Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
STRUTTURA DELLA PARETE DEI BATTERI

2
a | Assembly of the cell-wall subunit takes place on the cytosolic side of the bacterial plasma membrane. UDP-activated precursor sugars are assembled on a polyisoprenoid carrier, the coupling of which produces Lipid II, which is then transported across the membrane by an unknown mechanism. Next, the peptidoglycan subunit is transferred to the growing peptidoglycan chain and the polyisoprenoid carrier is recycled back to the cytoplasmic side to complete the cycle. b | Structure of Lipid II. The polyisoprenoid anchor consists of eight isoprene units in the cis-conformation followed by two units in the trans-conformation and the terminal isoprene unit. Depicted here is a Lipid II carrying a pentapeptide with a lysine at the third position. Together with diaminopimelic acid, this is the most common residue at this position. The third amino acid of the pentapeptide is coupled to the glutamate at position two via the side chain carboxylate. The red bars indicate the minimal binding sites in Lipid II of glycopeptide antibiotics (1), nisin (2), ramoplanin (3) and mersacidin (4). GlcNAc, N-acetylglucosamine; MurNAc, N-acetylmuramic acid.
, nisin (2), ramoplanin (3) and mersacidin (4). GlcNAc, N-acetylglucosamine; MurNAc, N-acetylmuramic acid..")
3
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina

4
Sintesi dei monomeri di mureina
UDP-Acido N-acetilmuramico L-Ala D-Glu L-Lys D-Ala D-Ala Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan PEPTIDE DI PARK
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan. PEPTIDE DI PARK.")
5
L-Ala D-Glu L-Lys Gly D-Ala UDP-Acido N-acetilmuramico

6
Sintesi dei monomeri di mureina
BP-NAM-NAG Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan.")
7
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina

8
Traslocazione dei monomeri di mureina
Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan.")
9
ANTIBIOTICI PEPTIDICI

10
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina Formazione di polimeri lineari di mureina (transglicosilazione) NAM Gly5 NAG NAM Gly5 NAG NAM Gly5 NAG NAM Gly5 NAG
 NAM. Gly5. NAG. NAM. Gly5. NAG. NAM. Gly5. NAG. NAM. Gly5. NAG.")
11
ANTIBIOTICI GLICOPEPTIDICI
Dhpg, dihydroxyphenylglycine; Glc, glucose; Hpg, hydroxyphenylglycine; Van, vancosamine.

12
Gly ANTIBIOTICI GLICOPEPTIDICI Glicopeptide polimero
L-Ala D-Glu L-Lys Gly D-Ala Acido N-acetilmuramico Glicopeptide polimero Acido N-acetilmuramico Glicopeptide polimero ANTIBIOTICI GLICOPEPTIDICI Acido N-acetilmuramico Glicopeptide polimero

13
The VanA gene cluster that confers vancomycin resistance
The VanA gene cluster that confers vancomycin resistance. A TWO-COMPONENT REGULATORY system VanR–VanS regulates vancomycin resistance in vancomycin-resistant ENTEROCOCCI (VRE) and vancomycin-resistant Staphylococcus aureus (VRSA) strains. VanS is a membrane-associated sensor (of vancomycin) that controls the level of phosphorylation of VanR. VanR is a transcriptional activator of the operon encoding VanH, VanA and VanX. VanH is a dehydrogenase that reduces pyruvate to D-Lac, whereas VanA is a ligase that catalyses the formation of an ester bond between D-Ala and D-Lac. Vancomycin does not bind to D-Ala-D-Lac, which leads to vancomycin resistance. VanX is a dipeptidase that hydrolyses the normal peptidoglycan component D-Ala-D-Ala, which prevents it from causing vancomycin sensitivity. VanY is a D,D-carboxypeptidase that hydrolyses the terminal D-Ala residue of late peptidoglycan precursors that are produced if elimination of D-Ala-D-Ala by VanX is not complete. So, D-Ala-D-Lac replaces the normal dipeptide D-Ala-D-Ala in peptidoglycan synthesis resulting in vancomycin resistance150. VanZ confers resistance to teicoplanin by an unknown mechanism.
 and vancomycin-resistant Staphylococcus aureus (VRSA) strains. VanS is a membrane-associated sensor (of vancomycin) that controls the level of phosphorylation of VanR. VanR is a transcriptional activator of the operon encoding VanH, VanA and VanX. VanH is a dehydrogenase that reduces pyruvate to D-Lac, whereas VanA is a ligase that catalyses the formation of an ester bond between D-Ala and D-Lac. Vancomycin does not bind to D-Ala-D-Lac, which leads to vancomycin resistance. VanX is a dipeptidase that hydrolyses the normal peptidoglycan component D-Ala-D-Ala, which prevents it from causing vancomycin sensitivity. VanY is a D,D-carboxypeptidase that hydrolyses the terminal D-Ala residue of late peptidoglycan precursors that are produced if elimination of D-Ala-D-Ala by VanX is not complete. So, D-Ala-D-Lac replaces the normal dipeptide D-Ala-D-Ala in peptidoglycan synthesis resulting in vancomycin resistance150. VanZ confers resistance to teicoplanin by an unknown mechanism.")
14
LANTIBIOTICI DI TIPO A (a catena lineare)
For all antibiotics shown, d-amino acids are colour-coded red. In the structure of nisin, the five lanthionine rings are labelled a–e from the amino terminus. Abu, aminobutyric acid; Dha, didehydroalanines; Dhb, didehydrobutyrines;

