Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
1-incontro 2-riconoscimento 3-reazione acrosomiale 4-cono di fecondazione 5- depolarizzazione della membrana 6-reazione corticale 7- fusione dei pronuclei 8 - attivazione del citoplasma

3
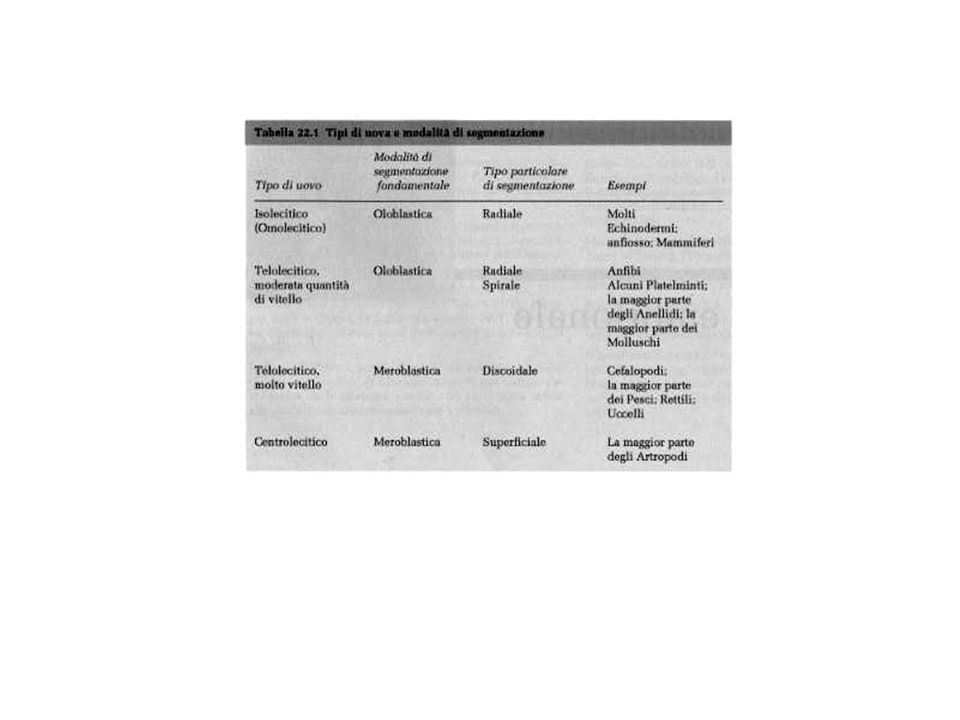
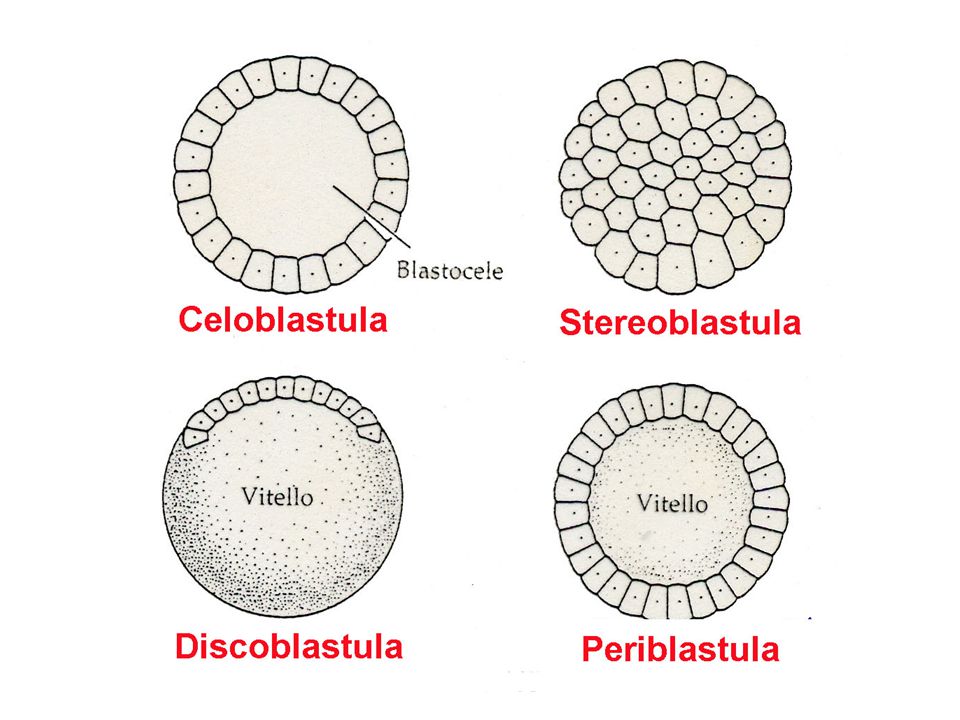
Si conoscono due tipi principali di segmentazione:
segmentazione totale (oloblastica) e segmentazione parziale (meroblastica. La segmentazione totale, o oloblastica, coinvolge tutta la cellula dello zigote; la segmentazione parziale, o meroblastica, coinvolge solo una parte della cellula zigotica.
 e segmentazione parziale (meroblastica. La segmentazione totale, o oloblastica, coinvolge tutta la cellula dello zigote; la segmentazione parziale, o meroblastica, coinvolge solo una parte della cellula zigotica.")
4
LA SEGMENTAZIONE OLOBLASTIACA PUO’ ESSERE
SPIRALE O RADIALE

