Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
CORSO DI IMMUNOLOGIA per il corso di Laurea in biotecnologie a. a
CORSO DI IMMUNOLOGIA per il corso di Laurea in biotecnologie a.a II semestre DOCENTE: Dott.ssa Vladia Monsurrò Dipartimento di patologia Sezione di immunologia Universita’ degli Studi di Verona ORARIO DELLE LEZIONI: lunedì: 4:00-5:30 giovedì 4:00-5:30 dal 20 marzo al 25 maggio 2006 aula F Facolta’ di Scienze

2
DOVE TROVARE LE LEZIONI on line:
In “materiale didattico” Per qunato riguarda il programma e l’esame e’ come discusso in classe

3
PROGRAMMA LEZIONI Mar 21 Marzo lezione 1 Generalità sul corso
Il sistema immunitario (Introduzione) Giov 23 Marzo lezione 2 Cellule e tessuti del sistema immunitario Lun 27 Marzo lezione 3 Riconoscimento dell’antigene: Molecole del Sistema Immunitario: Antigeni e anticorpi Giov 30 Marzo lezione 4 MHC (Major histocompatibility complex) Lun 3 Aprile lezione 5 MHC-Antigene: Sua captazione, processamento e presentazione. Giov 6 Aprile lezione 6 TCR Antigeni tumorali e immunoterapia (in preparazione del talk) Lun 10 Aprile lezione 7 Maturazione, attivazione e regolazione dei linfociti: Maturazione dei linfociti ed espressione dei recettori (genetica Ig e TCR) Giov 13 Aprile lezione 8 Immunita’ e tumori Parmiani Talk (ore 17:00)-vedi volantino
 Giov 23 Marzo lezione 2. Cellule e tessuti del sistema immunitario. Lun 27 Marzo lezione 3. Riconoscimento dell’antigene: Molecole del Sistema Immunitario: Antigeni e anticorpi. Giov 30 Marzo lezione 4. MHC (Major histocompatibility complex) Lun 3 Aprile lezione 5. MHC-Antigene: Sua captazione, processamento e presentazione. Giov 6 Aprile lezione 6. TCR. Antigeni tumorali e immunoterapia (in preparazione del talk) Lun 10 Aprile lezione 7. Maturazione, attivazione e regolazione dei linfociti: Maturazione dei linfociti ed espressione dei recettori (genetica Ig e TCR) Giov 13 Aprile lezione 8. Immunita’ e tumori. Parmiani Talk (ore 17:00)-vedi volantino.")
4
Giov 20 Aprile lezione 9 Attivazione linfocitaria, trasduzione del segnale. Lun 24 Aprile lezione 10 Attivazione dei linfociti B e produzione di Ab Giov 27 Aprile lezione 11 Tolleranza immunologica Giov 4 Maggio lezione 12 Meccanismi effettori Citochine, Immunita’ innata Lun 8 Maggio lezione 13 Meccanismi effettori dell’immunita’ cellulo-mediata, reazioni DTH Giov 11 Maggio lezione 14 Meccansimi effettori dell’immunita’ umorale complemento Lun 15 Maggio lezione 15 Rapporti tra fisiologia e patologia Ipersensibilità concetti generali Ipersensibilità di tipo I Giov 18 Maggio lezione 16 Test prove esami-recupero Merc 7 giugno e 28 giugno ESAMI

5
I RECETTORI PER GLI ANTIGENI:
MHC TCR

6
due catene pesanti (isotipo oppure , , , )
STRUTTURA FONDAMENTALE DI UNA MOLECOLA DI IMMUNOGLOBULINA: catene PESANTI catene LEGGERE due catene pesanti (isotipo oppure , , , ) due catene leggere (isotipo k o ) ponti disolfuro che legano tra loro le due catene pesanti e ciascuna catena leggera ad una catena pesante
 due catene leggere. (isotipo k o ) ponti disolfuro che legano tra loro le due catene pesanti e ciascuna catena leggera ad una catena pesante.")
7
REGIONI COSTANTI E REGIONI VARIABILI
VH regioni COSTANTI CH3 CH2 CL CH1 VL

