Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

3
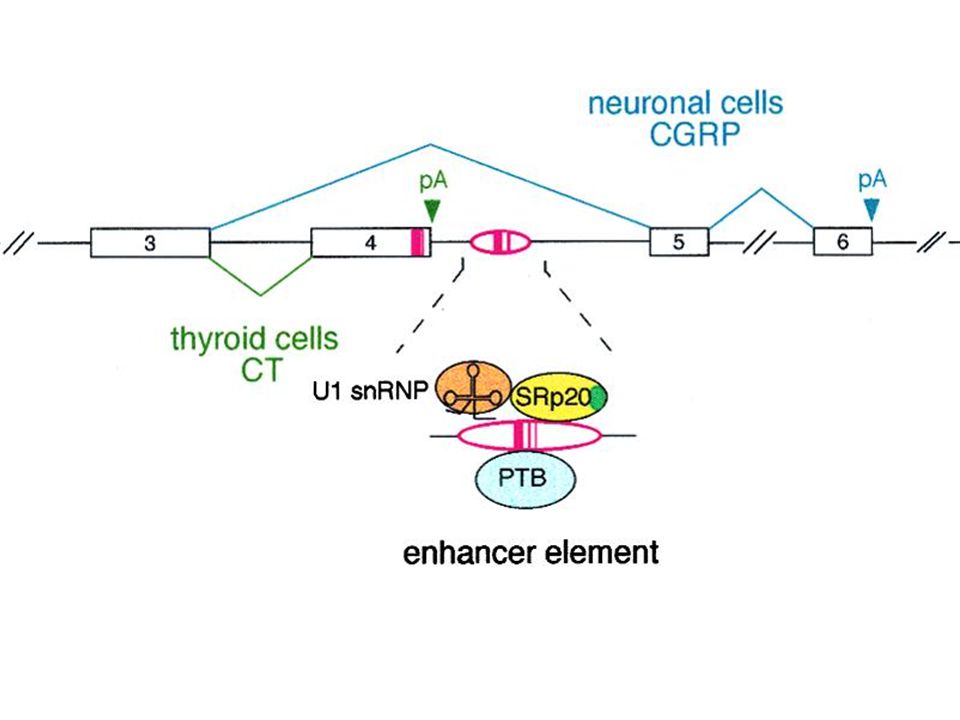
Scelta del sito di poliadenilazione
Sequenze cis-agenti che definiscono il sito di taglio Concentrazione o attività dei fattori costitutivi di poliadenilazione Espressione di fattori specifici di poliadenilazione e fattori di splicing

9
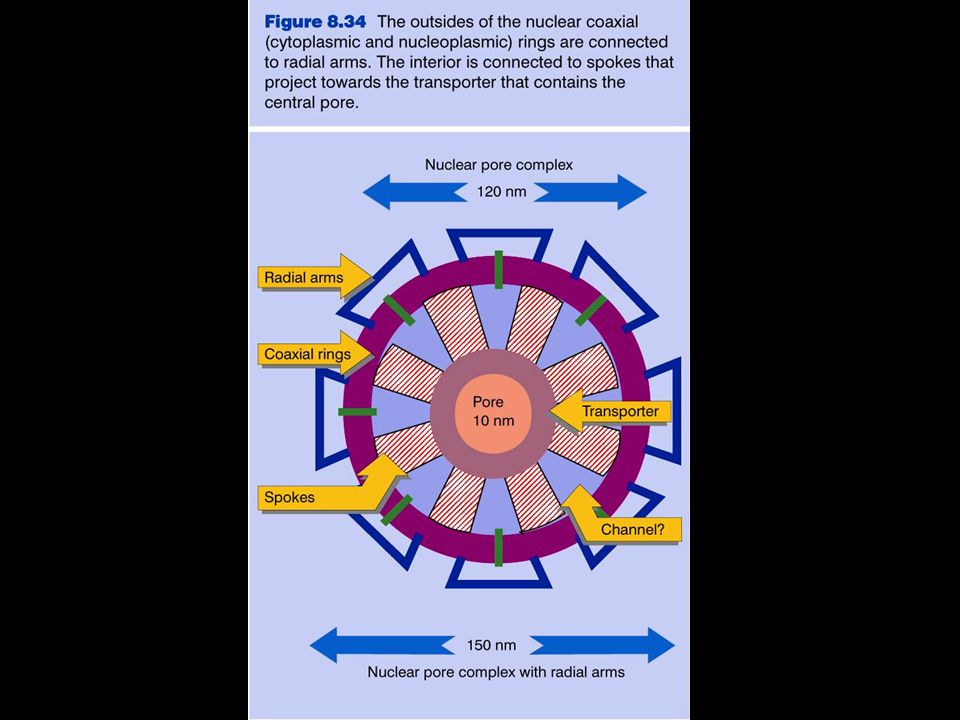
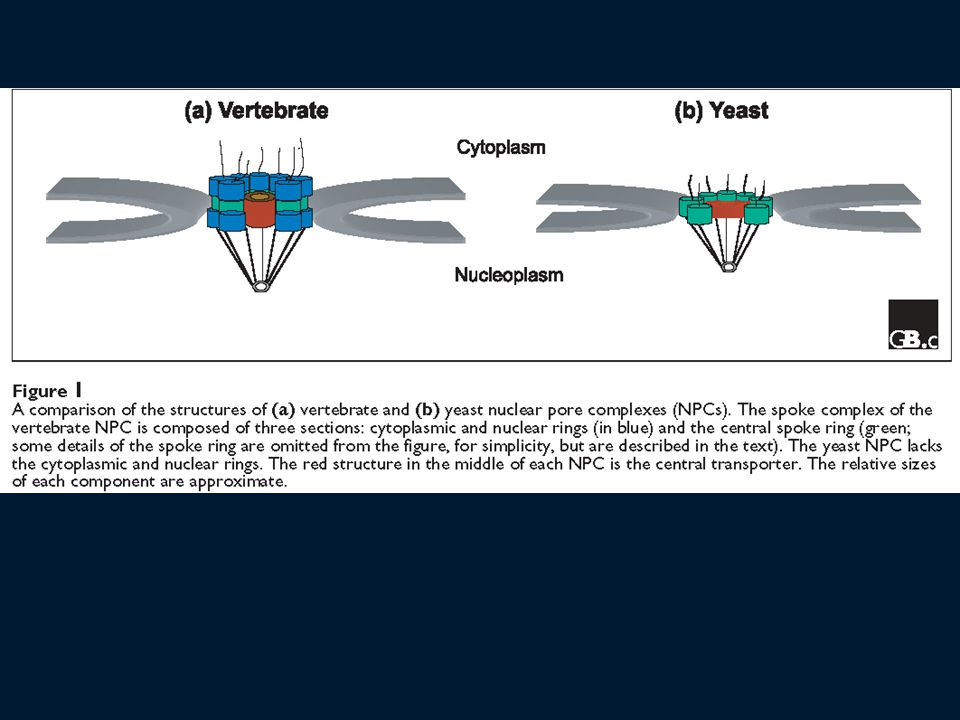
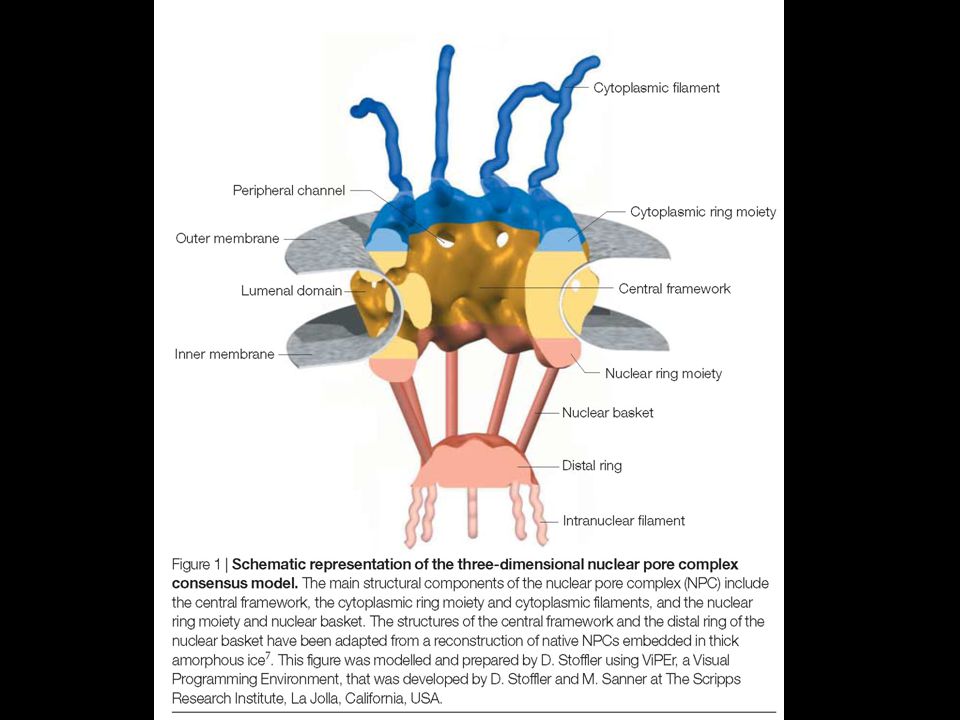
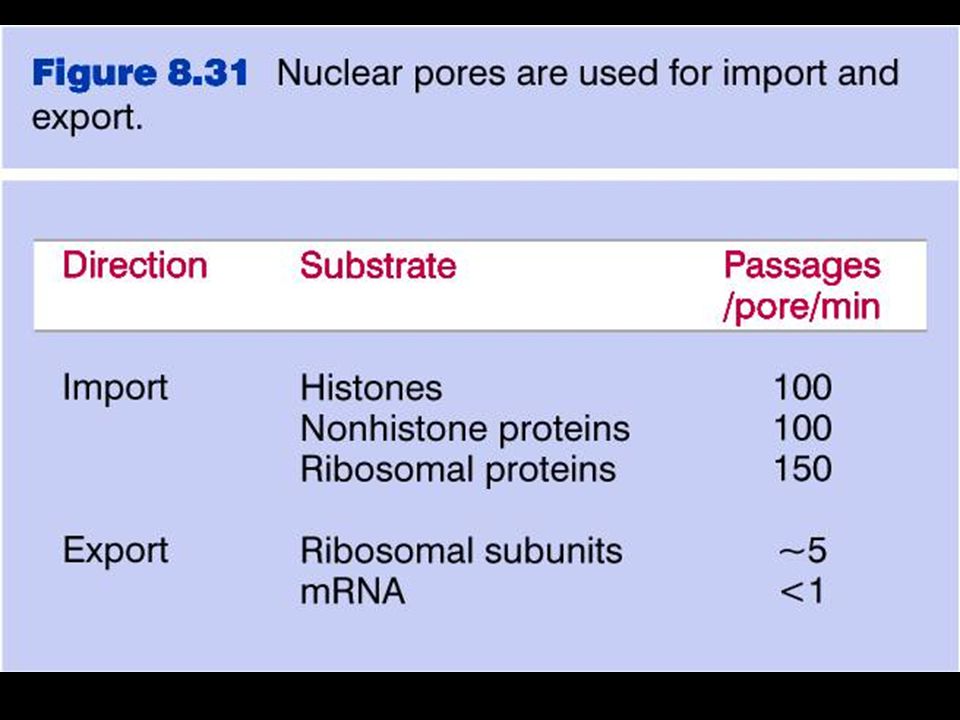
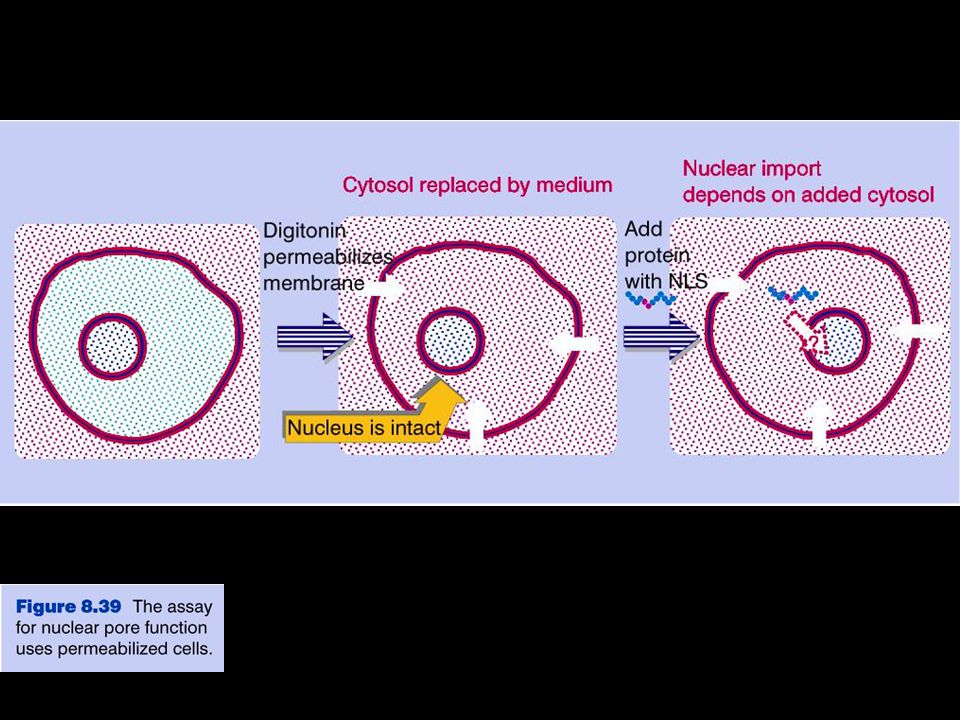
