Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Metabolismo dei nucleotidi Citosina Uracile Timina Adenina Guanina

2
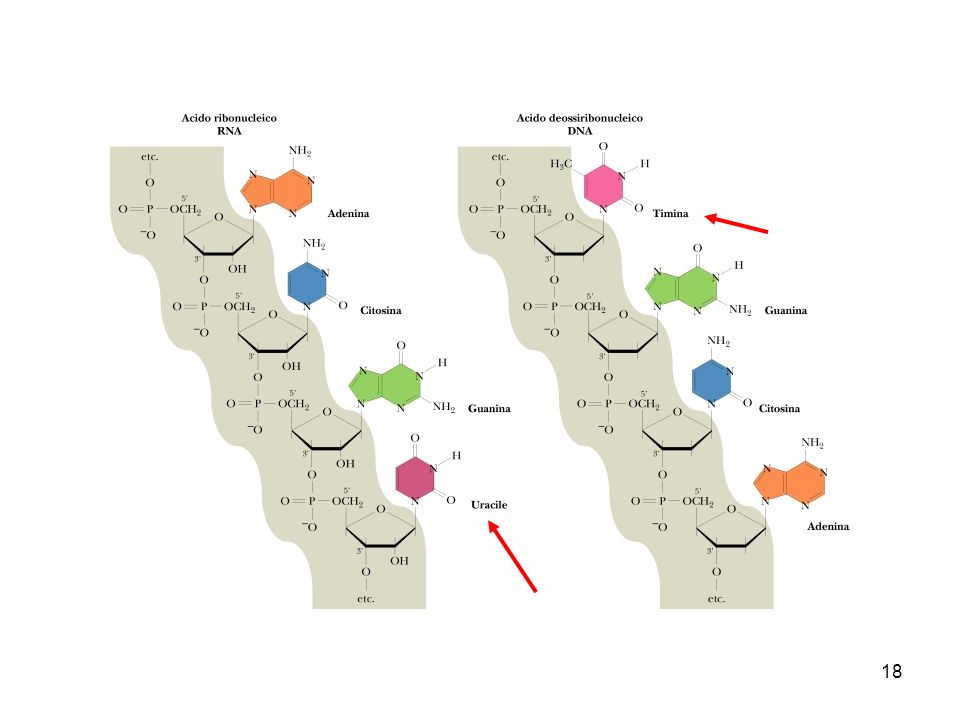
I nucleotidi sono esteri fosforici di un pentosio (ribosio o desossiribosio) in cui una base purinica o pirimidinica è legata all’atomo ci carbonio C1 dello zucchero 1 4

3
BASI PIRIMIDINICHE BASI PURINICHE

4
base + ribosio nucleoside + fosfato

5
I nucleotidi trifosfati sono precursori attivati degli acidi nucleici

6
I nucleotidi svolgono anche molte altre funzioni biologiche

7
1. Un nucleotide adeninico, l’ATP, è la “moneta” universale per gli scambi energetici nei processi biologici Anche il GTP è una fonte di energia per un numero più limitato di processi biologici

8
2. Derivati dei nucleotidi come l’UDP-glucosio partecipano a processi biosintetici come la formazione del glicogeno.

9
3. Molti nucleotidi sono componenti di cofattori fondamentali per il metabolismo. (Vit. B2) Flavina adenina dinucleotide (FAD). L’adenosina (in rosso) è legata alla riboflavina (in nero) mediante due gruppi fosforici.
. L’adenosina (in rosso) è legata alla riboflavina. (in nero) mediante due gruppi fosforici.")
10
Nicotinamide adenina dinucleotide (NAD+) e
Nicotinamide adenina dinucleotide fosfato (NADP+). Questi nucleotidi contengono adenosina legata a un nucleotide nicotinamidico. Il NADP+ contiene un ulteriore gruppo fosforico sull’ossidrile in posizione 2’ dell’adenosina. (Vit. PP)
. Questi nucleotidi contengono adenosina legata a un nucleotide nicotinamidico. Il NADP+ contiene un ulteriore gruppo fosforico sull’ossidrile in posizione 2’ dell’adenosina. (Vit. PP)")
11
4. Coenzima A (CoA). Questo derivato dell’adenosina trasporta gruppi acili uniti covalentemente mediante legami tioesteri al suo gruppo sulfidrilico.