5
La segmentazione parziale si suddivide in discoidali e superficiale.
La segmentazione parziale discoidale si osserva solamente nei Cefalopodi e in parte dei Cordati e si segmenta una piccola zona, a forma lenticolare, che contiene buona parte del plasma formativo.

6
SEGMENTAZIONE MEROBLASTICA SUPERFICIALE
SEGMENTAZIONE MEROBLASTICA DISCOIDALE

8
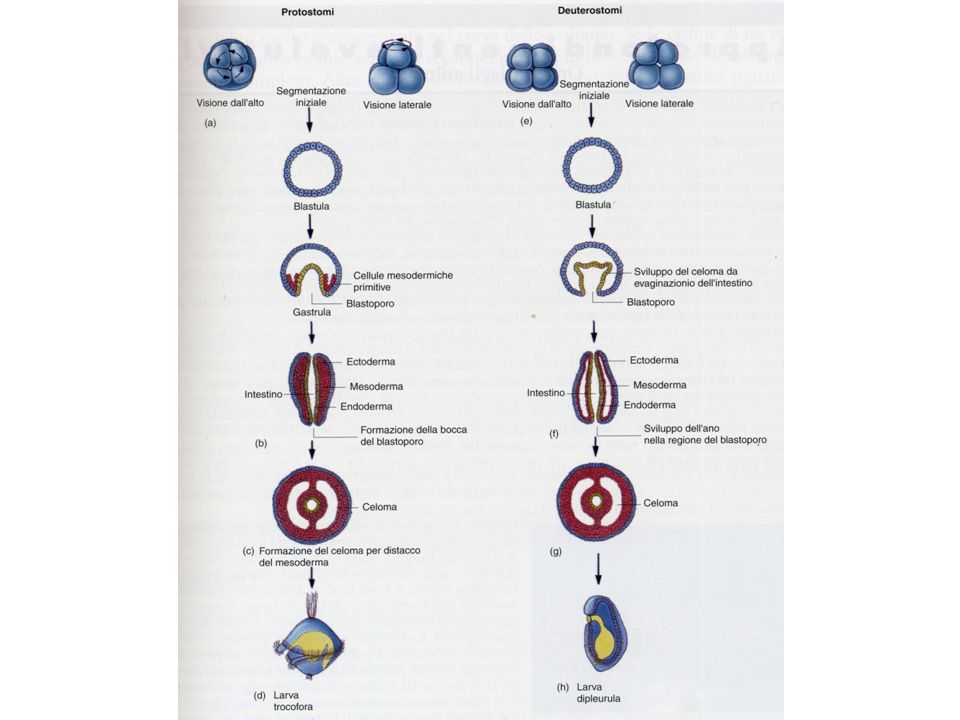
Successivamente interviene un processo di gastrulazione durante il quale le cellule della blastula si organizzano in tre foglietti embrionali (ectoderma, entoderma, mesoderma

