Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il complemento

2
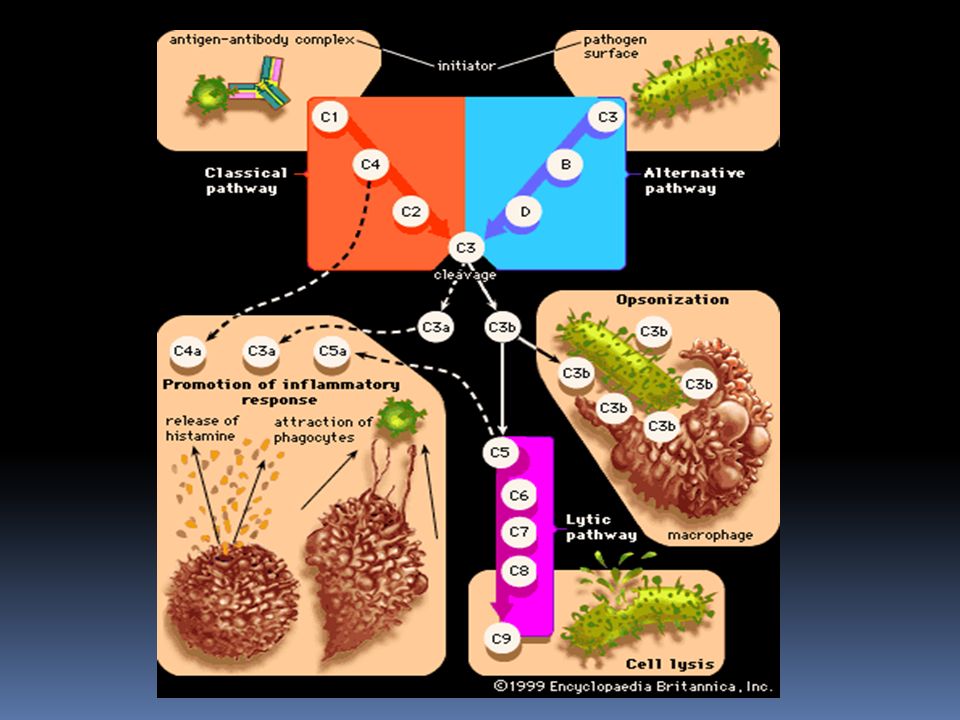
Complemento E’ formato da un complesso sistema multifattoriale, costituito da oltre 20 proteine sieriche , che ha la funzione di distruggere i batteri tramite la lisi o la fagocitosi. Soggetti affetti da deficit quantitativo o funzionale del complemento pur avendo livelli normali di Ig, sono particolarmente esposti a ripetute infezioni. Le molecole del complemento si trovano nel sangue o nei liquidi biologici, in forma inattiva finche non sono attivati da microrganismi o altri fattori. Il meccanismo di attivazione è un meccanismo a cascata, in cui il primo elemento agisce sul successivo , rendendolo attivo e quindi capace di agire sull’elemento successivo. L’attività è localizzata nella zona d’innesco con una emivita degli elementi di un millesimo di secondo. Le molecole del complemento sono indicate con una numerazione , che va da C1 al C9 , oppure con le lettere maiuscole (B,D,P). I frammenti derivati dalla molecola che ha subito l’azione enzimatica dell’elemento che lo precede nella cascata, sono indicati con le lettere minuscole (es. C5a, C5b) . Il meccanismo terminale dell’azione del complemento è la formazione di un “complesso litico”, detto complesso d’attacco alla membrana (MAC), costituito dai fattori che vanno dal C5 al C9.
. I frammenti derivati dalla molecola che ha subito l’azione enzimatica dell’elemento che lo precede nella cascata, sono indicati con le lettere minuscole (es. C5a, C5b) . Il meccanismo terminale dell’azione del complemento è la formazione di un complesso litico , detto complesso d’attacco alla membrana (MAC), costituito dai fattori che vanno dal C5 al C9.")