12
5. Infine, i nucleotidi sono componenti essenziali delle vie di trasduzione del segnale. AMP ciclico e GMP ciclico sono secondi messaggeri che trasmettono segnali all’interno di una cellula. L’ATP è il donatore dei gruppi fosforici che vengono trasferiti dalle proteine chinasi.

13
A differenza degli zuccheri, degli amminoacidi e degli acidi grassi, i nucleotidi non rappresentano una fonte importante di energia metabolica

14
Aspartato e glutammina sono i donatori dei gruppi NH2 nella via di formazione dei nucleotidi.
Alcune tappe della biosintesi dei nucleotidi sono bersaglio di agenti terapeutici.

15
Tutti gli organismi possono sintetizzare i nucleotidi
sia mediante vie de novo (da precursori semplici) sia recuperando i prodotti di degradazione degli acidi nucleici
 sia recuperando i prodotti di degradazione. degli acidi nucleici.")
16
Sia le vie de novo sia le vie di recupero producono ribonucleotidi.
Il DNA viene costruito utilizzando come precursori deossiribonucleotidi. Tutti i deossiribonucleotidi vengono sintetizzati a partire dai corrispondenti ribonucleotidi prodotto dalla riduzione del ribosio

17
BASI PIRIMIDINICHE BASI PURINICHE

19
Sintesi dei ribonucleotidi pirimidinici

20
La via de novo per la sintesi dei nucleotidi pirimidinici
(proveniente dalla glutammina) sintesi de novo dell’anello pirimidinico legame dell’anello al ribosio e formazione di un nucleotide pirimidinico.
 sintesi de novo dell’anello. pirimidinico. legame dell’anello al ribosio e. formazione di un nucleotide. pirimidinico.")
21
La prima tappa della biosintesi de novo delle pirimidine è la
Sintesi del carbamilfosfato CPS II 2 ATP + HCO3- + glutammina + H2O H2N – C –OPO ADP + Pi + glutammato O CPS II -sintesi pirimidine -enzima citosolico -utilizza glutammina -non necessita di N-acetil glutammato CPS I -ciclo dell’urea enzima mitocondriale utilizza ammoniaca come donatote di azoto richiede N-acetil glutammato

22
Struttura della carbamilfosfato sintetasi
Meccanismo d’azione della CPS [1] [2] [3] Struttura della carbamilfosfato sintetasi 2 catene polipeptidiche: -una più piccola (giallo) -una più grande con due ripiegamenti (blu e rosso) siti di legame dell’ATP
 -una più grande con due ripiegamenti (blu e rosso) siti di legame dell’ATP.")
23
forma attivata

24
Incanalamento del substrato i tre siti attivi della CPS sono uniti
da un canale attraverso il quale passano gli intermedi. carbossi-P 2 funzioni: gli intermedi restano associati all’enzima gli intermedi labili (carbonilfosfato e acido carbammico) sono protetti dall’idrolisi.
 sono protetti dall’idrolisi.")
25
Via di biosintesi de novo dei nucleotidi pirimidinici
enzima mitocondriale connesso alla catena di trasporto degli elettroni

26
le prime 3 attività enzimatiche si sono
D CAD e UMP sintasi le prime 3 attività enzimatiche si sono fuse insieme sottoforma di una singola proteina multifunzionale codificata da un singolo gene. le ultime 2 attività enzimatiche (orotato fosforibosil trasferasi e OMP decarbossilasi) sono presenti sottoforma di una singola proteina multifunzionale codificata da un singolo gene. La mancanza di UMP sintasi provoca orotico aciduria.
 sono presenti. sottoforma di una singola. proteina multifunzionale codificata da. un singolo gene. La mancanza di UMP sintasi provoca. orotico aciduria.")
27
VANTAGGIO FUNZIONALE DELLE PROTEINE MULTIENZIMATICHE
- i prodotti della prima reazione rimangono legati all’enzima e possono essere convogliati sul secondo sito attivo questo processo evita la diffusione degli intermedi metabolici nell’ambiente intracellulare migliore efficienza metabolica dei singoli passaggi.