9
GASTRULAZIONE

10
Il celoma

12
GASTRULAZIONE ORGANOGENESI SVILUPPO DIRETTO SVILUPPO INDIRETTO

16
PARTENOGENESI E GINOGENESI
La partenogenesi (dal greco παρθενος, «vergine» e γενεσις, «nascita», ovvero riproduzione verginale) è la modalità di riproduzione sessuata non anfigonica più frequente in natura e può essere considerata come una riproduzione sessuale “asessuata” perché, pur implicando la formazione di gameti, non richiede fecondazione. La discendenza può essere: -telitoca (discendenza da sole femmine) -arrenotoca (discendenza da soli maschi) -deuterotoca (discendenza composta da entrambi i sessi).
 è la modalità di riproduzione sessuata non anfigonica più frequente in natura e può essere considerata come una riproduzione sessuale asessuata perché, pur implicando la formazione di gameti, non richiede fecondazione. La discendenza può essere: -telitoca (discendenza da sole femmine) -arrenotoca (discendenza da soli maschi) -deuterotoca (discendenza composta da entrambi i sessi).")
17
- Fasmidi: partenogenesi diploide telitoca es. Bacillus rossius.
La partenogenesi si è evoluta in numerosi gruppi animali, indipendentemente da rigide compartimentazioni sistematiche, a più livelli di evoluzione e affinamento: partenogenesi rudimentale : processo di auto-attivazione dell’uovo, che porta alla degenerazione dell’embrione anormale partenogenesi accidentale: oltre all’auto attivazione dell’uovo, compare sempre accidentalmente una regolazione genetica per restituzione anafasica. Uno degli esempi più noti fra i Vertebrati è rappresentato dalla partenogenesi accidentale arrenotoca del tacchino partenogenesi facoltativa: In talune specie, i gameti femminili possono svilupparsi con ottima resa sia partenogeneticamente che anfigonicamente. - Fasmidi: partenogenesi diploide telitoca es. Bacillus rossius. - Imenotteri: partenogenesi aploide arrenotoca

18
Omotteri: Phylloxera vitifolii
Partenogenesi obbligatoria: si riproducono esclusivamente clonalmente. Diploide telitoca: popolazione di sole femmine. Partenogenesi ciclica: Omotteri: Phylloxera vitifolii Phylum Rotifera gli individui sessuati si accoppiano in autunno avanzato e depongono sul legno vecchio delle viti le "uova d'inverno"; da queste uova, in primavera si sviluppano forme giovanili, le fondatrici gallecole, che si spostano sulla pagina inferiore di una foglia, formando gradualmente una galla. Le gallecole, femmine partenogenetiche, si sviluppano in 15 giorni e danno luogo ad altre forme giovanili (neogallecole gallecole), che invadono tutte le parti della pianta originando nuove galle. Nell'arco dell'estate si susseguono sei o sette generazioni analoghe di queste femmine senza ali. All'avvicinarsi della stagione fredda, le forme giovanili (neogallecole radicicole) si spostano sulle radici, producendovi delle galle. Verso la fine dell'estate e in autunno alcune di queste gallecole radicicole danno origine a femmine alate che sfuggono dal terreno e volano sulle piante vicine, deponendo uova da cui si svilupperanno gli individui sessuati. A questo punto il ciclo si ripete.
, che invadono tutte le parti della pianta originando nuove galle. Nell arco dell estate si susseguono sei o sette generazioni analoghe di queste femmine senza ali. All avvicinarsi della stagione fredda, le forme giovanili (neogallecole radicicole) si spostano sulle radici, producendovi delle galle. Verso la fine dell estate e in autunno alcune di queste gallecole radicicole danno origine a femmine alate che sfuggono dal terreno e volano sulle piante vicine, deponendo uova da cui si svilupperanno gli individui sessuati. A questo punto il ciclo si ripete.")
19
La contemporanea presenza di caratteri sessuali secondari, maschili e femminili, in uno stesso individuo viene definita ginandromorfimo. ERMAFRODITISMO Specie monoiche Sufficiente: tenie (verme solitario) Insufficiente: Ermafroditi simultanei : lombrichi, turbellari Ermafroditi sequenziali, proterandri o proterogini
 Insufficiente: Ermafroditi simultanei : lombrichi, turbellari. Ermafroditi sequenziali, proterandri o proterogini.")
20
ERMAFRODITISMO INSUFFICIENTE:
Impedimenti morfologici, temporali, comportamentali ecc Es. temporale PROTERANDRIA Alcuni pesci ( es le cernie), nel corso del ciclo vitale, vanno incontro a inversione sessuale da maschio a femmina: producono spermatozoi da giovani e uova dall’età di circa 5 anni. .
, nel corso del ciclo vitale, vanno incontro a inversione sessuale da maschio a femmina: producono spermatozoi da giovani e uova dall’età di circa 5 anni. .")
21
Il Gasteropode Crepidula fornicata Gli individui di questa specie vivono a stretto contatto disponendosi l’uno sopra l’altro. L’individuo più in basso è quello di maggiori dimensioni; su di esso si dispongono individui di dimensioni decrescenti, fino al più piccolo all’apice della piramide. L’individuo in basso è una femmina seguono poi individui in procinto di cambiare il sesso e poi i piccoli maschi più in alto.