8
STRUTTURA FONDAMENTALE DI UNA MOLECOLA DI IMMUNOGLOBULINA
VH CH1 VL catena leggera: -1 dominio variabile VL -1 dominio costante CL CL CH2 catena pesante: 1 dominio variabile VH 3 o 4 domini costanti CH, a seconda dell'isotipo CH3

9
Struttura molecolare degli anticorpi:

10
PRODUZIONE DI ANTICORPI MONOCLONALI

11
I RECETTORI PER GLI ANTIGENI:
MHC TCR

12
RICONOSCIMENTO DELL’ANTIGENE :
- linfociti B antigene in forma nativa -linfociti T antigene processato da cellule accessorie (cellule presentanti l’antigene )
")
13
I linfociti T contro microbi intracellulari e l’attivazione di altre cellule (macrofagi e linfociti B)
")
14
Le APC (=Antigen Presenting Cell) l’antigene processato in peptidi
“PROFESSIONISTE” presentano l’antigene processato ai linfociti T associato alle molecole MHC recettore MHC legante l’antigene processato in peptidi Macrofago Linfocita B Cellula dendritica

15
Le molecole Le molecole MHC-II presentano MHC-I presentano
PRESENTAZIONE DELL’ANTIGENE CAPTATO DALL’ESTERNO E PROCESSATO DA PARTE DELLE CELLULE DENDRITICHE Cellula dendritica MHC-II Antigene processato MHC-I Antigene processato Linfocita T helper e suoi recettori e co-recettori per l’antigene Le molecole MHC-I presentano l’ Ag ai T citotossici Le molecole MHC-II presentano l’ Ag ai T helper Linfocita T citotossico, suoi recettori per l'antigene e co-recettori CD8

16
MOLECOLE MHC struttura e genetica

17
Cosa significa MHC? MHC (complesso maggiore di istocompatibilità)
è il locus dove mappano gli HLA È chiamato così perché storicamente e’ stato associato ai rigetti di trapianto (altamente polimorfico e immunogeno).
.")
18
Un allele è una variante di un gene polimorfo
Un individuo è: Omozigote se ha 2 alleli uguali Eterozigote se ha 2 alleli differenti

19
Topi singenici hanno tutti i geni identici
Due ceppi allogenici sono due gruppi di topi singenici differenti fra loro

20
Benacerraf e McDevitt scoprirono tra gli
IL COMPLESSO GENETICO MHC VENNE INIZIALMENTE IDENTIFICATO COME REGIONE GENETICA CHE CONTROLLAVA IL RIGETTO DEI TRAPIANTI TRA TOPI DI CEPPI INBRED DIVERSI (Snell-1940) Benacerraf e McDevitt scoprirono tra gli anni 1960/1970 che il complesso MHC controllava anche la RISPOSTA ANTICORPALE contro determinati antigeni proteici, ed era diversa a seconda degli alleli MHC di ciascun ceppo Questo perchè alcuni peptidi li legavano e generavano rispost aT helper, altri no Esistono dei geni di istocompatibilità
 Benacerraf e McDevitt scoprirono tra gli. anni 1960/1970 che il complesso MHC. controllava anche la RISPOSTA. ANTICORPALE contro determinati. antigeni proteici, ed era diversa a seconda. degli alleli MHC di ciascun ceppo. Questo perchè alcuni peptidi li legavano. e generavano rispost aT helper, altri no. Esistono dei geni di istocompatibilità.")
21
Regione MHC nell’uomo e nel topo
centromero telomero

22
Regione MHC nell’uomo centromero telomero
Map of the human MHC. This map is simplified to exclude many genes that are of unknown function. HLA-E, HLA-F, HLA-G, HLA-J, and HLA-X are class I-like molecules; HLA-H does not appear to be involved in the immune system. C4, C2, B, complement proteins; DM, TAP, proteasomes, proteins involved in antigen processing; HLA, human leukocyte antigen; HSP, heat shock protein; LT, TNF, cytokines