Poro nucleare Il complesso del poro nucleare (NPC) è grande 125 MDa (66 MDa in lievito) 2000 NPC nei vertebrati (200 in lievito) NPC è composto da circa 1000 proteine di tipi diversi (di ciascuna almeno 8 copie) Molecole fino a 9 nm (30-40 kDa) diffondono liberamente attraverso l’NPC Molecole fino a 25 nm vengono attivamente trasportate attraverso l’NPC
 Molecole fino a 9 nm (30-40 kDa) diffondono liberamente attraverso l’NPC. Molecole fino a 25 nm vengono attivamente trasportate attraverso l’NPC.")
11
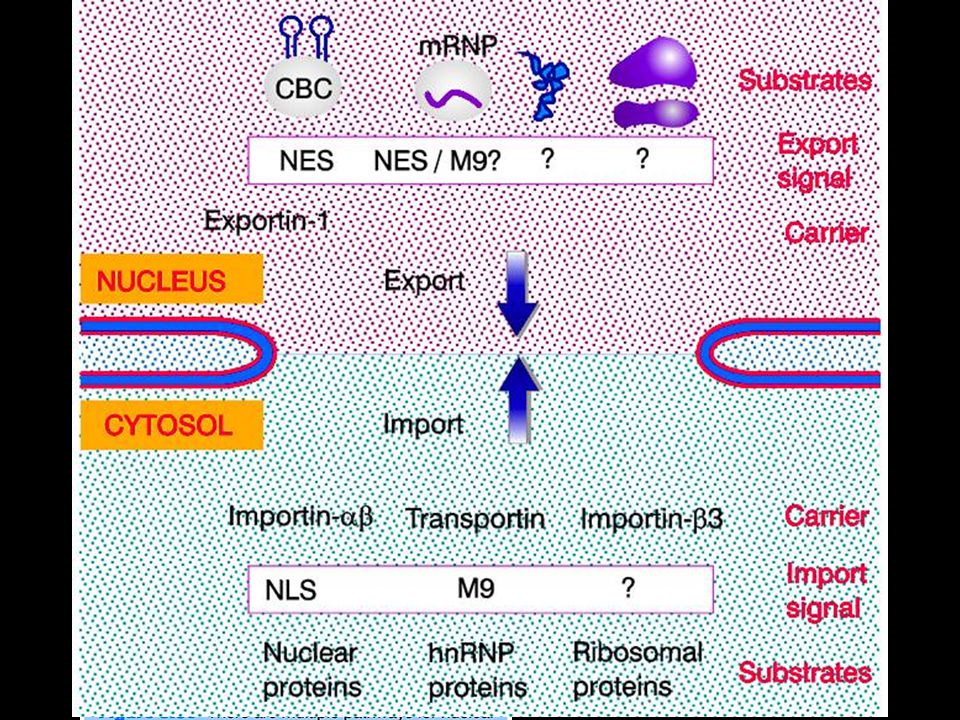
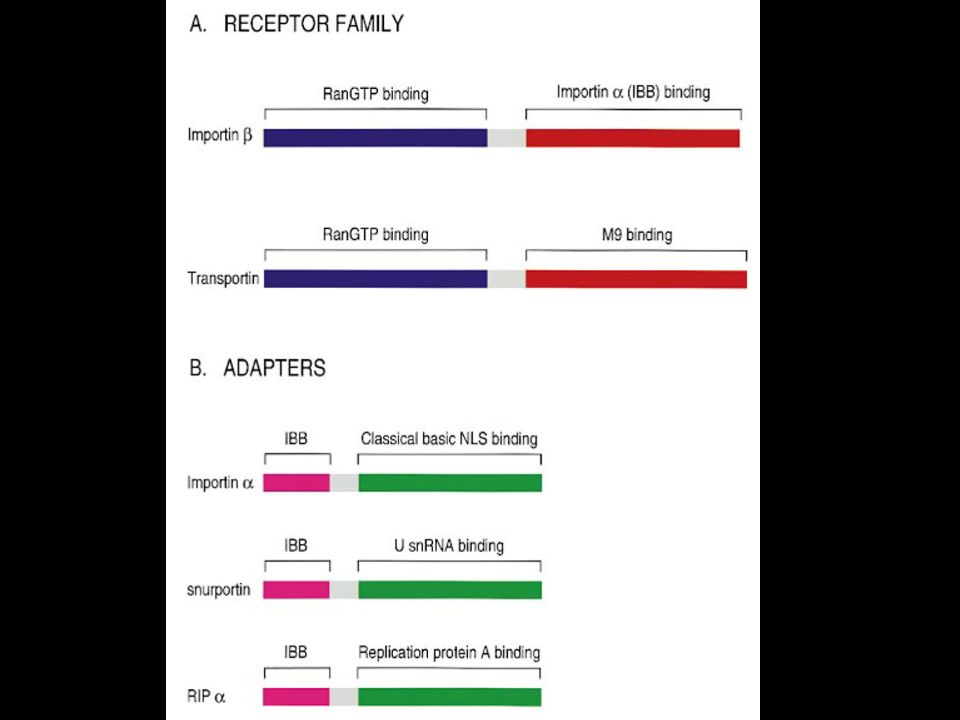
Substrato Carrier

20
Nuclear export of mRNA. mRNAs undergo several ordered processing steps before export to the cytoplasm. These