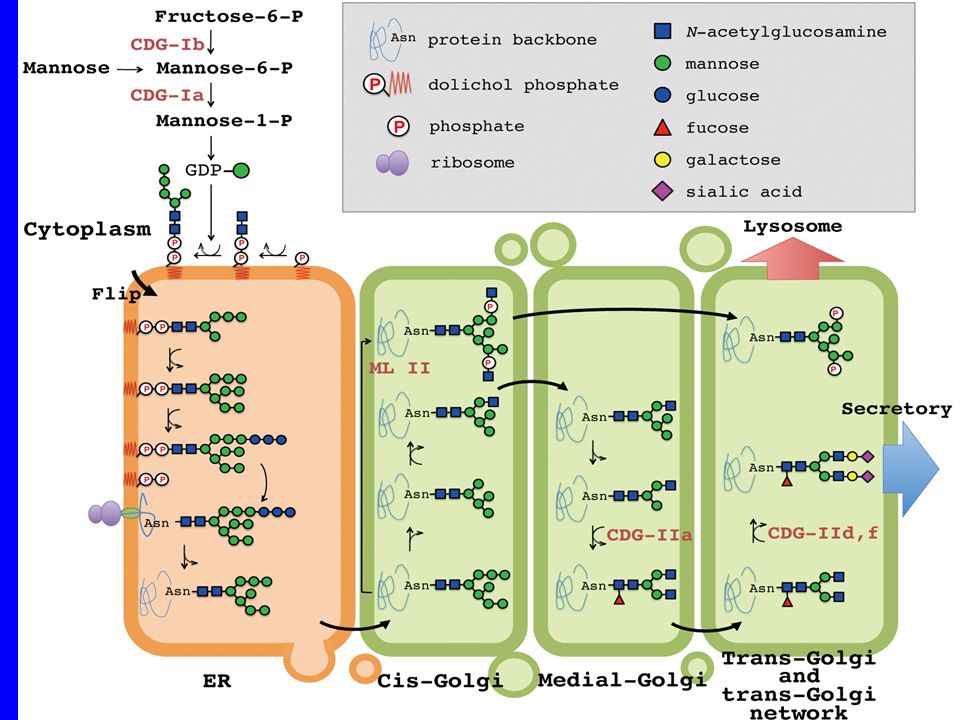
Il cluster tubolare vescicolare, indicato anche come ERGolgi intermediate compartment (ERGIC), media il traffico tra il reticolo endoplasmatico e complesso di Golgi, facilitando lo smistamento dei cargo. Scoperto nel 1984, il comparto è stato definito per la posizione di come la posizione della lectina di membrana mannosio specifica ERGIC 53. Vescicole COPII che escono dai siti di uscita del RE perdono il loro rivestimento e si fondono per formare il cluster tubolare vescicolare che sposta il suo contenuto nel cis Golgi.

50
COME VENGONO TRATTENUTE LE PROTEINE
RESIDENTI? SEGNALI DI RITENZIONE NEL LUME DEL RER: KDEL C-TERMINALE SEGNALI DI RITENZIONE NELLA MEMBRANA: KKXX XXRR KXKXX C-TERMINALE N-TERMINALE

51
LA SEQUENZA KDEL E’ UN SEGNALE DI
RITENZIONE NEL RER PDI ratto QKAVKDEL topo QKAVKDEL uomo QKAVKDEL pollo QKAVKDEL BiP ratto DTSEKDEL topo DTSEKDEL uomo DTAEKDEL pollo EAAEKDEL CALRETICULINA coniglio RRQAKDEL topo PAQAKDEL

52
What happens to proteins that have “escaped” the ER?
Retrieval signals: Proteins that predominantly function in the ER (such as BiP) contain retrieval signals (eg KDEL) recognized by a receptor mediate retrieval of escaped protein.
 contain retrieval signals (eg KDEL) recognized by a receptor. mediate retrieval of escaped protein.")
53
KISS AND RUN