23
Collocazione dei geni MHC e dei geni MHC-relati sui cromosomi umani

24
Gli MHC nell’uomo sono chiamati HLA (antigeni leucocitari umani) perche’ identificati per vie sierologiche mediante Ab che riconoscevano i leucociti durante le trasfusioni
 perche’ identificati per vie sierologiche mediante Ab che riconoscevano i leucociti durante le trasfusioni")
25
NOMENCLATURA HLA © 2003 by LIPPINCOTT WILLIAMS & WILKINS
Nomenclature of human leukocyte antigen (HLA) loci and alleles Nomenclature Definition HLA The HLA region and prefix for an HLA gene HLA-DRB1 A particular HLA locus (i.e., DRB1) HLA-DRB1*13 A group of alleles that encode the DR13 antigen defined serologically by microlymphocytotoxicity or by mixed lymphocyte reactivity HLA-DRB1*1301 A specific HLA allele HLA-DRB1*1301N A null allele (i.e., nonexpressed) HLA-DRB1*13012 An allele that differs by a synonymous mutation (i.e., identical amino acid encoded by a different codon) HLA-DRB1* An allele that contains a mutation outside the coding region HLA-DRB1* N A null allele that contains a mutation outside the coding region © 2003 by LIPPINCOTT WILLIAMS & WILKINS
 loci and alleles. Nomenclature. Definition. HLA. The HLA region and prefix for an HLA gene. HLA-DRB1. A particular HLA locus (i.e., DRB1) HLA-DRB1*13. A group of alleles that encode the DR13 antigen defined. serologically by microlymphocytotoxicity or by mixed lymphocyte reactivity. HLA-DRB1*1301. A specific HLA allele. HLA-DRB1*1301N. A null allele (i.e., nonexpressed) HLA-DRB1* An allele that differs by a synonymous mutation (i.e., identical amino acid. encoded by a different codon) HLA-DRB1* An allele that contains a mutation outside the coding region. HLA-DRB1* N. A null allele that contains a mutation outside the coding region. © 2003 by LIPPINCOTT WILLIAMS & WILKINS.")
26
© 2003 by LIPPINCOTT WILLIAMS & WILKINS Fundamental Immunology
. Renal graft survival improves with fewer human leukocyte antigen (HLA) mismatches. Cumulative data for graft survival are plotted as a function of time. Curves 1, 2, and 3 represent the groups with no class II HLA mismatches and four or fewer class I HLA mismatches, whereas curve 4 shows the graft survival of those with fewer than two class II mismatches and no class I mismatch. The worst survivals are clearly those with both HLA class I and class II mismatches © 2003 by LIPPINCOTT WILLIAMS & WILKINS Fundamental Immunology
 mismatches. Cumulative data for graft survival are plotted as a function of time. Curves 1, 2, and 3 represent the groups with no class II HLA mismatches and four or fewer class I HLA mismatches, whereas curve 4 shows the graft survival of those with fewer than two class II mismatches and no class I mismatch. The worst survivals are clearly those with both HLA class I and class II mismatches. © 2003 by LIPPINCOTT WILLIAMS & WILKINS Fundamental Immunology.")
27
RICONOSCIMENTO ANTIGENICO NEI TRAPIANTI

29
Gli alleli dell’ MHC sono altamente polimorfici
Codominanti Cl I e Cl II sono distinte ma omologhe

30
POLIMORFISMO DELLE MOLECOLE
MHC Un allele ad un locus genetico che abbia una frequenza 0.01 è denominato “rara variante allelica”. Un allele con una frequenza 0.01 (ma 1) è definito come “polimorfico”. Gli alleli ai loci MHC sono polimorfici poichè presentano sempre frequenze elevate nella popolazione. Il polimorfismo è concentrato negli esoni che codificano per i domini che legano il peptide e il TCR.
 è definito. come polimorfico . Gli alleli ai loci MHC sono polimorfici poichè presentano. sempre frequenze elevate nella popolazione. Il polimorfismo è concentrato negli esoni che codificano. per i domini che legano il peptide e il TCR.")
31
I loci MHC presentano un numero molto elevato di alleli:
NUMERO DI ALLELI HLA AD ALCUNI LOCI I loci MHC presentano un numero molto elevato di alleli:

32
Frequenze alleliche in popolazioni diverse

33
I geni MHC sono espressi in maniera codominante

34
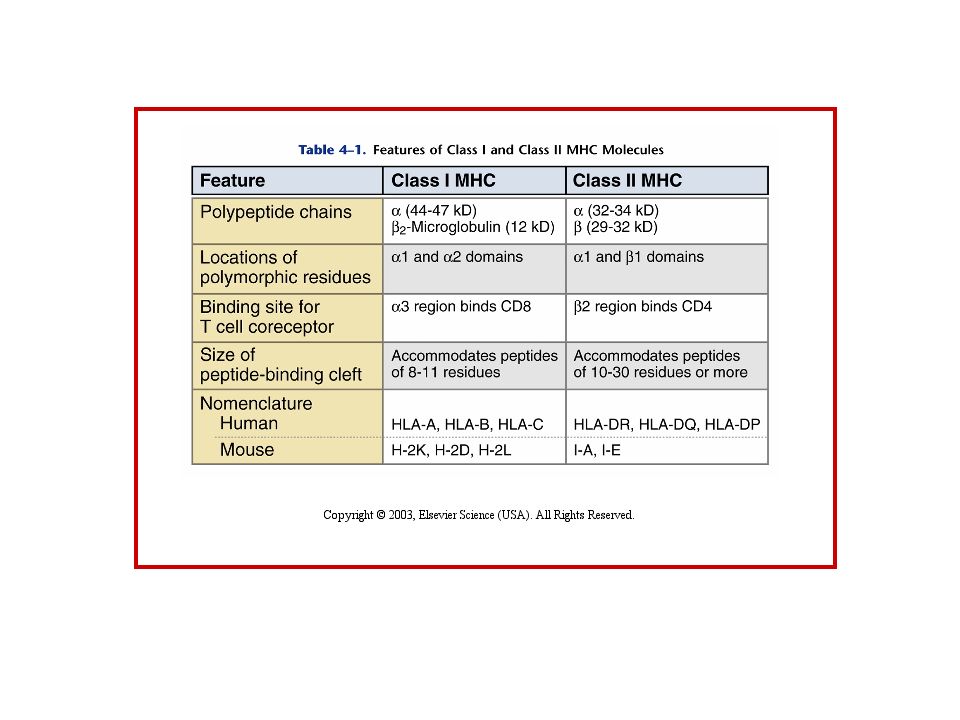
Struttura dell’MHC

35
Similitudini Cl I e Cl II
Hanno una tasca di legame per il peptide, due domini Ig, dominio transmembrana e dominio citoplasmatico Il polimorfismo e’ nella tasca Le zone non polimorfe legano i recettori CD4 o CD8

36
STRUTTURA SCHEMATICA DELLE MOLECOLE
MHC CLASSICHE Secondo la regola: 1 gene-1 catena polipeptidica -1 gene per la catena - 1 gene per la catena

37
Similitudini ClI e Cl II
Hanno una tasca di legame pe ril peptide, due domini Ig, dominio transmembrana e dominio citoplasmatico Il polimorfismo e’ nella tasca Le zone non polimorfe legano i recettori CD4 o CD8

38
STRUTTURA DEL TCR

39
T cell recognition of a peptide-MHC complex
T cell recognition of a peptide-MHC complex. This schematic illustration shows an MHC molecule binding and displaying a peptide and a T cell receptor recognizing two polymorphic residues of the MHC molecule and one residue of the peptide. Details of the interactions among peptides, MHC molecules, and T cell receptors are described in Chapters 4, 5, and 6.