28
I nucleosidi difosfato e trifosfato sono tra loro interconvertibili
ad opera dell’enzima nucleoside difosfato chinasi UMP + ATP UDP + ADP XDP + YTP XTP + YDP

29
Sintesi di CTP da UTP UMP
L’enzima CTP sintetasi catalizza la reazione di amminazione in posizione 4 dell’anello pirimidinico dell’UTP, per formare CTP. Negli eucarioti il gruppo amminico deriva dalla glutammina.

30
* La ragione di questo processo sta nella necessità di mantenere molto bassa la concentrazione di dUTP per impedire che il dUTP possa fungere da substrato per la sintesi del DNA * (d) (d) (d)
 (d) (d)")
31
(metabolismo cellulare)
Inibizione della sintesi del timidilato nella terapia del cancro FdUMP= substrato suicida Fluoro-uridina (Adrucin, Efudex, Fluracil) FdUMP (metabolismo cellulare) L’enzima timidilato sintasi resta congelato sotto forma di complesso ternario covalente enzima-FdUMP-THF
 FdUMP. (metabolismo cellulare) L’enzima timidilato sintasi resta congelato. sotto forma di complesso ternario covalente. enzima-FdUMP-THF.")
32
gruppo metilico CH3 gruppo metilenico CH2

33
La reazione della timidilato sintasi rappresenta un processo biochimico unico in sé, per il fatto che il THF viene ossidato a DHF; nessun’altra reazione che utilizza il THF come cofattore ne modifica lo stato di ossidazione

34
Il blocco della produzione di dTMP sta alla base di alcune forme di
Rigenerazione dell’N5, N10-metilenetetraidrofolato (timidilato) Il blocco della produzione di dTMP sta alla base di alcune forme di chemioterapia anticancro (analoghi del DHF) antifolati
 Il blocco della produzione di dTMP sta alla base di alcune forme di. chemioterapia anticancro. (analoghi del DHF) antifolati.")
35
Altre cellule in attiva proliferazione: cellule del midollo spinale
Un’interruzione della sintesi del dTMP uccide le cellule in attiva proliferazione, come le cellule cancerose. Altre cellule in attiva proliferazione: cellule del midollo spinale cellule dei follicoli dei capelli cellule della mucosa intestinale cell. del sangue cell. del sistema immunitario

36
REGOLAZIONE DELLA BIOSINTESI DEI NUCLEOTIDI PIRIMIDINICI

37
carbamilfosfato sintetasi La proteina multimerica CAD viene attivata mediante fosforilazione da parte di: mytogen-activated protein chinasi (MAPK) proteina chinasi A (PKA) CAD è degradata dalla proteolisi operata dalla caspasi 3 durante il fenomeno apoptotico. OMP decarbossilasi
 proteina chinasi A (PKA) CAD è degradata dalla proteolisi operata dalla caspasi 3 durante il fenomeno apoptotico. OMP decarbossilasi.")
38
Vie di salvataggio delle pirimidine
uracile + PRPP UMP + PPi uracil fosforibosil transferasi (UPRTasi) timidina + ATP TMP + ADP conversione di un nucleoside in nucleotide attraverso una reazione di fosforilazione diretta (via di salvataggio specifica per la timidina).
 timidina + ATP. TMP + ADP. conversione di un nucleoside in nucleotide attraverso una reazione di fosforilazione diretta (via di salvataggio specifica per la timidina).")
39
Le principali vie cataboliche dei nucleotidi pirimidinici
I prodotti finali del catabolismo delle pirimidine b alanina e acido b amminoisobutirrato sono amminoacidi e vengono metabolizzati come tali. Il malonil CoA è un precursore della biosintesi degli acidi grassi e il metilmalonil CoA è convertito nell’intermedio del ciclo dell’acido citrico succinil CoA. Anche se in quantità limitata forniscono precursori al metabolismo energetico

Presentazioni simili