54
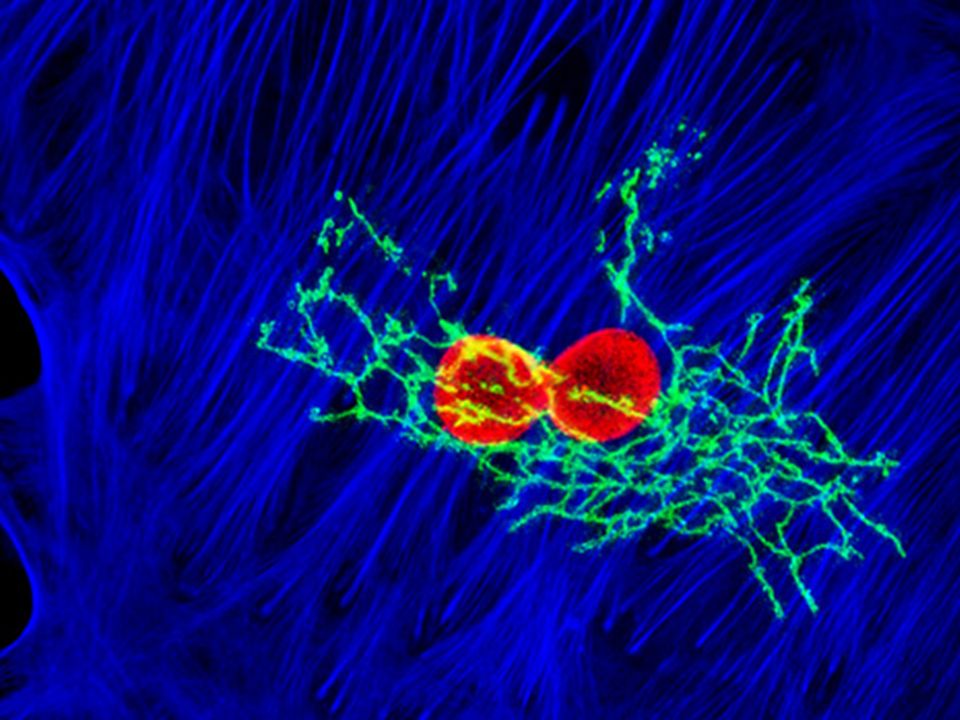
Il GOLGI

56
Ciascuna pila del Golgi ha due facce distinte: cis (di entrata) e trans (di uscita)
 e trans (di uscita)")
57
I componenti osservabili dal punto di vista ultrastrutturale nell’apparato di Golgi:
cisterne appiattite in pila microvescicole (vescicole di trasporto) macrovescicole (vescicole di secrezione) Il complesso di Golgi presenta anche una polarità interna anche a livello delle singole cisterne. Si possono quindi distinguere, in funzione anche dell’attività enzimatica e di sintesi: faccia cis: detta anche faccia prossimale faccia trans: detta anche faccia distale
 macrovescicole (vescicole di secrezione) Il complesso di Golgi presenta anche una polarità interna anche a livello delle singole cisterne. Si possono quindi distinguere, in funzione anche dell’attività enzimatica e di sintesi: faccia cis: detta anche faccia prossimale. faccia trans: detta anche faccia distale.")
58
La composizione biochimica delle cisterne è differente:
verso la parte prossimale o cis la membrana è ricca di lecitine verso la parte distale la membrana possiede parecchie sfingomieline e steroli, divenendo simile alla membrana plasmatica, con la quale le vescicole rilasciate dovranno fondersi

59
Ogni pila di cisterne dell’apparato di golgi è costituita da tre differenti compartimenti, ognuno dotato di uno specifico corredo enzimatico che catalizza le varie fasi di elaborazione delle glicoproteine giunte dal RER: cis mediano trans Questi tre compartimenti constano di una o due cisterne ciascuno e sono in rapporto tra loro mediante il distacco di vescicole laterali: le vescicole gemmano dai margini della cisterna e si fondono con la cisterna adiacente trasportano glicoproteine a differente stadio di elaborazione Nel Golgi si osservano sono due tipi di vescicole: vescicole transfer vescicole secretorie

61
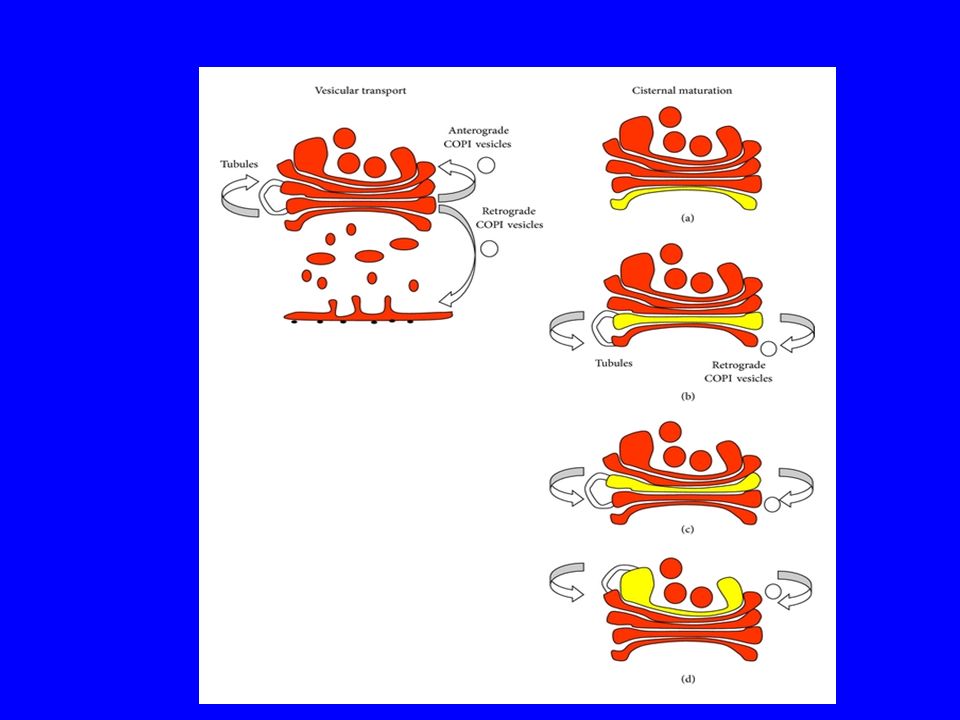
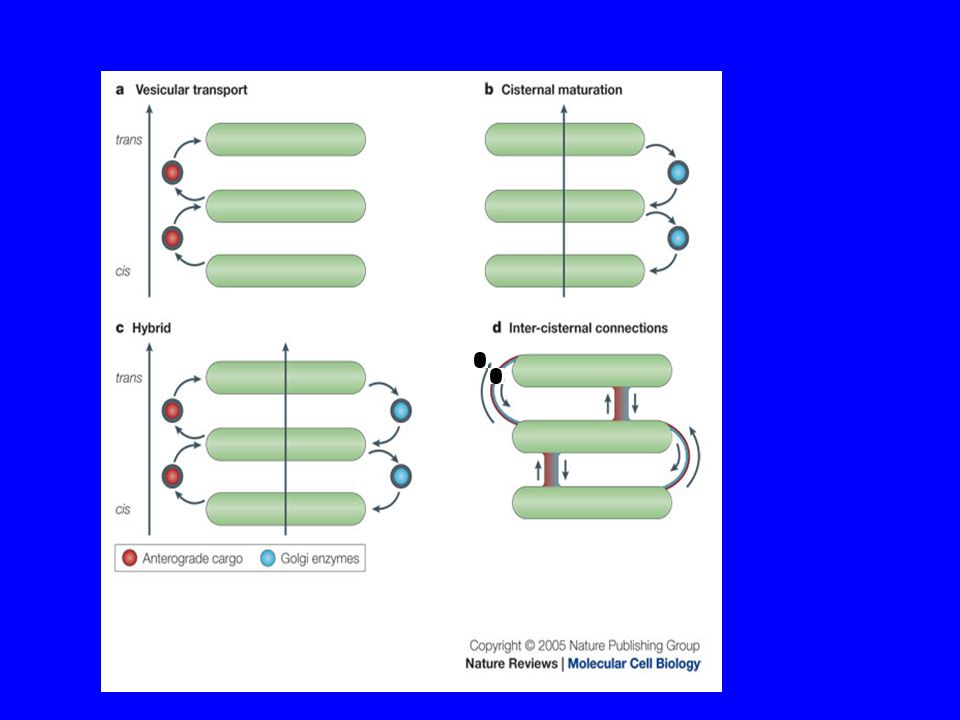
È più plausibile la teoria che le cisterne si scambino materiale attraverso vescicole piuttosto che il ricambio di materiale avvenga per trasformazione di un compartimento nei suoi vari stadi: le vescicole arrivano dagli organuli nel compartimento cis vengono elaborate e passate nel mediano attraverso piccole vescicole (laterali) vengono ulteriormente elaborate e passate al compartimento trans.
 vengono ulteriormente elaborate e passate al compartimento trans.")
64
Il complesso di Golgi è la sede in cui vengono convogliati e rielaborati i materiali sintetizzati nel RER e nel REL, inoltre attua specifiche attività di sintesi di polisaccaridi e lipidi complessi. Tutte le funzioni e la struttura dell’apparato di Golgi sono sotto il controllo diretto del nucleo: in corso di digiuno o di un inibitore della trascrizione le funzioni del Golgi si riducono considerevolmente (in relazione alla sintesi proteica effettuata)
")
65
Ogni compartimento del Golgi contiene enzimi differenti quindi ha una precisa competenza biochimica.
Questa compartimentazione rende possibile la simultanea elaborazone di tre tipologie di prodotti: quelli destinati ai lisosomi quelli secreti attraverso i granuli di secrezione quelli destinati al rinnovo dei componenti della membrana plasmatica