22
PROTEROGINIA Si sviluppa prima la gonade femminile. Es. alcuni Labridi delle barriere coralline, cernie (Epinephelus sp.), menola (Maena maena), donzella (Coris julis), pesci portaspada (Xiphophorus spp.). In molti Labridi delle barriere coralline un maschio può vivere insieme con varie femmine, costituendo una specie di "harem"; alla sua morte la femmina di maggiori dimensioni inverte la sua sessualità trasformandosi in maschio ed assumendo a sua volta il controllo dell’harem. In altri casi, se il numero delle femmine diviene molto elevato, superiore a quello che può gestire un singolo maschio, una femmina può cambiare sesso e portare via parte dell’harem.
, menola (Maena maena), donzella (Coris julis), pesci portaspada (Xiphophorus spp.). In molti Labridi delle barriere coralline un maschio può vivere insieme con varie femmine, costituendo una specie di harem ; alla sua morte la femmina di maggiori dimensioni inverte la sua sessualità trasformandosi in maschio ed assumendo a sua volta il controllo dell’harem. In altri casi, se il numero delle femmine diviene molto elevato, superiore a quello che può gestire un singolo maschio, una femmina può cambiare sesso e portare via parte dell’harem.")
23
l’ermafroditismo sequenziale si basa sul vantaggio selettivo della taglia dei sessi (ipotesi 1) e sull’impedimento dell’inincrocio (ipotesi 2). Ipotesi 1: nell’ermafroditismo proterandrico le femmine più grandi producono più uova, d’altra parte nel proteroginico il maschio più grande difende meglio il territorio e l’harem. Es. la cernia difende un territorio da altri maschi, dove feconda varie femmine. Ipotesi 2: spesso i pesci crescono in banchi insieme e nascendo di un unico sesso e invertendolo più o meno simultaneamente s’impedisce l’inincrocio.