maturation events are important for correct packaging of the mRNA. A key step is the deposition of the exon–exon junction complex (EJC) onto the mRNA. The EJC consists of multiple mRNA binding proteins including Y14, RNPS1, SRm160, DEK, Mago and Upf3 [39–48]. The EJC is dynamic and some components remain associated with the mRNA even after export to cytoplasm (EJC*). The EJC therefore links mRNA processing and transport to cytoplasmic events such as cytoplasmic localization and nonsense-mediated decay (NMD). In NMD, a premature stop codon is recognised if it is upstream of a splice junction. The EJC* is able to communicate the relative position of the splice junction to the NMD machinery. A translating ribosome encountering a stop codon is depicted in grey. An alternative exit route was described for ARE-containing mRNAs [64]. ARE-containing mRNAs can gain access to transportin 2 (Trn2) or Crm1 via the ARE-interacting protein HuR, either directly or through the additional adaptors pp32/April.
 onto the mRNA. The EJC consists of multiple mRNA binding proteins including Y14, RNPS1, SRm160, DEK, Mago and Upf3 [39–48]. The EJC is dynamic and some components remain associated with the mRNA even after export to cytoplasm (EJC*). The EJC therefore links mRNA processing and transport to cytoplasmic events such as cytoplasmic localization and nonsense-mediated decay (NMD). In NMD, a premature stop codon is recognised if it is upstream of a splice junction. The EJC* is able to communicate the relative position of the splice junction to the NMD machinery. A translating ribosome encountering a stop codon is depicted in grey. An alternative exit route was described for ARE-containing mRNAs [64]. ARE-containing mRNAs can gain access to transportin 2 (Trn2) or Crm1 via the ARE-interacting protein HuR, either directly or through the additional adaptors pp32/April..")