66
Le principali funzioni del Golgi
trasporto sorting trasformazione impacchettamento

67
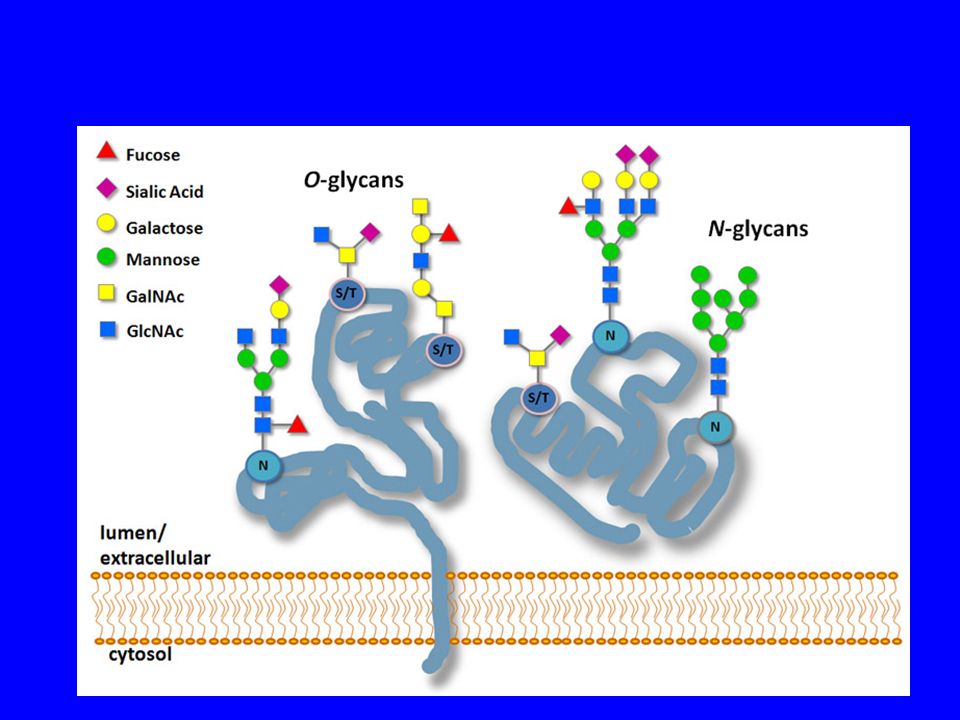
GLICOSILAZIONE DELLE PROTEINE
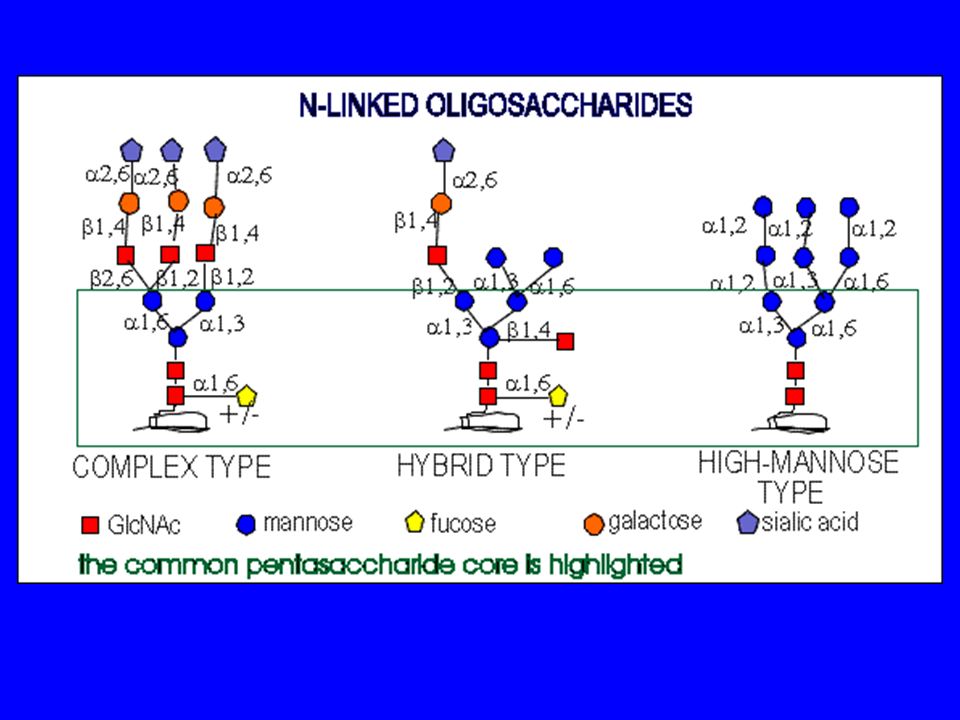
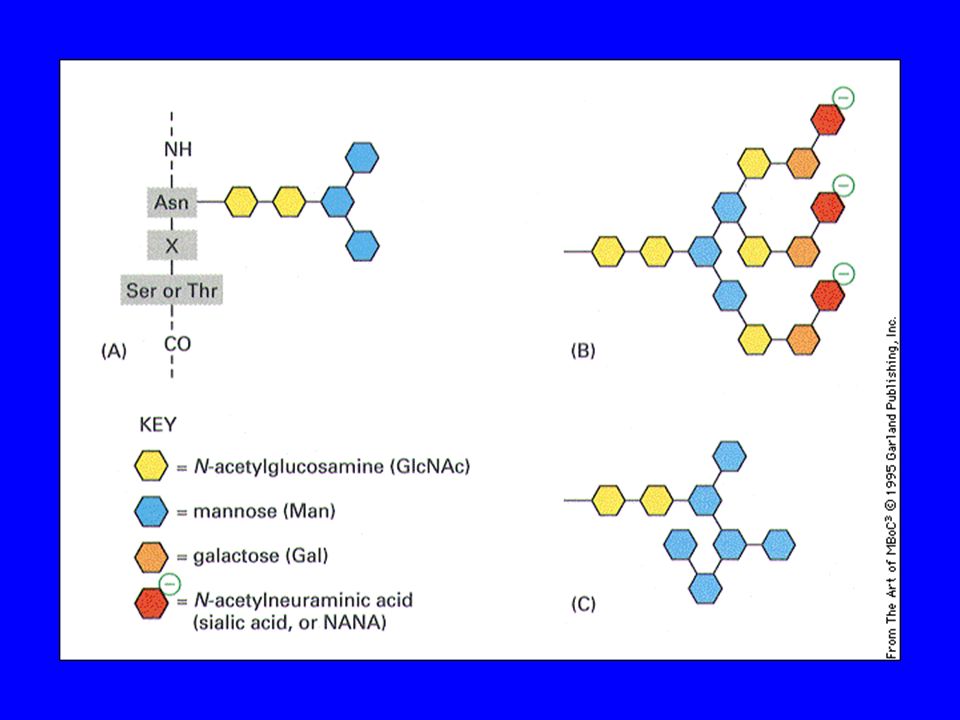
La maturazione delle proteine sintetizzate avviene preliminarmente nel RER e si completa nel Golgi, comprendendo una fase detta di glicosilazione la glicosilazione aggiunge catene laterali di carboidrati a specifici residui amminoacidici delle proteine esistono due tipi di glicosilazione: la glicosilazione legata ad azoto (N-glicosilazione) la glicosilazione legata ad ossigeno ( O-glicosilazione)
 la glicosilazione legata ad ossigeno ( O-glicosilazione)")
68
Modifications on the N-glycosilation pattern
cis-Golgi: mannose-type oligosaccharides complex oligosaccharides TGN: substitution with sialic acids - negatively charged

