Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Riproduzione e sviluppo

2
Dimorfismo sessuale Gli esseri umani sono sessualmente dimorfici, cioè l’uomo e la donna sono fisicamente diversi Gli organi sessuali maschili e femminili consistono di tre insiemi di strutture: le gonadi, i genitali interni, i genitali esterni

3
Le gonadi Le gonadi sono gli organi che producono i gameti, le cellule riproduttive (ovulo e spermatozoo), che si uiniscono per formare un nuovo individuo Le gonadi maschili sono i testicoli e producono gli spermatozoi Le gonadi femminili sono le ovaie e producono le uova o ovuli Le cellule gonadiche indifferenziate destinate a produrre ovuli o spermatozoi sono definite cellule germinali
, che si uiniscono per formare un nuovo individuo. Le gonadi maschili sono i testicoli e producono gli spermatozoi. Le gonadi femminili sono le ovaie e producono le uova o ovuli. Le cellule gonadiche indifferenziate destinate a produrre ovuli o spermatozoi sono definite cellule germinali.")
4
I genitali I genitali interni consistono di ghiandole e dotti accessori che connettono le gonadi con l’ambiente esterno I genitali esterni includono tutte le strutture riproduttive esterne

5
La fecondazione La riproduzione dei singoli individui, ai fini del mantenimento della specie, avviene per effetto della fecondazione, processo per il quale uno spermatozoo penetra in una cellula uovo dando inizio allo sviluppo di un nuovo individuo (zigote) Lo spermatozoo e l’uovo, con il loro contributo cromosomico, conferiranno al nuovo individuo caratteristiche somatiche e sessuali proprie
 Lo spermatozoo e l’uovo, con il loro contributo cromosomico, conferiranno al nuovo individuo caratteristiche somatiche e sessuali proprie.")
6
I cromosomi Oogoni e spermatogoni, cellule primordiali dei gameti, contengono 46 cromosomi (23 coppie, corredo diploide) e, in particolare: L’oogonio presenta 44 autosomi e 2 gonosomi (cromosomi sessuali) identici XX Lo spermatogonio presenta 44 autosomi e 2 gonosomi diversi X e Y
 identici XX. Lo spermatogonio presenta 44 autosomi e 2 gonosomi diversi X e Y.")
7
I cromosomi Gli autosomi contengono l’informazione per lo sviluppo del corpo umano e delle sue caratteristiche variabili (colore degli occhi, gruppi sanguigni, ecc.) I gonosomi contengono i geni che regolano lo sviluppo degli organi sessuali interni ed esterni Il cromosoma X è più grande del cromosoma Y e contiene molti più geni
 I gonosomi contengono i geni che regolano lo sviluppo degli organi sessuali interni ed esterni. Il cromosoma X è più grande del cromosoma Y e contiene molti più geni.")
8
La meiosi Prima della fecondazione oogoni e spermatogoni vanno incontro alla meiosi e ogni cellula figlia riceverà soltanto 23 cromosomi (corredo aploide) uno da ognuna delle 23 paia di cromosomi Al termine della riduzione cromosomica,quindi, tutti gli oociti avranno 22 autosomi e 1 gonosoma X, gli spermatozoi avranno 22 autosomi e un gonosoma X o Y
 uno da ognuna delle 23 paia di cromosomi. Al termine della riduzione cromosomica,quindi, tutti gli oociti avranno 22 autosomi e 1 gonosoma X, gli spermatozoi avranno 22 autosomi e un gonosoma X o Y.")
9
La gametogenesi

10
Lo zigote L’uovo, che contiene sempre l’autosoma X, potrà essere fecondato da 2 tipi di spermatozoi: Spermatozoo contenente l’autosoma X, e si avrà uno zigote con autosomi XX che si svilupperà in una femmina Spermatozoo contenente l’autosoma Y, e si avrà uno zigote con autosomi XY che si svilupperà in un maschio