40
Modello delle interazioni molecolari tra MHC-II/peptide,
TCR e CD4, e MHC-I/peptide, TCR e CD4 APC o cell. bersaglio MHC-II CD8 si lega al dominio 3 di MHC-I MHC-I CD4 CD4 si lega al dominio 2 di MHC-II CD8 TCR TCR Cellula T

41
MHC Cl I

42
2 tipi di Molecole MHC-I classiche, MHC-Ia: non classiche, MHC-Ib:
HLA-A,-B,-C (uomo) H-2K,-D,-L (topo) sono polimorfiche e legano peptidi antigenici che presentano ai TCR associati a CD8 Essendo codominanti ogni cellula ne esprime 6 diverse non classiche, MHC-Ib: CD1 H2-M3 MICA nFcR Rae1 Presentano polimorfismo molto limitato e spesso il solco di legame con il peptide è molto ridotto
 H-2K,-D,-L (topo) sono polimorfiche e legano peptidi antigenici che presentano ai TCR associati a CD8. Essendo codominanti ogni cellula ne esprime 6 diverse. non classiche, MHC-Ib: CD1. H2-M3. MICA. nFcR. Rae1. Presentano polimorfismo molto limitato e spesso il solco di legame con il peptide è molto ridotto.")
43
STRUTTURA SCHEMATICA DELLE MOLECOLE MHC
Catena : 44-47kD Peptide 8-10 aa Catena : 2-microglobulina, 12kD 2 catene polipeptidiche e associate non covalentemente con strutture secondarie costituite da foglietti e -eliche.

44
MHC Cl II

45
STRUTTURA SCHEMATICA DELLE MOLECOLE MHC
Catena : 32-34kD Peptide fino a 30 aa Catena : 29-32kD 2 catene polipeptidiche e associate non covalentemente con strutture secondarie costituite da foglietti e -eliche.

46
Circa 20 possibili combinazioni perche’ DQ puo’ legarsi anche a DP e DR, sia materno che paterno

47
Le molecole MHC-I ed MHC-II completamente
assemblate sono eterotrimeri, composti da: una catena ; una catena ; il peptide antigenico nella tasca. L’espressione stabile sulla membrana cellulare richiede la presenza di tutti e tre i componenti.

49
CDR= ragione che determina la complementarieta’
Ig= immunoglobulina Kd= costante di dissociazione MHC= complesso maggiore di istocompatibilita’ VH= dominio variabile della catena pesante VL= dominio variabile della catena leggera

50
Legame MHC-antigene

51
Similitudini ClI e Cl II
Hanno una tasca di legame pe ril peptide, due domini Ig, dominio transmembrana e dominio citoplasmatico Il polimorfismo e’ nella tasca Le zone non polimorfe legano i recettori CD4 o CD8

52
La variabilità delle sequenze aminoacidiche tra i diversi
alleli ad un locus è diversa a seconda del dominio molecolare: è concentrata nei domini che legano il peptide.

53
Polymorphic residues of a class I MHC molecule
Polymorphic residues of a class I MHC molecule. The polymorphic residues of class I and class II MHC molecules (shown as red circles) are located in the peptide-binding clefts and the a-helices around the clefts. In the class II molecule shown (HLA-DR), essentially all the polymorphism is in the b chain. However, other class II molecules in humans and mice show varying degrees of polymorphism in the a chain and usually much more in the b chain.
 are located in the peptide-binding clefts and the a-helices around the clefts. In the class II molecule shown (HLA-DR), essentially all the polymorphism is in the b chain. However, other class II molecules in humans and mice show varying degrees of polymorphism in the a chain and usually much more in the b chain.")
54
Peptide binding to MHC molecules. A
Peptide binding to MHC molecules. A. These top views of the crystal structures of MHC molecules show how peptides (in yellow) lie on the floors of the peptide-binding clefts and are available for recognition by T cells. The class I molecule shown is HLA-A2, and the class II molecule is HLA-DR1. Note that the cleft of the class I molecule is closed, whereas that of the class II molecule is open. As a result, class II molecules accommodate longer peptides than do class I molecules. (Courtesy of Dr. P. Bjorkman, California Institute of Technology, Pasadena.) B. The side view of a cut-out of a peptide bound to a class II MHC molecule shows how anchor residues of the peptide hold it in the pockets in the cleft of the MHC molecule. (From Scott CA, PA Peterson, L Teyton, and IA Wilson. Crystal structures of two I-Ad-peptide complexes reveal that high affinity can be achieved without large anchor residues. Immunity 8: , Copyright 1998, with permission from Elsevier Science.)
 lie on the floors of the peptide-binding clefts and are available for recognition by T cells. The class I molecule shown is HLA-A2, and the class II molecule is HLA-DR1. Note that the cleft of the class I molecule is closed, whereas that of the class II molecule is open. As a result, class II molecules accommodate longer peptides than do class I molecules. (Courtesy of Dr. P. Bjorkman, California Institute of Technology, Pasadena.) B. The side view of a cut-out of a peptide bound to a class II MHC molecule shows how anchor residues of the peptide hold it in the pockets in the cleft of the MHC molecule. (From Scott CA, PA Peterson, L Teyton, and IA Wilson. Crystal structures of two I-Ad-peptide complexes reveal that high affinity can be achieved without large anchor residues. Immunity 8: , Copyright 1998, with permission from Elsevier Science.)")
55
MHC-I: peptide di 8-10 AA MHC-II: peptide anche di più di 30 AA