69
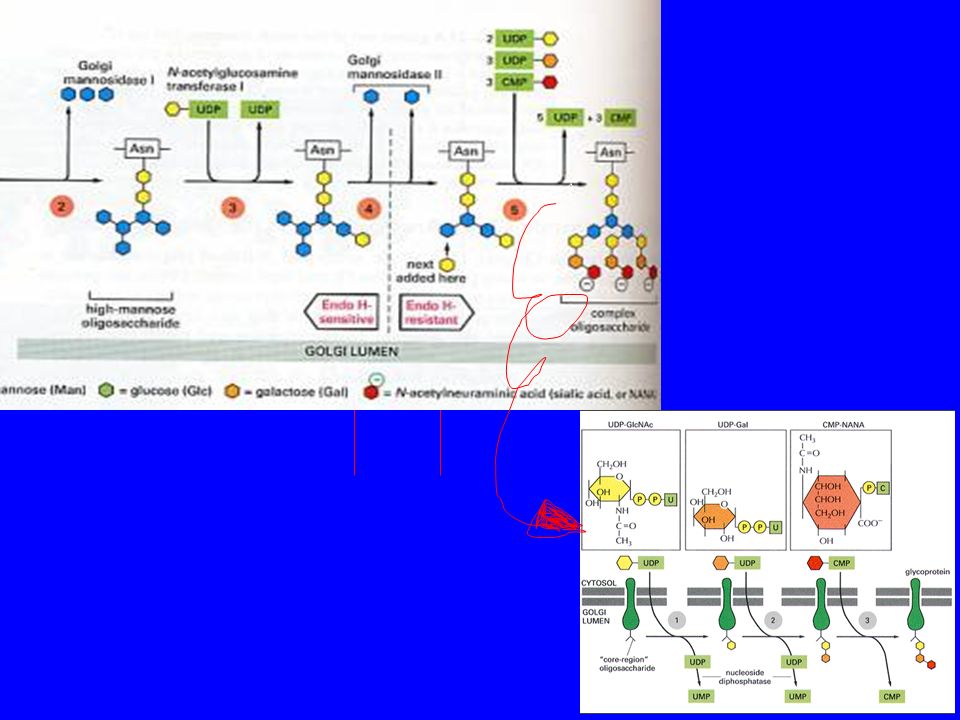
Tra glicosilazione nel ER e nel Golgi c’è una differenza fondamentale:
-nel reticolo endoplasmatico la glicosilazione è un evento "seriale", che non varia al variare del substrato, -nel Golgi ogni specifica proteina viene riconosciuta e modificata in base alla futura funzione. Si possono osservare infatti rimozioni o aggiunte di singoli zuccheri o di catene più lunghe la specificità delle singole catene glucidiche è il meccanismo utilizzato dalla cellula per lo smistamento delle proteine alle varie sedi di destinazione (lisosomi, membrana, perossisomi).
.")
70
Le proteine che hanno subito sia la glicosilazione che il primo rimaneggiamento nel ER trasportate nel Golgi,subiscono una sequenza ordinata di importanti cambiamenti. L’oligosaccaride attaccato in N nell’ER viene modificato: Una mannosidasi stacca 3 mannosi e degli zuccheri attaccati nell’ER restano solo 2 acetil glucosammine 5 mannosi

75
O-glicosilazione La O-glicosilazione è un processo altamente specifico, che si svolge completamente nell'apparato del Golgi Singoli zuccheri vengono legati uno per volta al peptide a livello dell'atomo di ossigeno delle catene laterali di serina o treonina. Solitamente il numero di zuccheri legati durante questo processo è limitato a pochi residui.