11
Patologie ereditarie legate al cromosoma X
Molti geni legati al cromosoma X (X-linked genes) non corrispondono ad alcun gene sul cromosoma Y che è molto più piccolo. Le femmine hanno sempre due copie dei geni che si trovano su tale cromosoma, e la loro espressione segue il modello classico di dominanza e recessione genica. I maschi, avendo un solo cromosoma X, mostrano sempre i tratti associati ai geni che si trovano su questo cromosoma. Quindi se il gene ereditato dalla madre è difettoso, il figlio maschio manifesterà la mutazione (distrofia muscolare di Duchenne, emofilia, daltonismo)
 non corrispondono ad alcun gene sul cromosoma Y che è molto più piccolo. Le femmine hanno sempre due copie dei geni che si trovano su tale cromosoma, e la loro espressione segue il modello classico di dominanza e recessione genica. I maschi, avendo un solo cromosoma X, mostrano sempre i tratti associati ai geni che si trovano su questo cromosoma. Quindi se il gene ereditato dalla madre è difettoso, il figlio maschio manifesterà la mutazione (distrofia muscolare di Duchenne, emofilia, daltonismo)")
12
Anomalie nella distribuzione dei cromosomi sessuali
X0 = Sindrome di Turner. Individui con aspetto femminile ma genitali esterni ed interni ambigui (gonade incompleta) XXY = Sindrome di Klinefelter. Individui con genitali esterni ed interni maschili, ma con testicoli ridotti, sviluppo anomalo dei tubuli seminiferi e deficiente spermatogenesi
 XXY = Sindrome di Klinefelter. Individui con genitali esterni ed interni maschili, ma con testicoli ridotti, sviluppo anomalo dei tubuli seminiferi e deficiente spermatogenesi.")
13
I processi di sessualizzazione
In base alla distribuzione cromosomica, si ha la differenziazione del sesso e la determinazione degli organi sessuali di un individuo che costituiscono l’insieme dei processi di sessualizzazione che coprono tre periodi: 1) sviluppo delle gonadi 2) sviluppo dei genitali interni 3) sviluppo dei genitali esterni
 sviluppo delle gonadi. 2) sviluppo dei genitali interni. 3) sviluppo dei genitali esterni.")
14
La differenziazione sessuale
Le strutture riproduttive non iniziano a differenziarsi prima della settima settimana Fino ad allora i tessuti embrionali sono bipotenziali e non possono essere identificati come maschili né come femminili

15
La gonade bipotenziale consta di una porzione corticale esterna e di una midollare interna
I genitali interni bipotenziali sono costituiti dai dotti di Wolff (derivati dal rene embrionale) nella midollare,e dai dotti di Muller nella corticale A seguito di un adeguato segnale di sviluppo, la midollare si svilupperà in testicolo; in assenza di tale segnale, la corticale si svilupperà in tessuto ovarico
 nella midollare,e dai dotti di Muller nella corticale. A seguito di un adeguato segnale di sviluppo, la midollare si svilupperà in testicolo; in assenza di tale segnale, la corticale si svilupperà in tessuto ovarico.")
16
I dotti di Wolff e di Müller avranno destino diverso. In particolare:
Nell’embrione geneticamente femmina, il sistema dei dotti di Müller si differenzierà in tube uterine, utero, cervice e vagina e i dotti di Wolff regrediranno Nell’embrione geneticamente maschio i dotti di Wolff si svilupperanno in epididimo, deferente, vescicole seminali e dotto eiaculatore e i dotti di Müller regrediranno

17
Sviluppo delle gonadi Sul cromosoma Y è presente un gene detto SRY o regione determinante il sesso da cui, appunto, dipende la determinazione del sesso In assenza del gene SRY e dei suoi prodotti, le gonadi si sviluppano in ovaia In presenza di tale gene funzionale, le gonadi bipotenziali si sviluppano in testicoli