3
Complemento Il sistema del complemento può essere attivato , nelle sue fasi iniziali fino al fattore centrale C3 , mediante 3 vie : Via classica : viene attivata dall’interazione fra l’ immunocomplesso ( formato da antigeni ed anticorpi delle classi IgG ed IgM ), oppure fra complessi formati da Ig aggregate o denaturate, e il primo componente , il C1q. Via alternativa o properdinica : viene attivata dalla presenza di particolari prodotti d’origine batterica o virale, che interagiscono con il componente C3 . Viene innescata in assenza di anticorpi. Via lectinica : attivata dall’interazione tra una proteina della fase acuta (lectina legante il mannosio, MBL) e i residui di mannosio, presenti nelle glicoproteine o carboidrati della superficie dei microrganismi.
, oppure fra complessi formati da Ig aggregate o denaturate, e il primo componente , il C1q. Via alternativa o properdinica : viene attivata dalla presenza di particolari prodotti d’origine batterica o virale, che interagiscono con il componente C3 . Viene innescata in assenza di anticorpi. Via lectinica : attivata dall’interazione tra una proteina della fase acuta (lectina legante il mannosio, MBL) e i residui di mannosio, presenti nelle glicoproteine o carboidrati della superficie dei microrganismi.")
4
Complemento I fattori del complemento svolgono molteplici funzionalità biologiche : La capacità di lisi : delle membrane cellulari , batteriche e dell’involucro lipoproteico di alcuni virus per azione del complesso litico (C5b-C9) La capacità della risposta infiammatoria : mediante liberazione di sostanze ad azione anafilattica (vasoattiva) e chemiotattica, che favoriscono il richiamo locale di cellule fagocitanti (C3a, C5a) L’opsonizzazione : delle cellule capaci di fagocitosi (C3b,C4b) La rimozione degli immunocomplessi : avviene tramite il legame tra un frammento del complemento (C3b) e i recettori presenti su alcune cellule.
 La capacità della risposta infiammatoria : mediante liberazione di sostanze ad azione anafilattica (vasoattiva) e chemiotattica, che favoriscono il richiamo locale di cellule fagocitanti (C3a, C5a) L’opsonizzazione : delle cellule capaci di fagocitosi (C3b,C4b) La rimozione degli immunocomplessi : avviene tramite il legame tra un frammento del complemento (C3b) e i recettori presenti su alcune cellule.")
6
Via classica Il primo fattore attivato è C1 .
Il fattore C1 è costituito da un complesso tri-molecolare di 750 kDa legato in maniera non covalente , C1q, C1r, C1s , in un rapporto molare di 1/2/2 (C1qr2s2). C1q è una proteina complessa, ricca in idrossiprolina, idrossilisina e glicina che per le sue caratteristiche elettroforetiche presenta delle analogie con le Ig, mentre per la sua struttura amminoacidica ricorda le proteine del collagene. La molecola C1q è costituita da 6 unità ognuna costituita da tre distinte subunità A,B,C E’ possibile distinguere una parte centrale compatta , che lega C1r e C1 s , ed una struttura più esterna che termina con una forma a calice. La struttura a calice svolge la funzione di riconoscere e legare particolari siti presenti nelle Ig, i quali si evidenziano durante la formazione dell’immunocomplesso. La sub-unità C1q si deve legare ai domini CH2 di almeno due anticorpi IgG adiacenti . La sub-unità C1q si può legare ad un solo anticorpo IgM perche dotato di molti domini CH3. Esistono 4 sottoclassi di IgG nell’uomo ciascuna delle quali ha una differente affinità per C1q. La più affine è IgG3 , seguita dall’IgG1 e IgG2 mentre l’IgG4 non attiva il complemento.