15
First, nisin reaches the bacterial plasma membrane (a), where it binds to Lipid II via two of its amino-terminal rings (b). This is then followed by pore formation (c), which involves a stable transmembrane orientation of nisin. During or after assembly of four 1:1 (nisin: Lipid II) complexes, four additional nisin molecules are recruited to form the pore complex (d).
, which involves a stable transmembrane orientation of nisin. During or after assembly of four 1:1 (nisin: Lipid II) complexes, four additional nisin molecules are recruited to form the pore complex (d)..")
16
LANTIBIOTICI DI TIPO B (a struttura globulare)
For all antibiotics shown, d-amino acids are colour-coded red. In the structure of nisin, the five lanthionine rings are labelled a–e from the amino terminus. In the structure of mersacidin, the arrowhead indicates the location of the hinge region that was revealed by NMR. Abu, aminobutyric acid; Aiha, -amino- -[4'-(2'-iminoimidazolidinyl)]- -hydroxypropionic acid; aThr, allo-threonine; Chp, 3-chloro-4-hydroxyphenylglycine; Dha, didehydroalanines; Dhb, didehydrobutyrines; Dhpg, dihydroxyphenylglycine; Glc, glucose; HAsn, -hydroxyasparagine; HAsp, -hydroxy-aspartic acid; Hleu, -hydroxyleucine; Hpg, hydroxyphenylglycine; HPro, 3-hydroxy-proline; Man, mannose; Me-phe, -methylphenylalanine; Orn, ornithine; PhSer, -phenylserine; Van, vancosamine.
]- -hydroxypropionic acid; aThr, allo-threonine; Chp, 3-chloro-4-hydroxyphenylglycine; Dha, didehydroalanines; Dhb, didehydrobutyrines; Dhpg, dihydroxyphenylglycine; Glc, glucose; HAsn, -hydroxyasparagine; HAsp, -hydroxy-aspartic acid; Hleu, -hydroxyleucine; Hpg, hydroxyphenylglycine; HPro, 3-hydroxy-proline; Man, mannose; Me-phe, -methylphenylalanine; Orn, ornithine; PhSer, -phenylserine; Van, vancosamine.")
17
RAMOPLANINA For all antibiotics shown, d-amino acids are colour-coded red. In the structure of nisin, the five lanthionine rings are labelled a–e from the amino terminus. In the structure of mersacidin, the arrowhead indicates the location of the hinge region that was revealed by NMR. Abu, aminobutyric acid; Aiha, -amino- -[4'-(2'-iminoimidazolidinyl)]- -hydroxypropionic acid; aThr, allo-threonine; Chp, 3-chloro-4-hydroxyphenylglycine; Dha, didehydroalanines; Dhb, didehydrobutyrines; Dhpg, dihydroxyphenylglycine; Glc, glucose; HAsn, -hydroxyasparagine; HAsp, -hydroxy-aspartic acid; Hleu, -hydroxyleucine; Hpg, hydroxyphenylglycine; HPro, 3-hydroxy-proline; Man, mannose; Me-phe, -methylphenylalanine; Orn, ornithine; PhSer, -phenylserine; Van, vancosamine.
]- -hydroxypropionic acid; aThr, allo-threonine; Chp, 3-chloro-4-hydroxyphenylglycine; Dha, didehydroalanines; Dhb, didehydrobutyrines; Dhpg, dihydroxyphenylglycine; Glc, glucose; HAsn, -hydroxyasparagine; HAsp, -hydroxy-aspartic acid; Hleu, -hydroxyleucine; Hpg, hydroxyphenylglycine; HPro, 3-hydroxy-proline; Man, mannose; Me-phe, -methylphenylalanine; Orn, ornithine; PhSer, -phenylserine; Van, vancosamine.")
18
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina Formazione di polimeri lineari di mureina (transglicosilazione) Transpeptidazione (formazione di legami crociati tra polimeri lineari)
 Transpeptidazione (formazione di legami crociati tra polimeri lineari)")
19
Gly Glicopeptide polimero Acido N-acetilmuramico Glicopeptide polimero
L-Ala D-Glu L-Lys Gly D-Ala Transpeptidasi Acido N-acetilmuramico Glicopeptide polimero