18
Il gene SRY Il gene SRY induce la sintesi della proteina SRY o fattore di determinazione testicolare che si lega al DNA attivando altri geni I prodotti proteici di questi geni dirigono lo sviluppo della midollare gonadica in testicolo Quindi lo sviluppo testicolare NON richiede ormoni sessuali maschili (testosterone) che non può essere secreto finché le gonadi non si sviluppano in testicoli
 che non può essere secreto finché le gonadi non si sviluppano in testicoli.")
19
I genitali esterni bipotenziali consistono di un tubercolo genitale, di pieghe uretrali, del solco uretrale e dei rigonfiamenti labioscrotali Queste strutture si differenzieranno nelle strutture riproduttive maschili e femminili nelle successive fasi di sviluppo

20
Sviluppo dei genitali interni ed esterni maschili
Quando i testicoli si differenziano (tra la sesta e la settima settimana), essi inziano a secernere tre ormoni che influenzano lo sviluppo dei genitali maschili interni ed esterni: Le cellule del Sertoli secernono l’ormone glicoproteico antimulleriano (o sostanza inibente il dotto di Müller, AMH) Le cellule di Leydig secernono testosterone e il suo derivato deidrotestosterone (DTH), ormoni steroidei dominanti nel maschio (entrambi si legano allo stesso recettore ma con effetto diverso)
, essi inziano a secernere tre ormoni che influenzano lo sviluppo dei genitali maschili interni ed esterni: Le cellule del Sertoli secernono l’ormone glicoproteico antimulleriano (o sostanza inibente il dotto di Müller, AMH) Le cellule di Leydig secernono testosterone e il suo derivato deidrotestosterone (DTH), ormoni steroidei dominanti nel maschio (entrambi si legano allo stesso recettore ma con effetto diverso)")
21
Sviluppo maschile In particolare, nell’embrione geneticamente maschio:
Dotto di Müller in fase di degenerazione Stadio dello sviluppo sessuale indifferenziato In particolare, nell’embrione geneticamente maschio: L’ormone antimulleriano (AMH) causa la regressione dei dotti di Müller Il testosterone converte i dotti di Wolff nelle strutture accessorie maschili (epididimo, vasi deferenti, vescicole seminali) Più tardi nello sviluppo fetale, il testosterone controlla la migrazione del testicolo dall’addome nello scroto o sacco scrotale
 causa la regressione dei dotti di Müller. Il testosterone converte i dotti di Wolff nelle strutture accessorie maschili (epididimo, vasi deferenti, vescicole seminali) Più tardi nello sviluppo fetale, il testosterone controlla la migrazione del testicolo dall’addome nello scroto o sacco scrotale.")
22
Differenziazione dei genitali esterni maschili
La differenziazione dei genitali esterni è controllata principalmente dal DTH

23
Nell’embrione geneticamente femmina, in assenza del gene SRY non si sviluppa il testicolo e viene a mancare l’inibizione da parte dell’AMH testicolare, quindi: I dotti di Müller si sviluppano nella porzione superiore della vagina, nell’utero e nelle tube di Falloppio o ovidotti I dotti di Wolff degenerano Verso la 12° settimana gli oogoni cominciano la divisione meiotica per formare oociti segnando l’inizio della differenziazione ovarica che si completerà tra la 20° e la 25° settimana Sviluppo femminile Stadio dello sviluppo sessuale indifferenziato