. C1q è una proteina complessa, ricca in idrossiprolina, idrossilisina e glicina che per le sue caratteristiche elettroforetiche presenta delle analogie con le Ig, mentre per la sua struttura amminoacidica ricorda le proteine del collagene. La molecola C1q è costituita da 6 unità ognuna costituita da tre distinte subunità A,B,C. E’ possibile distinguere una parte centrale compatta , che lega C1r e C1 s , ed una struttura più esterna che termina con una forma a calice. La struttura a calice svolge la funzione di riconoscere e legare particolari siti presenti nelle Ig, i quali si evidenziano durante la formazione dell’immunocomplesso. La sub-unità C1q si deve legare ai domini CH2 di almeno due anticorpi IgG adiacenti . La sub-unità C1q si può legare ad un solo anticorpo IgM perche dotato di molti domini CH3. Esistono 4 sottoclassi di IgG nell’uomo ciascuna delle quali ha una differente affinità per C1q. La più affine è IgG3 , seguita dall’IgG1 e IgG2 mentre l’IgG4 non attiva il complemento.")
7
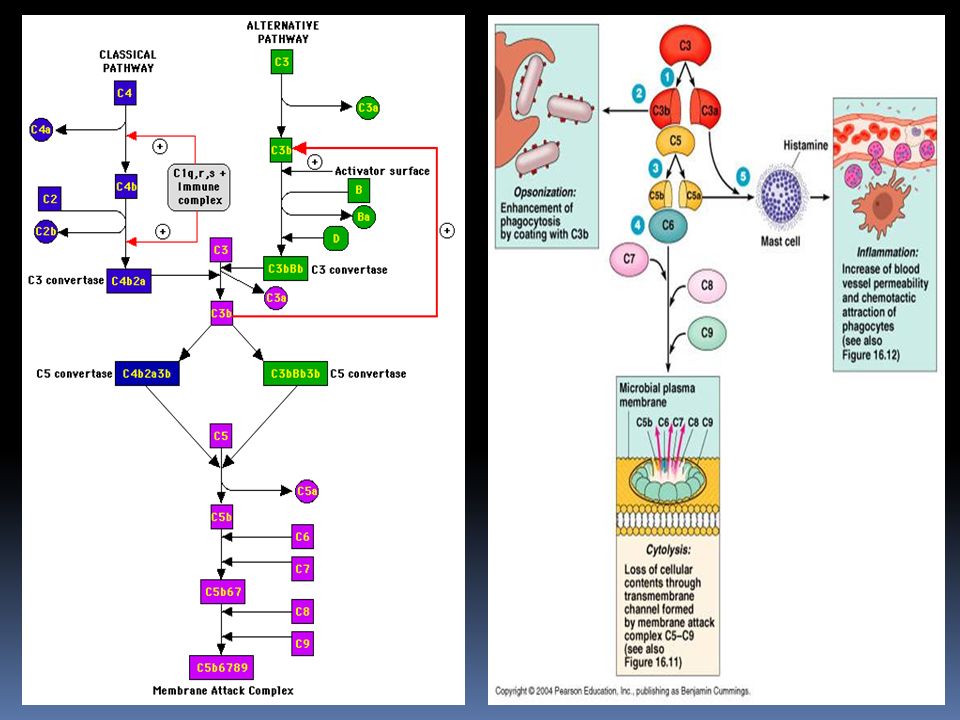
Via classica C1s inattivo ha un peso molecolare intorno a 90 kDa ed è costituito da una sola catena polipeptidica L’attivazione di C1s comporta la formazione di due catene una più grande (a) ed una più piccola (b) , in cui si trova il sito enzimatico necessario per l’attivazione dei componenti successivi . L’attività enzimatica è di tipo serino-esterasi, ed agisce su due substrati C4 e C2 che vengono scissi rispettivamente in C4a e C4b , e in C2a e C2b dando origine al complesso C4b2a, denominato C3 convertasi che ha la capacità di attivare il fattore C3.
 ed una più piccola (b) , in cui si trova il sito enzimatico necessario per l’attivazione dei componenti successivi . L’attività enzimatica è di tipo serino-esterasi, ed agisce su due substrati C4 e C2 che vengono scissi rispettivamente in C4a e C4b , e in C2a e C2b dando origine al complesso C4b2a, denominato C3 convertasi che ha la capacità di attivare il fattore C3.")