56
Rappresentazione della localizzazione di un peptide
legato nella tasca formata dai domini 1 e 2 di una molecola MHC-I. Peptide (in viola) Molecole di acqua (sfere azzurre)
 Molecole di acqua. (sfere azzurre)")
57
MHC-binding residues and T cell receptor contact residues of a model peptide antigen. The immunodominant epitope of the protein hen egg lysozyme (HEL) in H-2k mice is a peptide composed of residues The ribbon diagrams modeled after crystal structures show the surface of the peptide-binding cleft of the I-Ak class II molecule and the bound HEL peptide with amino acid residues (P1-P9) indicated as spheres (A) and with side chains (B). Mutational analysis of the peptide has shown that the residues involved in binding to MHC molecules are P1 (Asp52), P4 (Ile55), P6 (Gln57), P7 (Ile 58), and P9 (Ser60); these are the residues that project down and fit into the peptide-binding cleft. The residues involved in recognition by T cells are P2 (Tyr53), P5 (Leu56), P8 (Asn59), P10 (Arg61), and P11 (Trp62); these residues project upward and are available to T cells. (From Fremont DH, D Monnale, CA Nelson, WA Hendrickson, and ER Unanue. Crystal structure of I-A in complex with a dominant epitope of lysozyme. Immunity 8: , Copyright 1998, with permission from Elsevier Science.)
 in H-2k mice is a peptide composed of residues The ribbon diagrams modeled after crystal structures show the surface of the peptide-binding cleft of the I-Ak class II molecule and the bound HEL peptide with amino acid residues (P1-P9) indicated as spheres (A) and with side chains (B). Mutational analysis of the peptide has shown that the residues involved in binding to MHC molecules are P1 (Asp52), P4 (Ile55), P6 (Gln57), P7 (Ile 58), and P9 (Ser60); these are the residues that project down and fit into the peptide-binding cleft. The residues involved in recognition by T cells are P2 (Tyr53), P5 (Leu56), P8 (Asn59), P10 (Arg61), and P11 (Trp62); these residues project upward and are available to T cells. (From Fremont DH, D Monnale, CA Nelson, WA Hendrickson, and ER Unanue. Crystal structure of I-A in complex with a dominant epitope of lysozyme. Immunity 8: , Copyright 1998, with permission from Elsevier Science.).")
58
Caratteristiche delle interazioni peptide-MHC:
- le molecole MHC hanno una certa promiscuità nel legame col peptide; ogni molecola MHC-I o -II possiede una sola tasca di legame con il peptide, che può ospitare, in momenti diversi, molti peptidi differenti, ma con alcune caratteristiche strutturali in comune ; - la fine specificità del riconoscimento antigenico è dovuta prevalentemente alla selettività del TCR; il legame dei peptidi alle molecole MHC è un’interazione non covalente tra residui AA del peptide e residui AA della tasca della molecola MHC, saturabile a bassa affinità, con bassi tassi di associazione e di dissociazione; - le molecole MHC di un individuo non distinguono tra peptidi estranei (es. da Ag microbici) e peptidi self (= da Ag derivati da proteine proprie).
 e peptidi self (= da Ag derivati da proteine proprie).")
59
Antigen competition for T cells
Antigen competition for T cells. A T cell recognizes a peptide presented by one MHC molecule. An excess of a different peptide that binds to the same MHC molecule competitively inhibits presentation of the peptide that the T cell recognizes. APC, antigen-presenting cell.