24
Differenziazione dei genitali esterni femminili
Senza DHT i genitali esterni assumono caratteristiche femminili

25
Riassunto

26
Deficit enzimatici e recettoriali nel maschio
Individui geneticamente maschi che mancano dei recettori per gli androgeni sono affetti da sindrome di femminilizzazione testicolare: La presenza del cromosoma Y porta allo sviluppo dei testicoli e alla regressione del dotto di Müller,ma la mancanza dei recettori degli androgeni non permette lo sviluppo dei dotti di Wolff. Quindi mancano i genitali interni Individui geneticamente maschi con deficit dell’enzima 5-a-reduttasi (converte il testosterone in DHT) mostrano una ridotta mascolinizzazione dei genitali esterni (riduzione di grado diverso a seconda del deficit) senza interferire con la formazione di testicoli normali
 mostrano una ridotta mascolinizzazione dei genitali esterni (riduzione di grado diverso a seconda del deficit) senza interferire con la formazione di testicoli normali.")
27
Deficit enzimatici nella femmina
L’eccessiva presenza di androgeni nelle prime fasi di sviluppo embrionale porta a differenziazione dei genitali esterni maschili Ciò accade quando la madre secerne un eccesso di androgeni o quando individui geneticamente femmina sono carenti degli enzimi surrenalici 11-idrossilasi o 21-idrossilasi necessari per la sintesi dei gluco- e mineralcorticoidi. In assenza di questi enzimi, il surrene sintetizza androgeni dal colesterolo L’ipersecrezione di androgeni porta alla sindrome adrenogenitale (iperplasia surrenalica congenita) con vario grado di mascolinizzazione dei genitali esterni Una severa mascolinizzazione dei genitali esterni femminili porta allo pseudoermafroditismo femminile che può indurre all’errore nell’identificazione del sesso del neonato
 con vario grado di mascolinizzazione dei genitali esterni. Una severa mascolinizzazione dei genitali esterni femminili porta allo pseudoermafroditismo femminile che può indurre all’errore nell’identificazione del sesso del neonato.")
28
Modelli base della riproduzione
I testicoli maschili e le ovaie femminili producono entrambi sia ormoni, sia gameti (quindi hanno secrezione sia endocrina, sia esocrina) Gli ovuli (gameti femminili) sono tra le cellule più voluminose dell’organismo, non sono mobili e sono spostati lungo l’apparato riproduttivo dalle correnti create dalla contrazione della muscolatura liscia o dal movimento delle ciglia Gli spermatozoi (gameti maschili) sono piccoli, molto mobili e sono le uniche cellule dell’organismo dotate di flagelli che permettono loro di risalire lungo l’apparato riproduttivo femminile alla ricerca dell’uovo da fecondare
 Gli ovuli (gameti femminili) sono tra le cellule più voluminose dell’organismo, non sono mobili e sono spostati lungo l’apparato riproduttivo dalle correnti create dalla contrazione della muscolatura liscia o dal movimento delle ciglia. Gli spermatozoi (gameti maschili) sono piccoli, molto mobili e sono le uniche cellule dell’organismo dotate di flagelli che permettono loro di risalire lungo l’apparato riproduttivo femminile alla ricerca dell’uovo da fecondare.")
29
Apparato genitale maschile
Organi sessuali primari: Gonadi o testicoli o didimi Organi sessuali secondari: Vie spermatiche (tubuli retti e rete testis, epididimi, canali deferenti, dotti eiaculatori, uretra comune); ghiandole annesse (vescichette seminali, prostata, gh. bulbo-uretrali); genitali esterni (pene, borsa scrotale o scroto); organi rudimentali (appendice del testicolo e dell’epididimo, paradidimo, condotti aberranti) Apparato genitale maschile
; ghiandole annesse (vescichette seminali, prostata, gh. bulbo-uretrali); genitali esterni (pene, borsa scrotale o scroto); organi rudimentali (appendice del testicolo e dell’epididimo, paradidimo, condotti aberranti) Apparato genitale maschile.")
30
Gli organi sessuali primari
Testicolo o didimo: organo pari a secrezione sia esterna (spermatozoi) sia interna (ormoni androgeni, il più importante dei quali è il testosterone) La secrezione esterna o spermatogenesi si verifica a temperatura inferiore a quella corporea, per cui, fin dal 7° mese di vita intrauterina, i testicoli iniziano la loro migrazione verso le borse scrotali, dove la temperatura è nettamente inferiore ( almeno 2°C) La secrezione ormonale, invece, non è influenzata da variazioni di temperatura
 sia interna (ormoni androgeni, il più importante dei quali è il testosterone) La secrezione esterna o spermatogenesi si verifica a temperatura inferiore a quella corporea, per cui, fin dal 7° mese di vita intrauterina, i testicoli iniziano la loro migrazione verso le borse scrotali, dove la temperatura è nettamente inferiore ( almeno 2°C) La secrezione ormonale, invece, non è influenzata da variazioni di temperatura.")
31
Il testicolo E’ una struttura avvolta dalla tonaca albuginea, di natura fibrosa ricca di terminazioni nocicettive, a sua volta ricoperta dalla tunica sierosa (tonaca vaginale) Dalla tonaca albuginea partono sottili sepimenti che dividono il testicolo in lobuli piramidali, ciascuno dei quali contiene 2-4 tubuli seminiferi ( cm ciascuno) che si riuniscono nella rete testis, poi nei dotti efferenti, quindi nell’epididimo
 Dalla tonaca albuginea partono sottili sepimenti che dividono il testicolo in lobuli piramidali, ciascuno dei quali contiene 2-4 tubuli seminiferi ( cm ciascuno) che si riuniscono nella rete testis, poi nei dotti efferenti, quindi nell’epididimo.")
32
Testa dell’epididimo Coda dell’epididimo