9
meccanismi di controllo
Via classica meccanismi di controllo Le reazioni che caratterizzano la via classica sono soggette al controllo di diverse proteine solubili quali : C1 inibitore ( C1 INH ) : questo fattore agisce su C1, bloccando l’attività enzimatica di C1s ( edema angioneurotico , dipende dall’assenza di questo fattore) C4bp ( C4 binding protein) : questo fattore si lega al C4b, bloccando la formazione del complesso C4b2a e permettendo , cosi, il legame del fattore H, capace a sua volta di inattivare , oltre che il C3b, anche il C4b. Fattore I : questo fattore inattiva il C3b, formando il C3b inattivo (iC3b). Questi meccanismi sono necessari per mantenere gli effetti dell’innesco della cascata del complemento solamente a livello del sito d’attivazione.
 : questo fattore agisce su C1, bloccando l’attività enzimatica di C1s ( edema angioneurotico , dipende dall’assenza di questo fattore) C4bp ( C4 binding protein) : questo fattore si lega al C4b, bloccando la formazione del complesso C4b2a e permettendo , cosi, il legame del fattore H, capace a sua volta di inattivare , oltre che il C3b, anche il C4b. Fattore I : questo fattore inattiva il C3b, formando il C3b inattivo (iC3b). Questi meccanismi sono necessari per mantenere gli effetti dell’innesco della cascata del complemento solamente a livello del sito d’attivazione.")
10
Via alternativa La via alternativa differisce dalla via classica essenzialmente per due motivi: viene attivata in assenza di anticorpi inizia dal componente C3 Costituisce un meccanismo di difesa anti-infettiva aspecifico ed immediato Può essere innescata da diverse sostanze : lipolisaccaride (LPS) dei batteri gram-negativi endotossine batteriche particelle virali Va tenuta in considerazione che il componente C3 và in contro ad una lenta scissione spontanea perche presenta al suo interno un legame molto instabile , un legame tioestere. Il C3 formatosi , alla presenza dei fattori d’iniziazione lega la proteina B, e forma il complesso C3bB. Se C3b non si lega alle strutture estranee viene trasformato in una molecola inattiva, il iC3b, importante per l’ opsonizzazione.
 dei batteri gram-negativi. endotossine batteriche. particelle virali. Va tenuta in considerazione che il componente C3 và in contro ad una lenta scissione spontanea perche presenta al suo interno un legame molto instabile , un legame tioestere. Il C3 formatosi , alla presenza dei fattori d’iniziazione lega la proteina B, e forma il complesso C3bB. Se C3b non si lega alle strutture estranee viene trasformato in una molecola inattiva, il iC3b, importante per l’ opsonizzazione.")
11
Via alternativa La proteina (fattore) B, una volta inserita nel complesso, è scissa dalla proteina D in Bb e Ba. Il complesso attivo C3bBb ( C3 convertasi della via alternativa) , potenziato dalla properdina (P) , è capace di scindere il C3 in C3a e C3b. C3b può rientrare nel ciclo, generando un meccanismo d’amplificazione del sistema , oppure legarsi al complesso precedente e dare origine, cosi , alla C5 convertasi della via alternativa (C3bBbC3b) La via alternativa è regolata dall’azione di due proteine solubili: la proteina H: fattore di controllo anche nella via classica . Si lega al C3b della C3 convertasi occupando il posto di Bb, bloccando , cosi, la successiva attivazione della cascata complementare. il fattore I : cliva il C3b, legato al fattore H, in iC3b, C3c e C3d.
 , potenziato dalla properdina (P) , è capace di scindere il C3 in C3a e C3b. C3b può rientrare nel ciclo, generando un meccanismo d’amplificazione del sistema , oppure legarsi al complesso precedente e dare origine, cosi , alla C5 convertasi della via alternativa (C3bBbC3b) La via alternativa è regolata dall’azione di due proteine solubili: la proteina H: fattore di controllo anche nella via classica . Si lega al C3b della C3 convertasi occupando il posto di Bb, bloccando , cosi, la successiva attivazione della cascata complementare. il fattore I : cliva il C3b, legato al fattore H, in iC3b, C3c e C3d.")