76
In O-glycans oligosaccharides are attached to a hydroxyl group of:
Serine or threonine ; then the first sugar residue is usually N-Acetylgalactosamine (GalNAc). Less commonly, galactose, mannose or xylose form O-glycosidic bonds with Ser or Thr. Although most cytosolic and nuclear proteins are not glycosylated, when they are, a single N-Acetylglucosamine residue is linked to the Ser or Thr hydroxyl group. These exceptions include some nuclear-pore complex proteins and some transcription factors. Hydroxylysine (Hyl); it is glycosylated by the attachment of single Gal residue or glucosylgalactose disaccharide. Hydroxyproline (Hyp); arabinose residue is linked to it. Hyl and Hyp residues occur only in collagens. O-linked oligosaccharides are generally short (1-4 sugar residues). But for example o-glycans of ABO blood group antigens are longer. The longest O-linked carbohydrate chains occur in proteoglycans. They contain up to 1000 disaccharide units.
. Less commonly, galactose, mannose or xylose form O-glycosidic bonds with Ser or Thr. Although most cytosolic and nuclear proteins are not glycosylated, when they are, a single N-Acetylglucosamine residue is linked to the Ser or Thr hydroxyl group. These exceptions include some nuclear-pore complex proteins and some transcription factors. Hydroxylysine (Hyl); it is glycosylated by the attachment of single Gal residue or glucosylgalactose disaccharide. Hydroxyproline (Hyp); arabinose residue is linked to it. Hyl and Hyp residues occur only in collagens. O-linked oligosaccharides are generally short (1-4 sugar residues). But for example o-glycans of ABO blood group antigens are longer. The longest O-linked carbohydrate chains occur in proteoglycans. They contain up to 1000 disaccharide units.")
78
Glycophosphatidylinositol (GPI) membrane anchors are attached to polypeptide chain through an amide bond between mannose-6-phosphoethanolamine and the C-terminal carboxyl group :
 membrane anchors are attached to polypeptide chain through an amide bond between mannose-6-phosphoethanolamine and the C-terminal carboxyl group :")
79
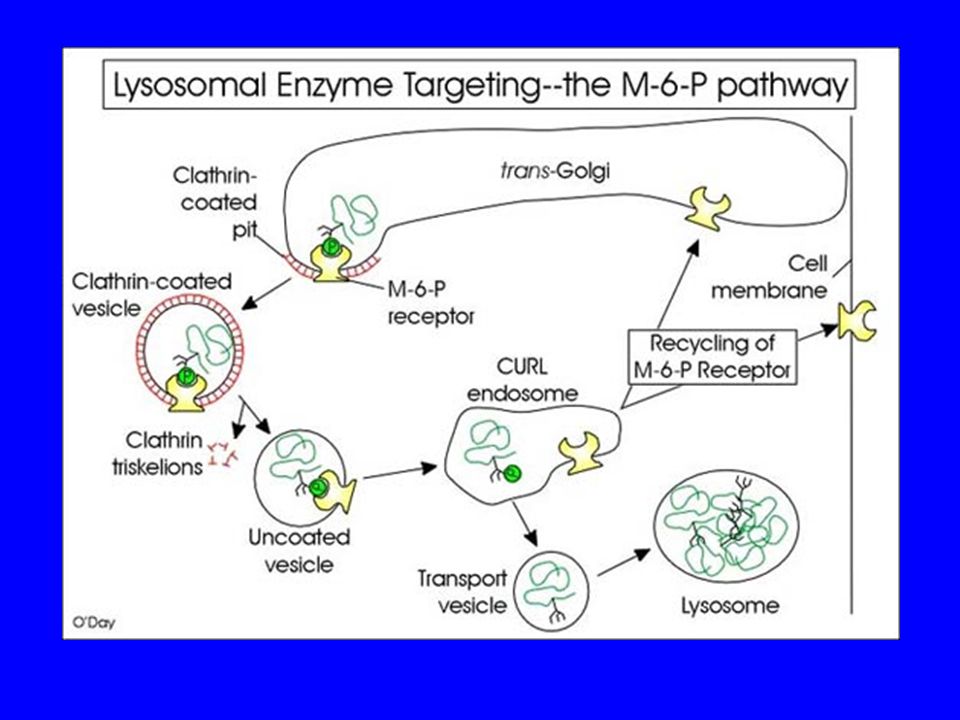
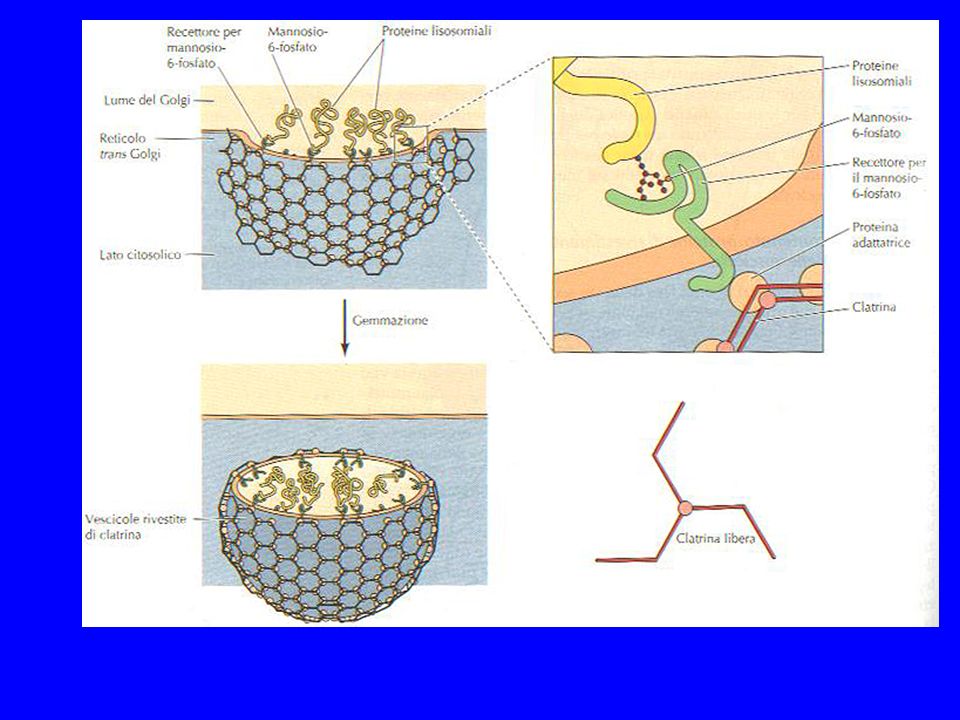
… e poi ci sono e proteine destinate al lisosoma sono caratterizzate dalla presenza di un gruppo specifico, il mannosio-6-fosfato (M6P), che serve a farle riconoscere da un recettore specifico presente sul trans-Golgi. MANNOSIO fosfotrasferasi UDP-GlcNAc UMP MAN-6-P-GlcNAc MAN-6-P fosfoglicosidasi GlcNAc PROTEINA L’M6P è aggiunto agli oligosaccaridi legati all’asparagina, nel reticolo cis del Golgi.