20
Gly Antibiotici -lattamici Glicopeptide polimero
L-Ala D-Glu L-Lys Gly D-Ala Acido N-acetilmuramico Glicopeptide polimero Acido N-acetilmuramico Glicopeptide polimero Antibiotici -lattamici Transpeptidasi Acido N-acetilmuramico Glicopeptide polimero

21
STRUTTURA DEGLI ANTIBIOTICI -LATTAMICI

22
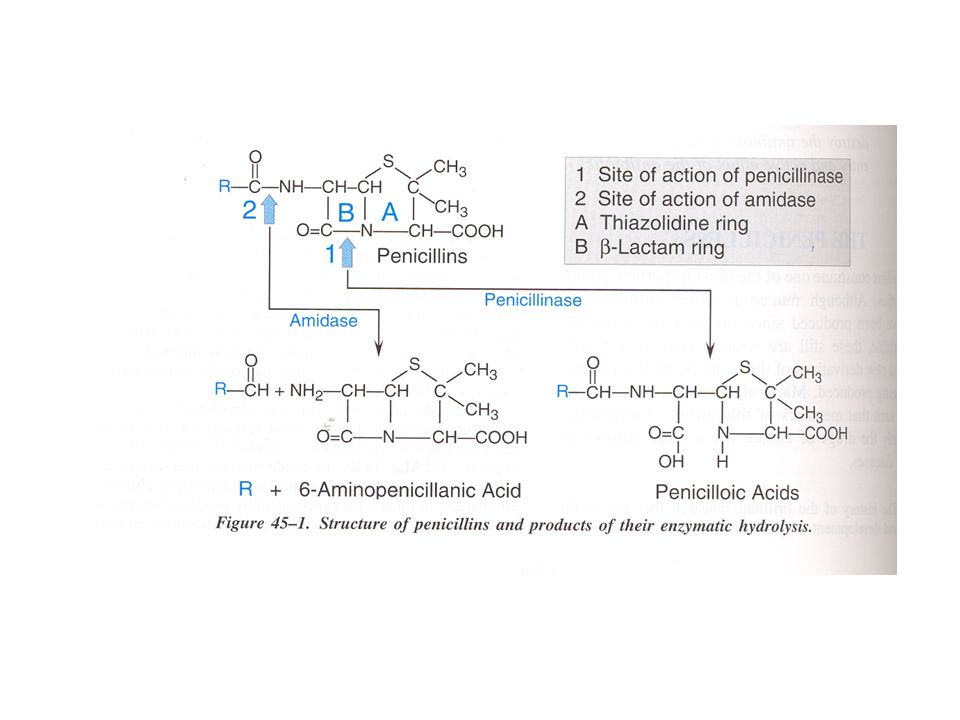
CLASSIFICAZIONE DELLE PENICILLINE: 1 - PENICILLINE NATURALI
ANELLO -LATTAMICO ANELLO TIAZOLIDNICO ACIDO 6-AMINOPENICILLANICO