33
All’interno dei lobuli, i tubuli seminiferi sono circondati da tessuto fibrillare lasso, interstiziale, le cui cellule di Leydig sono riunite a formare cordoni cellulari a funzione endocrina I tubuli seminiferi ( μm di diametro, m di lunghezza complessiva) sono costituiti da cellule del Sertoli (in una sola fila) e più strati di epitelio germinativo o seminale che poggiano su una membrana basale
 sono costituiti da cellule del Sertoli (in una sola fila) e più strati di epitelio germinativo o seminale che poggiano su una membrana basale.")
34
Le cellule interstiziali di Leydig
Rappresentano la parte endocrina: secernono androgeni, soprattutto testosterone ma anche diidrotestosterone e androstenedione Si differenziano dalle cellule mesenchimali indifferenziate solo in presenza di LH adenoipofisario o di gonadotropina corionica (hCG) prodotta dalla placenta: i recettori sono membranari e il II messaggero è il cAMP Nucleo grande, citoplasma abbondante, ricche di lipidi ed enzimi, tra cui quelli che catalizzano la steroidogenesi
 prodotta dalla placenta: i recettori sono membranari e il II messaggero è il cAMP. Nucleo grande, citoplasma abbondante, ricche di lipidi ed enzimi, tra cui quelli che catalizzano la steroidogenesi.")
35
Le cellule del Sertoli Cellule grandi con digitazioni di membrana che sembrano formare un sincizio (sincizio di Sertoli) 2 tipi di recettori Recettori di membrana specifici per l’FSH adenoipofisario (adenilato ciclasi– cAMP) Recettori citosolici, specifici per gli androgeni (ABP- proteina legante gli androgeni- soprattutto testosterone) Modesta attività endocrina stereidogenetica. Sintesi di inibina Funzione trofica sull’epitelio germinativo cui forniscono lipidi e glicogeno attraverso gap junctions; forniscono anche il liquido per il trasporto nel lume. Il distacco degli spermatozoi dalle cellule del Sertoli viene definito spermiazione La loro stretta giustapposizione impedisce la penetrazione dai capillari circostanti di proteine (es immunoglobuline) che potrebbero interferire con lo sviluppo degli spermatogoni (Barriera ematotesticolare)
 Recettori citosolici, specifici per gli androgeni (ABP- proteina legante gli androgeni- soprattutto testosterone) Modesta attività endocrina stereidogenetica. Sintesi di inibina. Funzione trofica sull’epitelio germinativo cui forniscono lipidi e glicogeno attraverso gap junctions; forniscono anche il liquido per il trasporto nel lume. Il distacco degli spermatozoi dalle cellule del Sertoli viene definito spermiazione. La loro stretta giustapposizione impedisce la penetrazione dai capillari circostanti di proteine (es immunoglobuline) che potrebbero interferire con lo sviluppo degli spermatogoni (Barriera ematotesticolare)")
37
Le cellule dell’epitelio germinativo
Sono rappresentate dagli spermatogoni e dalle cellule che da essi derivano in via maturativa: spermatociti di I e di II ordine spermatidi spermatozoi, cioè i gameti maturi Tale trasformazione rappresenta il ciclo della spermatogenesi e si compie in gg