80
FOSFORILAZIONE DI ENZIMI LISOSOMIALI

81
I recettori sono riciclati verso il trans Golgi.
Il recettore è una proteina transmembrana localizzata sul trans Golgi. L’unione con l’idrolasi inizia la formazione di vescicole rivestite di clatrina, e si verifica a pH 7. Le idrolasi si dissociano dai recettori negli endosomi tardivi a pH 6. I recettori sono riciclati verso il trans Golgi.

84
Lisosomi Svolgono la funzione di “sistema digestivo” della cellula, degradando sia materiale trasportato dall’esterno della cellula, che componenti cellulari non più utili. Tutti gli enzimi dei lisosomi sono idrolasi acide, attive al pH acido dei lisosomi (circa 5.0) ,ma non al pH neutro del citoplasma (7,2). Questo meccanismo protegge la cellula dalla eventuale rottura della membrana del lisosoma. Infatti le idrolasi rilasciate sarebbero inattive al pH neutro del citosol IDROLASI ACIDE: Nucleasi, Proteasi, Glicosidasi, Lipasi, Fosfatasi, Solfolipasi Fosfolipasi
 ,ma non al pH neutro del citoplasma (7,2). Questo meccanismo protegge la cellula dalla eventuale rottura della membrana del lisosoma. Infatti le idrolasi rilasciate sarebbero inattive al pH neutro del citosol. IDROLASI ACIDE: Nucleasi, Proteasi, Glicosidasi, Lipasi, Fosfatasi, Solfolipasi. Fosfolipasi.")
85
Per mantenere acido il ph al loro interno i lisosomi devono attivamente concentrare ioni H+. Ciò è assicurato dalla presenza nella membrana di una pompa protonica, che trasporta attivamente protoni dal citosol nei lisosomi. L’attività di questa pompa richiede consumo di energia che è fornita da idrolisi di ATP per mantenere nei lisosomi una concentrazione di H+ circa 100 volte più alta rispetto al citosol LISOSOMA Idrolasi acide pH 5 H+ CITOSOL ATP ADP H+ pH 7

86
Nei lisosomi convergono le sostanze che devono essere digerite ad opera delle idrolasi lisosomali
Vie per arrivare al lisosoma: Autofagia (ER si trasforma in lisosoma) Fagocitosi Endosoma precoci e tardivi Alcune proteine sono trasportate nel lisosoma direttamente con l’uso di un segnale lisosomale: KFERQ (Lys, Phe, Glut, Arg, glutammina)
 Fagocitosi. Endosoma precoci e tardivi. Alcune proteine sono trasportate nel lisosoma direttamente con l’uso di un segnale lisosomale: KFERQ (Lys, Phe, Glut, Arg, glutammina)")
87
FAGOCITOSI Cellule specializzate nella degradazione di particelle di grandi dimensioni e di microrganismi (es. macrofagi), fagocitano al loro interno queste particelle formando un FAGOSOMA, il quale si fonde con un lisosoma assicurando la digestione del contenuto. I lisosomi derivati da questo processo prendono il nome di FAGOLISOSOMI e possono essere di varie forme e dimensioni, in base al tipo di materiale fagocitato. Le sostanze indigeribili permangono nei lisosomi quali corpi residui.
, fagocitano al loro interno queste particelle formando un FAGOSOMA, il quale si fonde con un lisosoma assicurando la digestione del contenuto. I lisosomi derivati da questo processo prendono il nome di FAGOLISOSOMI e possono essere di varie forme e dimensioni, in base al tipo di materiale fagocitato. Le sostanze indigeribili permangono nei lisosomi quali corpi residui.")
88
AUTOFAGIA Rappresenta la via degradativa degli organuli cellulari. Essi vengono inglobati in membrane derivanti dal RE e la vescicola così formata ( AUTOFAGOSOMA), si fonde con un lisosoma , degradando il proprio contenuto
, si fonde con un lisosoma , degradando il proprio contenuto.")
89
MALATTIE LISOSOMIALI DA ACCUMULO
Gravi malattie genetiche causate dal difetto di enzimi che hanno il compito di degradare diverse molecole all’interno della cellula. I composti non degradati si accumulano nei lisosomi e ne alterano la funzione. CLASSIFICAZIONE MUCOPOLISACCARIDOSI OLIGOSACCARIDOSI SFINGOLIPIDOSI Hanno un decorso progressivo che porta al deterioramento delle funzioni vitali. Molte di esse sono letali. Ereditate come m.autosomiche recessive, si presentano con una frequenza di 1/5000 nati vivi.

90
MALATTIA DI FABRY La malattia di Fabry, nota anche come malattia di Anderson-Fabry, è una malattia ereditaria multisistemica del metabolismo degli sfingolipidi. E’ una patologia legata al cromosoma X, causata da mutazioni a carico del gene codificante per l’enzima alfa-galattosidasi A che alterano o aboliscono l'attività enzimatica.