23
Figure 3. The Ran GTPase Cycle
Ran is maintained as RanGTP in the nucleus by the activity of the Ran GTP-GDP exchange factor (RanGEF or RCC1) and as RanGDPin the cytoplasm by the RanGTPase activating protein (RanGAP). RanBP1 in the cytoplasm and RanBP1-like domains on the cytoplasmic fibrils of the NPC are coactivators of RanGAP (not shown). The structures of RanGDP, RanGEF, RanGAP and a RanGTP-RanBP1 domain complex have been solved, providing structural explanations for some of the biochemical properties of Ran and its regulators, and highlighting the importance of the C-terminal extension of Ran, which is unique among small GTPases. This C-terminal region functions as a novel molecular switch (Macara, 1999).
 and as RanGDPin the cytoplasm by the RanGTPase activating protein (RanGAP). RanBP1 in the cytoplasm and RanBP1-like domains on the cytoplasmic fibrils of the NPC are coactivators of RanGAP (not shown). The structures of RanGDP, RanGEF, RanGAP and a RanGTP-RanBP1 domain complex have been solved, providing structural explanations for some of the biochemical properties of Ran and its regulators, and highlighting the importance of the C-terminal extension of Ran, which is unique among small GTPases. This C-terminal region functions as a novel molecular switch (Macara, 1999).")