24
DETERMINAZIONE DEL SESSO
La determinazione del sesso negli animali può avvenire al momento della fecondazione (determinazione del sesso singamica), prima (progamica) o dopo la fecondazione (metagamica).
, prima (progamica) o dopo la fecondazione (metagamica).")
25
Meccanismi di natura genetica (GSD, Genetic Sex Determination)
DETERMINAZIONE DEL SESSO SINGAMICA Meccanismi di natura genetica (GSD, Genetic Sex Determination) Presenza di un gene dominante Sistema a digametia maschile: il maschio è portatore di 2 cromosomi sessuali diversi (XY), la femmina è indicata XX (es. insetti, rettili, pesci, mammiferi, ); sistema femminile XX e maschile X0 di molti insetti. Sistema a digametia femminile: presente negli uccelli, nei lepidotteri e in alcuni rettili in cui sono presenti cromosomi sessuali: maschi ZZ e femmine WZ.
 Presenza di un gene dominante. Sistema a digametia maschile: il maschio è portatore di 2 cromosomi sessuali diversi (XY), la femmina è indicata XX (es. insetti, rettili, pesci, mammiferi, ); sistema femminile XX e maschile X0 di molti insetti. Sistema a digametia femminile: presente negli uccelli, nei lepidotteri e in alcuni rettili in cui sono presenti cromosomi sessuali: maschi ZZ e femmine WZ.")
26
Meccanismi di natura genetica (GSD, Genetic Sex Determination)
2) meccanismo dosaggio-dipendente (DSD, Dosage-dependent Sex Determination) Sia Drosophila che Caenorabditis elegans presentano un meccanismo di determinazione del sesso del tipo dosaggio-dipendente: lo sviluppo dei caratteri sessuali è controllato dal rapporto tra i cromosomi sessuali X e gli autosomi. Infatti, nonostante Drosophila presenti un cromosoma Y nel maschio, esso è irrilevante ai fini della determinazione del sesso, poiché individui XO sono maschi e individui XXY sono femmine.
 meccanismo dosaggio-dipendente. (DSD, Dosage-dependent Sex Determination) Sia Drosophila che Caenorabditis elegans presentano un meccanismo di determinazione del sesso del tipo dosaggio-dipendente: lo sviluppo dei caratteri sessuali è controllato dal rapporto tra i cromosomi sessuali X e gli autosomi. Infatti, nonostante Drosophila presenti un cromosoma Y nel maschio, esso è irrilevante ai fini della determinazione del sesso, poiché individui XO sono maschi e individui XXY sono femmine.")
27
Meccanismi di natura ambientale
(ESD, Environmental Sex Determination) La determinazione ambientale del sesso è regolata da vari fattori ambientali es.temperatura, il fotoperiodo e la densità di popolazione prima ella fecondazione (progamica): nel polichete Dinophilus gyrociliatus sono prodotte uova più grandi che daranno femmine e più piccole, maschi. dopo la fecondazione (metagamica) Es. Bonellia viridis, la cui sessualità viene determinata dalla CO2 ambientale: la concentrazione bassa di CO2 nell'acqua di mare , infatti, trasforma le larve di questo animale in femmine. Se. però, una larva non ancora determinata viene a contatto con una femmina adulta, l'ulteriore CO2 prodotta da tali organismi provoca lo sviluppo della larva in direzione maschile e precisamente in un piccolo maschio parassita, permanentemente attaccato all'organo escretore della femmina.
 La determinazione ambientale del sesso è regolata da vari fattori ambientali es.temperatura, il fotoperiodo e la densità di popolazione. prima ella fecondazione (progamica): nel polichete Dinophilus gyrociliatus sono prodotte uova più grandi che daranno femmine e più piccole, maschi. dopo la fecondazione (metagamica) Es. Bonellia viridis, la cui sessualità viene determinata dalla CO2 ambientale: la concentrazione bassa di CO2 nell acqua di mare , infatti, trasforma le larve di questo animale in femmine. Se. però, una larva non ancora determinata viene a contatto con una femmina adulta, l ulteriore CO2 prodotta da tali organismi provoca lo sviluppo della larva in direzione maschile e precisamente in un piccolo maschio parassita, permanentemente attaccato all organo escretore della femmina.")
28
DETERMINAZIONE AMBIENTALE DEL SESSO
In alcuni pesci il fenotipo sessuale può variare nell'adulto ed è controllato da un insieme di fattori dinamici che includono segnali ormonali e visivi; in molti rettili (alligatori e tartarughe) il fenotipo sessuale dipende dalla temperatura di incubazione dell'embrione.
 il fenotipo sessuale dipende dalla temperatura di incubazione dell embrione.")
29
CARATTERI SESSUALI Il tipo di gonadi presenti in un individuo ne definisce i caratteri sessuali primari; altre caratteristiche invece definiscono i caratteri sessuali secondari e comprendono la presenza di barba, creste, speroni, particolari colorazioni, mammelle, marsupi (nei marsupiali), organi che facilitano la copulazione o la deposizione delle uova, una diversa distribuzione corporea del pelo, un particolare timbro della voce e così via.
, organi che facilitano la copulazione o la deposizione delle uova, una diversa distribuzione corporea del pelo, un particolare timbro della voce e così via.")
30
apparati riproduttori
maschili e femminili negli insetti

31
CARATTERI SESSUALI SECONDARI
I caratteri sessuali secondari possono essere di due tipi: somatosessuali ed eusessuali; i primi, controllati geneticamente, sono caratteristici degli artropodi e di pochi vertebrati, gli altri, sotto il controllo ormonale, sono tipici dei vertebrati. Particolari ormoni influenzano la differenziazione dei caratteri sessuali secondari durante l’adolescenza nell'uomo, quali lo sviluppo del seno e dei fianchi nella femmina, la barba ed allargamento del pomo d’Adamo nel maschio.

32
DIMORFISMO SESSUALE E SELEZIONE SESSUALE: Il “costo del sesso”

Presentazioni simili



 Dominanza incompleta>")


