23
Meccanismi di secrezione degli acidi (A) e delle
basi organiche (B) nel tubulo prossimale
 nel tubulo prossimale.")
24
Meccanismo d’azione degli antibiotici -lattamici
Interazione con le penicillin-binding proteins (PBPs) Inibizione della reazione di transpeptidazione Inattivazione di un inibitore di enzimi autolitici della parete cellulare (p.es. mureina idrolasi) Distruzione enzimatica del farmaco da parte di amido- idrolasi (poco efficaci), ma soprattutto di -lattamasi
 Inibizione della reazione di transpeptidazione. Inattivazione di un inibitore di enzimi autolitici della. parete cellulare (p.es. mureina idrolasi) Distruzione enzimatica del farmaco da parte di amido- idrolasi (poco efficaci), ma soprattutto di -lattamasi.")
26
CLASSIFICAZIONE DELLE PENICILLINE: 2 -PENICILLINE ANTI-STAFILOCOCCICHE
(resistenti alla -lattamasi)
")
27
CLASSIFICAZIONE DELLE PENICILLINE: 3 - PENICILLINE AD AMPIO SPETTRO

28
Pseudomonas aeruginosa: struttura e
fattori di virulenza

29
Colonization Chronic infection Acute infection
break in front line defenses adhesion thanks to cell-associated virulence factors Chronic infection in cystic fibrosis patients selection of alginate-producing mitants protects from host defense mechanisms low production of extracellular virulence factors tissue damage due to chronic inflammation Acute infection Cell-to-cell signaling systems allow coordinate cell density-dependent production of high amounts of extracellular virulence factors tissue damage by protease and…, leading to blood vessel invasion, diseemination, systemic inflammatory response syndrome, multiple organ failure and death

31
CLASSIFICAZIONE DELLE PENICILLINE: 4 - PENICILLINE A SPETTRO ESTESO
(attive contro Pseudomonas)
")
32
Accessory protein

33
Classificazione delle -lattamasi
Classe A: comprende la maggior parte delle forme di rilevanza clinica, generalmente codificate da plasmidi e sensibili agli inibitori attualmente in commercio; contengono un residuo di serina a livello del sito attivo Classe B: sono metalloenzimi, contenenti un atomo di Zn a livello del sito attivo e sono quelle con la più ampia specificità di substrato; sono insensibili agli inibitori attualmente in commercio Classe C: sono generalmente codificate da DNA cromosomiale e attive nei confronti delle cefalosporine, che sono in grado di indurne l’espressione attraverso derepressione genica; contengono un residuo di serina a livello del sito attivo Classe D: sono attive principalmente nei confronti delle oxacilline; contengono un residuo di serina a livello del sito attivo