91
I-cell disease MALATTIA DI HURLER
La sindrome è dovuta a mutazioni del gene IDUA che esitano nella totale assenza dell'enzima alfa L-iduronidasi e nell'accumulo nei lisosomi di dermatan solfato ed eparan solfato. La trasmissione è autosomica recessiva. L'incidenza della malattia risulta un caso su persone I-cell disease I substrati delle idrolasi si accumulano non digeriti nel lisosomi, per cui si formano inclusioni nelle cellule. Le idrolasi alterate sfuggono ai lisosomi, sono secrete dalla via di default e si accumulano nel sangue: il difetto è dovuto alla mancanza o al difetto di una fosfotransferasi GlcNAc. Le idrolasi sono secrete.

92
Other modifications glucose-amino-glycane (GAG) chains
sulphatation (proteoglycanes, Tyr res. of peptides) - TGN proteolytic modifications - secretion vesicle
 - TGN. proteolytic modifications - secretion vesicle.")
93
Synthesis of lipids in the Golgi
ceramide glycolipids sphingomyelin

94
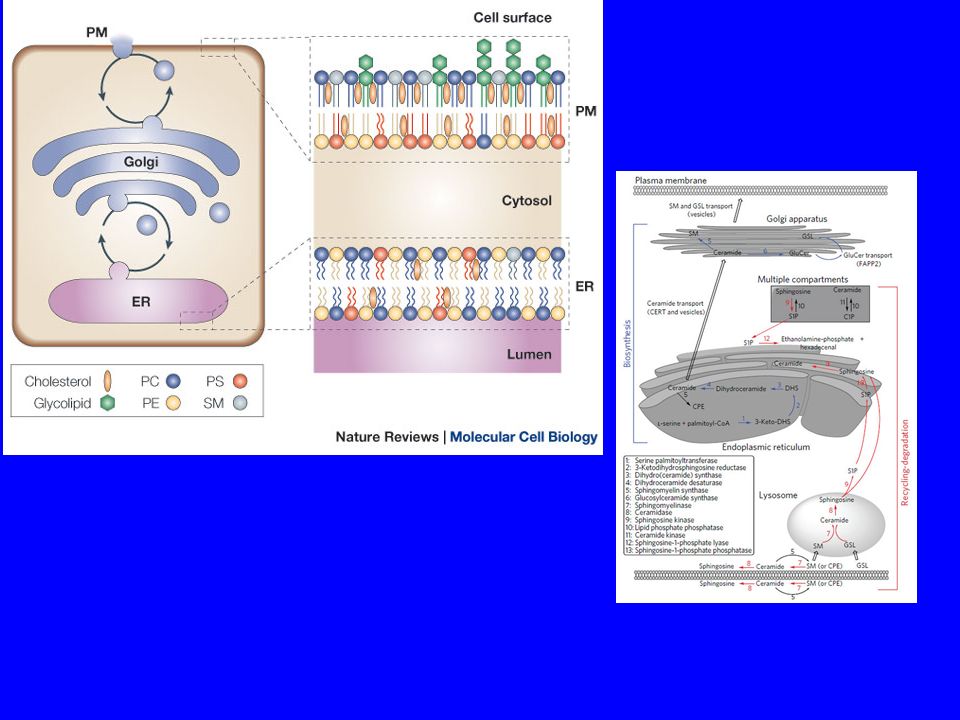
Mechanisms of intermembrane lipid transport.
CERT Il trasporto della ceramide dal RE al Golgi è un punto critc0 Mechanisms of intermembrane lipid transport. (A) Membrane transport moves lipids together with proteins in vesicular and tubular carriers that bud off from a donor membrane, and are transported along cytoskeletal tracks to the acceptor membrane, in which they fuse to deliver their cargo. (B) Cytosolic carrier proteins transfer lipids in hydrophobic pockets that show selectivity toward one or a few lipid types. Carrier proteins often contain domains that bind to the donor and/or acceptor membranes. (C) Lipid exchange may also occur between membranes that are in very close proximity. Transfer via such membrane contact sites may be facilitated by carrier proteins (combination of models B and C). Tomas Blom et al. Cold Spring Harb Perspect Biol 2011;3:a004713 ©2011 by Cold Spring Harbor Laboratory Press
 Membrane transport moves lipids together with proteins in vesicular and tubular carriers that bud off from a donor membrane, and are transported along cytoskeletal tracks to the acceptor membrane, in which they fuse to deliver their cargo. (B) Cytosolic carrier proteins transfer lipids in hydrophobic pockets that show selectivity toward one or a few lipid types. Carrier proteins often contain domains that bind to the donor and/or acceptor membranes. (C) Lipid exchange may also occur between membranes that are in very close proximity. Transfer via such membrane contact sites may be facilitated by carrier proteins (combination of models B and C). Tomas Blom et al. Cold Spring Harb Perspect Biol 2011;3:a ©2011 by Cold Spring Harbor Laboratory Press.")
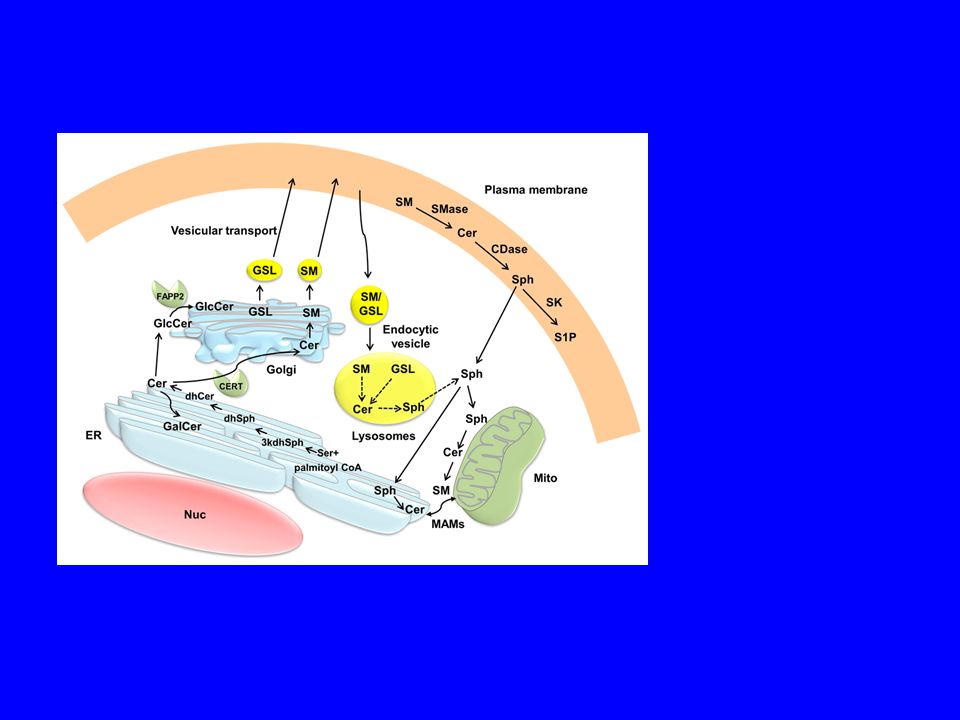
95
Biosynthetic trafficking of major membrane lipids.
Biosynthetic trafficking of major membrane lipids. (Left) Glycerophospholipids are synthesized in the ER, with phosphatidylserine synthases enriched in mitochondria associated membrane fractions. Glycerophospholipids are transported from the ER both along exocytic membrane transport and by nonvesicular, yet poorly characterized mechanisms. (Middle) De novo sphingolipid synthesis is initiated in the ER. The ER to Golgi transport of ceramide for the assembly of more complex sphingolipids is mediated by CERT, and to a lesser extent by membrane transport. The post-Golgi transport of complex sphingolipids is dependent on membrane transport. Sphingosine that stems from lysosomally degraded complex sphingolipids can be acylated to form ceramide in the ER and recycled to sphingomyelin and glycosphingolipids. (Right) Cholesterol biosynthetic enzymes reside in the ER, with some presqualene enzymes also localized in peroxisomes (Px). Sterols are transported to the plasma membrane (PM) largely via Golgi bypass route(s) and the ORP proteins play a role in this process, as well as in the reverse transport from the plasma membrane to the ER. Sterols imported into mitochondria by StAR can be used for steroid hormone synthesis. Excess cholesterol can be esterified in the ER by acyl-CoA cholesterol acyltransferase and stored in lipid droplets (LD). Arrows indicate the direction of lipid transport. Please note that arrows do not necessarily reflect the transport distance as membranes move constantly and lipid transfer may be facilitated by close apposition of membranes. Carrier proteins are indicated by black circles. Tomas Blom et al. Cold Spring Harb Perspect Biol 2011;3:a004713 ©2011 by Cold Spring Harbor Laboratory Press