61
Lista di alcuni peptidi trovati associati a
molecole MHC-I

62
Schema dell’interazione MHC-I/peptide

63
Schema dell’interazione MHC-II/peptide
Lista di alcuni peptidi trovati associati a molecole MHC-II

64
Scoperta di epitopi: iTOPIA (filmino)
")
65
CD8 CD3 TCR CTL Tumor cell Monsurro’ and Nagorsen ASHI Q 2002 epitopes
b 2- microglobulin HLA TAP HLA b 2- microglobulin TAA epitopes Monsurro’ and Nagorsen ASHI Q 2002

66
b2 microglobulin Soluble tagged HC (HLA) peptide Avidin labeled with PE Refolding and biotinylation Biotinilated monomer tHLA PE

67
CTL Tumor cell Monsurro’ and Nagorsen ASHI Q 2002 tPE CD8FITC b 2-
microglobulin HLA CD8 TCR epitopes TAP TAA CTL tPE CD8FITC tPE CD8FITC Monsurro’ and Nagorsen ASHI Q 2002

68
of vaccine specific CD8+ cells (ex vivo)
Kinetics of expansion of vaccine specific CD8+ cells (ex vivo) 0i 8i 16i 24i CD8 FITC t209-2M PE 0% 0.4% 0.3% 2.2% Monsurro’ Blood 2005
 0i. 8i. 16i. 24i. CD8 FITC. t209-2M PE. 0% 0.4% 0.3% 2.2% Monsurro’ Blood")
69
Utilizzo biotecnologico dell’MHC
Tetrameri (filmino)
")
70
Enhancement of class II MHC expression by IFN-g
Enhancement of class II MHC expression by IFN-g. IFN-g, produced by NK cells during innate immune reactions to microbes or by T cells during adaptive immune reactions, stimulates class II MHC expression on antigen-presenting cells (APCs) and thus enhances the activation of CD4+ T cells. IFN-g has a similar effect on the expression of class I MHC molecules and the activation of CD8+ T cells
 and thus enhances the activation of CD4+ T cells. IFN-g has a similar effect on the expression of class I MHC molecules and the activation of CD8+ T cells.")
71
The end

72
Response rate after peptide-based vaccinations against melanoma
Responses Patients % 20 11 13 31 42 Treatment : MART - 1 27 35 in IFA g209 2M in IFA 2M in IFA + IL 2 (720,000 IU/Kg TID) Rosenberg et al. Nature Med,1998
 Rosenberg et al. Nature Med,1998.")
73
Come è possibile che le molecole MHC
presentino un numero di peptidi diversi sufficiente a coprire la maggior parte delle proteine antigeniche ? Attraverso duplicazione genica ed elevato polimorfismo: tanti loci codificanti per molecole simili alleli polimorfici allo stesso locus codominanza

74
Ricadute del polimorfismo dell’MHC sull’ immunopatologia :
Trapianti Suscettibilità a varie patologie, spesso autoimmuni E’ importante in alcuni casi tipizzare i pazienti per il genotipo MHC

75
Corrispondenza tra gli esoni dei geni MHC
e i domini della struttura proteica MHC-I MHC-II

76
Ereditarietà dell’MHC nell’uomo

77
Lista degli alleli HLA di classe II

78
Lista degli alleli HLA di classe I

Presentazioni simili











 SPECIFICA/SELETTIVA:>")







>")