12
Via lectinica Viene attivata in assenza di Ab .
E’ un meccanismo di difesa aspecifico Viene indicata come MBL , ossia la via della lectina legante il mannosio La sua attivazione dipende dal riconoscimento non specifico di sostanze estranee (carboidrati). Presenta analogie strutturali alla via classica. La MBL è simile al C1q, infatti dopo che si è legata ad un microrganismo si unisce, per formare un complesso attivo , ad un enzima , detto MASP (MBL-associated serine-protease) , che è simile al C1r e al C1s della via classica. Il complesso MBL-MASP cliva il C4 e il C2 e porta alla formazione di una C3 convertasi che ha come conseguenza la formazione della C5 convertasi.
. Presenta analogie strutturali alla via classica. La MBL è simile al C1q, infatti dopo che si è legata ad un microrganismo si unisce, per formare un complesso attivo , ad un enzima , detto MASP (MBL-associated serine-protease) , che è simile al C1r e al C1s della via classica. Il complesso MBL-MASP cliva il C4 e il C2 e porta alla formazione di una C3 convertasi che ha come conseguenza la formazione della C5 convertasi.")
13
Formazione del complesso d’attacco alla membrana
( MAC ) La C5 convertasi provoca la scissione del C5 in C5a e C5b. La C5a liberato contribuisce con le sue capacità anafilattiche e chemiotattiche alla risposta infiammatoria locale La C5b si lega al C6 e al C7 per formare il complesso C5b-7, che presentano la capacità di legarsi alle membrane biologiche. Il complesso C5b-7 lega una molecola del fattore C8 e circa 10 molecole di C9, formando il complesso d’attacco alla membrana (MAC), il C5b-9. Il complesso d’attacco alla membrana (MAC) ha attività litica sulle membrane, causando la formazione di pori di circa 10 nm di diametro Attraverso i pori avviene il passaggio d’acqua e di Sali che portano alla lisi osmotica della cellula o del microrganismo. La proteina S si lega al complesso C5b-9, la quale blocca l’attività di lisi e quindi regola l’attività biologica del MAC.
 La C5 convertasi provoca la scissione del C5 in C5a e C5b. La C5a liberato contribuisce con le sue capacità anafilattiche e chemiotattiche alla risposta infiammatoria locale. La C5b si lega al C6 e al C7 per formare il complesso C5b-7, che presentano la capacità di legarsi alle membrane biologiche. Il complesso C5b-7 lega una molecola del fattore C8 e circa 10 molecole di C9, formando il complesso d’attacco alla membrana (MAC), il C5b-9. Il complesso d’attacco alla membrana (MAC) ha attività litica sulle membrane, causando la formazione di pori di circa 10 nm di diametro. Attraverso i pori avviene il passaggio d’acqua e di Sali che portano alla lisi osmotica della cellula o del microrganismo. La proteina S si lega al complesso C5b-9, la quale blocca l’attività di lisi e quindi regola l’attività biologica del MAC.")
14
Fattori regolatori Esistono molecole di membrana ad attività regolatoria tra cui: fattore accellerante il decadimento ( DAF) e la proteina cofattore di membrana (MCP), che agiscono sulla C3 convertasi della via classia e della via alternativa. fattore di restrizione omologa (HRF) e l’ inibitore di membrana della reazione di lisi (MIRL/CD59), che lega il complesso C5b-8 e blocca il legame del C9 alle cellule autologhe.
 e la proteina cofattore di membrana (MCP), che agiscono sulla C3 convertasi della via classia e della via alternativa. fattore di restrizione omologa (HRF) e l’ inibitore di membrana della reazione di lisi (MIRL/CD59), che lega il complesso C5b-8 e blocca il legame del C9 alle cellule autologhe.")
Presentazioni simili



 Costituito da una parte liquida: plasma e.>")







>")