 Glycerophospholipids are synthesized in the ER, with phosphatidylserine synthases enriched in mitochondria associated membrane fractions. Glycerophospholipids are transported from the ER both along exocytic membrane transport and by nonvesicular, yet poorly characterized mechanisms. (Middle) De novo sphingolipid synthesis is initiated in the ER. The ER to Golgi transport of ceramide for the assembly of more complex sphingolipids is mediated by CERT, and to a lesser extent by membrane transport. The post-Golgi transport of complex sphingolipids is dependent on membrane transport. Sphingosine that stems from lysosomally degraded complex sphingolipids can be acylated to form ceramide in the ER and recycled to sphingomyelin and glycosphingolipids. (Right) Cholesterol biosynthetic enzymes reside in the ER, with some presqualene enzymes also localized in peroxisomes (Px). Sterols are transported to the plasma membrane (PM) largely via Golgi bypass route(s) and the ORP proteins play a role in this process, as well as in the reverse transport from the plasma membrane to the ER. Sterols imported into mitochondria by StAR can be used for steroid hormone synthesis. Excess cholesterol can be esterified in the ER by acyl-CoA cholesterol acyltransferase and stored in lipid droplets (LD). Arrows indicate the direction of lipid transport. Please note that arrows do not necessarily reflect the transport distance as membranes move constantly and lipid transfer may be facilitated by close apposition of membranes. Carrier proteins are indicated by black circles. Tomas Blom et al. Cold Spring Harb Perspect Biol 2011;3:a ©2011 by Cold Spring Harbor Laboratory Press.")
97
Topologia di eventi biosintetici dei sfingolipidi nel complesso di Golgi Nel complesso Golgi la ceramide (Cer) può spontaneamente flip-floppare tra i due strati della membrana, ed è convertito in uno sfingomielina (SM) nello strato luminale, o glucosilceramide (GlcCer) nel foglietto citosolico. La sintasi luminale SM trasferisce il il gruppo di testa della fosforilcolina da fosfatidilcolina (PtdCho) alla ceramide per la generare sfingomielina e diacilglicerolo (DAG) . Anche DAG passa spontaneamente tra i due foglietti, e può essere consumata nel foglio citosolica mediante conversione in PtdCho o fosfatidiletanolamina. GlcCer sintasi aggiunge un gruppo di testa di glucosio alla ceramide nel foglietto citosolico del cis-Golgi. GlcCer deve poi traslocare al foglietto luminale per ulteriori glicosilazione producendo così un gruppo eterogeneo di glicosfingolipidi complessi (GSLs) [87]. Sebbene GlcCer può flip-flopare spontaneamente in membrane artificiali, proprio come GlcCer guadagna l'accesso alle glicosiltransferasi luminali non è chiaro
 alla ceramide per la generare sfingomielina e diacilglicerolo (DAG) . Anche DAG passa spontaneamente tra i due foglietti, e può essere consumata nel foglio citosolica mediante conversione in PtdCho o fosfatidiletanolamina. GlcCer sintasi aggiunge un gruppo di testa di glucosio alla ceramide nel foglietto citosolico del cis-Golgi. GlcCer deve poi traslocare al foglietto luminale per ulteriori glicosilazione producendo così un gruppo eterogeneo di glicosfingolipidi complessi (GSLs) [87]. Sebbene GlcCer può flip-flopare spontaneamente in membrane artificiali, proprio come GlcCer guadagna l accesso alle glicosiltransferasi luminali non è chiaro.")
Presentazioni simili











>")
 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")


 collegati a livello di membrane>")