24
Figure 4. The Differential Effects of RanGTP on Nuclear Import and Nuclear Export Receptor- Cargo Complexes Import receptors bind their cargos in a RanGTP-independent manner and RanGTP causes dissociation of these complexes. They are thus permitted to form in the cytoplasm and dissociate in the RanGTP-rich nucleus. Export receptors form stable complexes with their cargos only in the presence of RanGTP. These ternary complexes are thought to be the export unit, and dissociate in the cytoplasm and/or on the cytoplasmic filaments of the pore where RanGAP activity converts the RanGTP to RanGDP. Crystal structures of import receptors bound to cargo (importin b–IBB complex) and bound to Ran (transportin-RanGppNHp and importin b–RanGppNHp complexes) suggest how Ran may mediate cargo unloading (Macara, 1999; Mattaj and Conti, 1999). Ran contacts an acidic loop in the central region of the receptor molecules, and this interaction is probably responsible, at least in part, for cargo displacement. Structures of export receptors have not yet been reported.
 and bound to Ran (transportin-RanGppNHp and importin b–RanGppNHp complexes) suggest how Ran may mediate cargo unloading (Macara, 1999; Mattaj and Conti, 1999). Ran contacts an acidic loop in the central region of the receptor molecules, and this interaction is probably responsible, at least in part, for cargo displacement. Structures of export receptors have not yet been reported..")
25
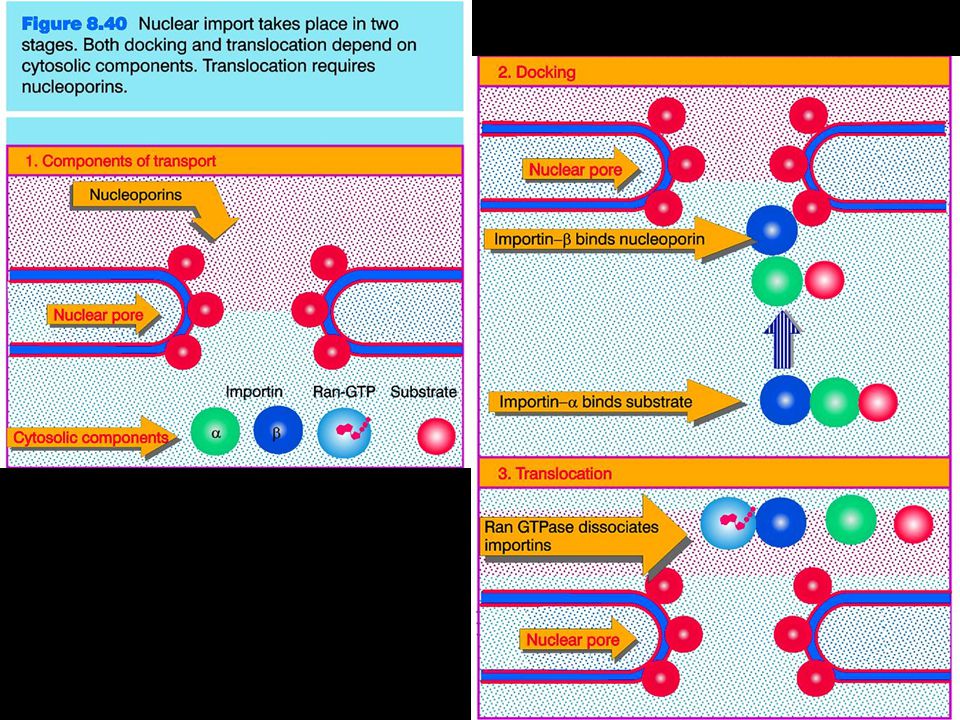
Transport through the nuclear pores
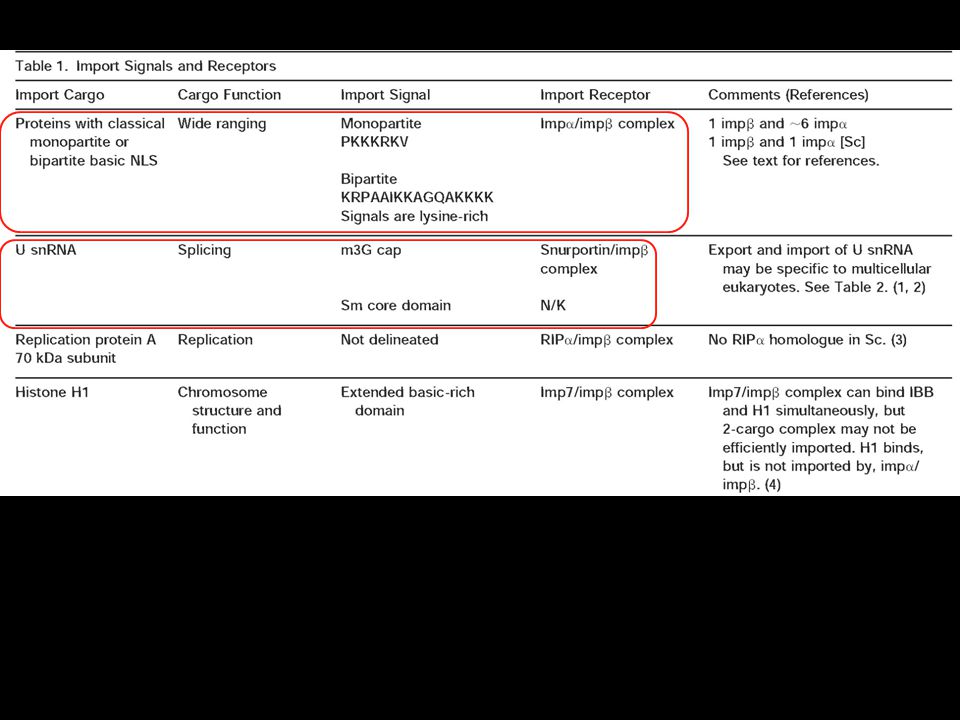
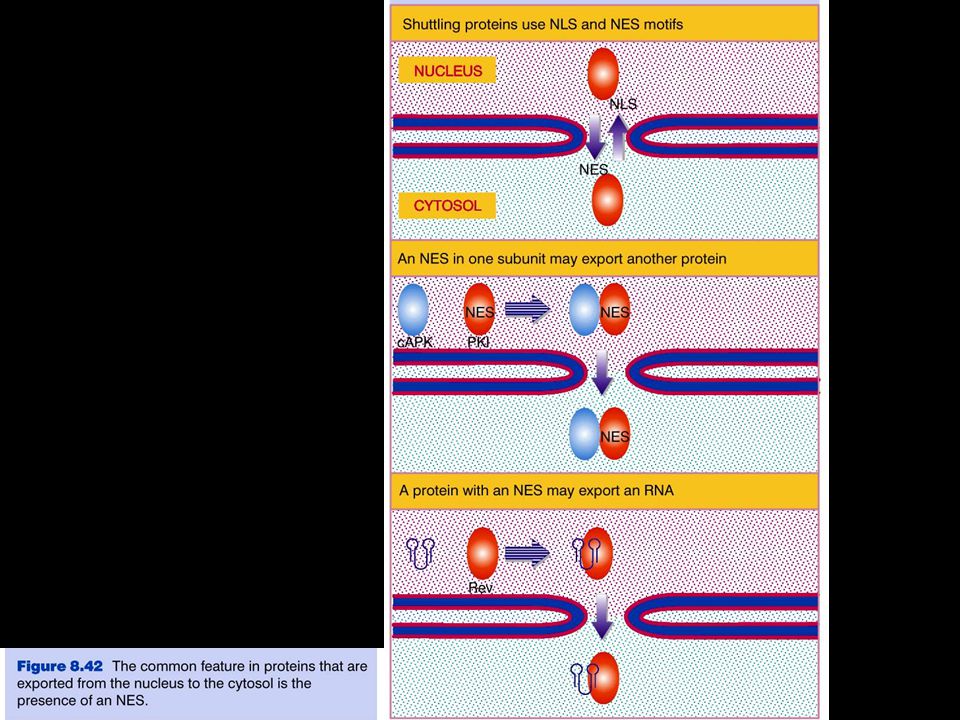
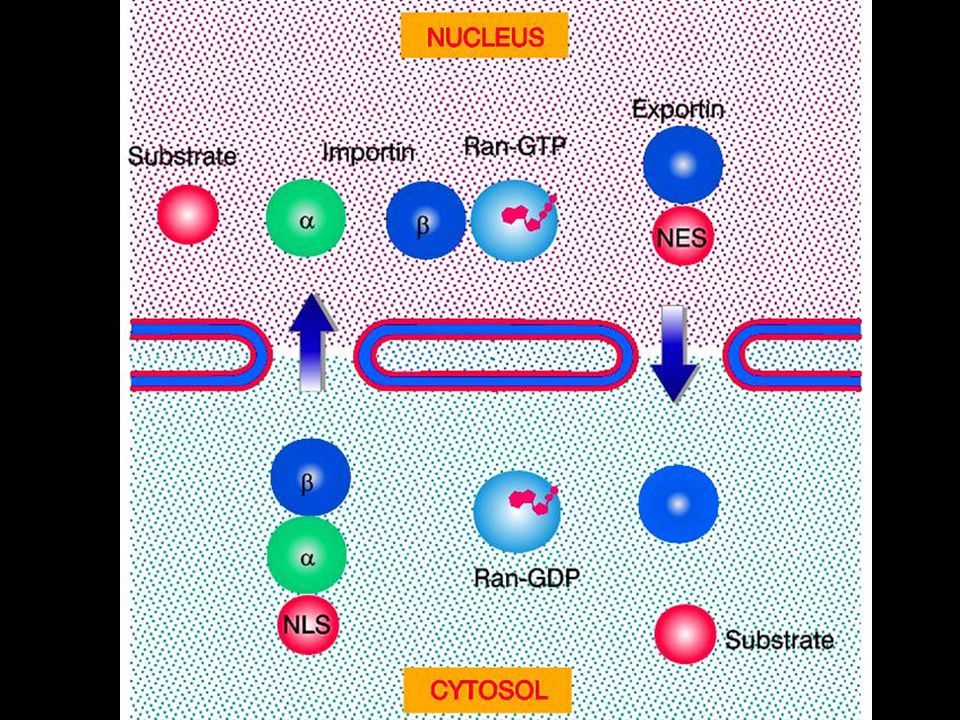
The NLS and NES consist of short sequences that are necessary and sufficient for proteins to be transported through the nuclear pores. Transport receptors have the dual properties of recognizing NLS or NES sequences and binding to the nuclear pore. The direction of transport is controlled by the state of the monomeric G protein, Ran. The nucleus contains Ran-GTP, which stabilizes export complexes, while the cytosol contains Ran-GDP, which stabilizes import complexes. The mechanism of movement does not involve a motor.

Presentazioni simili













 Piccole molecole di RNA (20-22 nt)>")