34
Spettro d’azione delle -lattamasi

35
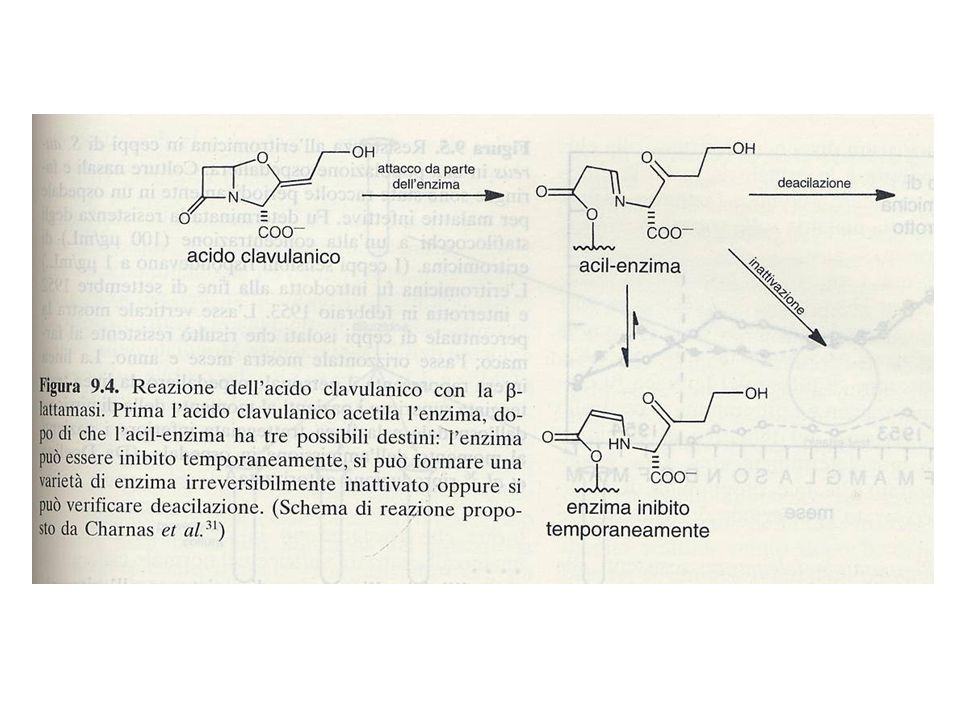
Inibitori delle -lattamasi

37
CEFALOSPORINE COOH ANELLO -LATTAMICO ANELLO DIIDROTIAZINICO
ACIDO 7-AMINOCEFALOSPORANICO

38
CEFALOSPORINE I GENERAZIONE Attività vs Resistenza alle Gram + Gram -
-lattamasi I GENERAZIONE (es. cefalotina, cefazolina, cefalexina) II GENERAZIONE (es. cefamandolo, cefoxitina, cefuroxima) III GENERAZIONE (es. cefotaxime, ceftazidime, cefoperazone) IV GENERAZIONE (es. cefepime, cefpirome)
 II GENERAZIONE. (es. cefamandolo, cefoxitina, cefuroxima) III GENERAZIONE. (es. cefotaxime, ceftazidime, cefoperazone) IV GENERAZIONE. (es. cefepime, cefpirome)")
39
ANTIBIOTICI MONOBATTAMICI
SO3H O

40
ANTIBIOTICI CARBAPENEMICI

41
IMIPENEM DEGRADAZIONE DIPEPTIDASI CILASTATINA

42
Classificazione delle -lattamasi
Classe A: comprende la maggior parte delle forme di rilevanza clinica, generalmente codificate da plasmidi e sensibili agli inibitori attualmente in commercio; contengono un residuo di serina a livello del sito attivo Classe B: sono metalloenzimi, contenenti un atomo di Zn a livello del sito attivo e sono quelle con la più ampia specificità di substrato; sono insensibili agli inibitori attualmente in commercio Classe C: sono generalmente codificate da DNA cromosomiale e attive nei confronti delle cefalosporine, che sono in grado di indurne l’espressione attraverso derepressione genica; contengono un residuo di serina a livello del sito attivo Classe D: sono attive principalmente nei confronti delle oxacilline; contengono un residuo di serina a livello del sito attivo

43
Spettro d’azione delle -lattamasi

44
Beta-lactam toxicity. UPPER PANEL: Beta-lactams can modify amino groups on human proteins, creating an immunogenic beta-lactam hapten. LOWER PANEL: in the absence of modification, human proteins are nonantigenic. Modification of endogenous proteins by the addition of a beta-lactam antibiotic results in the formation of a new antigenic determinant that can be recognized as “foreign” by antibodies of the host immune system

45
ANTIBIOTICI PEPTIDICI
POLIMIXINA B The polymyxins (Polymixin B and colistin) are basic peptides that act as cationic detergents. Their antimicrobial activity is restricted to Gram-negative bacteria. They are surface active, amphipathic agents that interact strongly with phospholipids and penatrate into and disrupt the structure of cell membranes.
 are basic peptides that act as cationic detergents. Their antimicrobial activity is restricted to Gram-negative bacteria. They are surface active, amphipathic agents that interact strongly with phospholipids and penatrate into and disrupt the structure of cell membranes.")
46
LIPOPEPTIDI CICLICI Daptomycin is an antibacterial agent of a new calss of antibiotics, the cyclic lipopeptides. Daptomycin is a natural product which has clinical utility in the treatment of infections caused by aerobic Gram-positive bacteria. The mechanism of action of daptomycin is distincts from any other antibiotic. Daptomycin binds to baterial membranes and causes a rapid depolarization of membrane potential. The loss of membrane potential leads to inhibition of protein, DNA and RNA synthesis, which results in bacterial cell death. FDA Drug Approva List (cited 29 October 2003)
")
Presentazioni simili

















>")