38
La gametogenesi Il periodo della riproduzione dei gameti o gametogenesi è molto diverso nel maschio e nella femmina Le donne alla nascita posseggono già tutti gli ovuli, o ovociti, e non ne produrranno altri nel corso della vita. Durante il periodo fertile gli ovuli maturano e sono rilasciati dalle ovaie approssimativamente una volta al mese per circa 40 anni. Poi il ciclo riproduttivo cessa (menopausa) Gli uomini, invece, producono continuamente spermatozoi da quando raggiungono la maturità riproduttiva. La produzione diminuisce con l’età ma non cessa mai completamente.
 Gli uomini, invece, producono continuamente spermatozoi da quando raggiungono la maturità riproduttiva. La produzione diminuisce con l’età ma non cessa mai completamente.")
39
Gametogenesi maschile
Alla nascita: i testicoli contengono solo cellule germinali immature Dopo la nascita: le gonadi restano inattive fino alla pubertà, quando maturano Alla pubertà: le cellule germinali dette spermatogoni, hanno due possibilità. Alcune vanno in mitosi per tutta la vita riproduttiva del maschio, altre vanno in meiosi e diventano spermatociti primari Gli spermatociti primari daranno due spermatociti secondari alla prima divisione meiotica e ciascuno di questi alla seconda divisione meiotica darà due spermatidi aploidi che matureranno in spermatozoi (1 spermatocita primario diploide= 4 spermatozoi aploidi)
")
41
Ormoni essenziali per la spermatogenesi
Testosterone - cell di Leydig: essenziale per la divisione e lo sviluppo delle cell germinali Ormone luteinizzante (LH) – adenoipofisi: stimola le cell. di Leydig a secernere testosterone Ormone follicolo stimolante (FSH) – adenoipofisi: stimola le cell. del Sertoli a mediare la conversione di spermatidi in spermatozoi e a secernere inibina che inibisce la secrezione di FSH con meccanismo a feed-back Estrogeni – cell. del Sertoli: dal testosterone per azione dell’FSH: ruolo poco chiaro nella spermatogenesi (sicuramente implicati nella maturazione dell’osso) Proteina legante androgeni – cell. del Sertoli: lega il testosterone e gli estrogeni e li trasporta nel liquido dei tubuli seminiferi Ormone della crescita (GH) – adenoipofisi: promuove la divisione degli spermatogoni ed è necessario per il normale svolgimento delle funzioni metaboliche di base del testicolo
 – adenoipofisi: stimola le cell. di Leydig a secernere testosterone. Ormone follicolo stimolante (FSH) – adenoipofisi: stimola le cell. del Sertoli a mediare la conversione di spermatidi in spermatozoi e a secernere inibina che inibisce la secrezione di FSH con meccanismo a feed-back. Estrogeni – cell. del Sertoli: dal testosterone per azione dell’FSH: ruolo poco chiaro nella spermatogenesi (sicuramente implicati nella maturazione dell’osso) Proteina legante androgeni – cell. del Sertoli: lega il testosterone e gli estrogeni e li trasporta nel liquido dei tubuli seminiferi. Ormone della crescita (GH) – adenoipofisi: promuove la divisione degli spermatogoni ed è necessario per il normale svolgimento delle funzioni metaboliche di base del testicolo.")
43
Maturazione degli spermatozoi
Poiché alla fine della spermatogenesi gli spermatozoi sono immobili, ma devono raggiungere l’epididimo per completare la maturazione, essi vengono spinti verso la rete testis dall’epitelio ciliato presente nel lume e dalla muscolatura liscia dei condotti. Gli spermatozoi impiegano gg per attraversare tutto l’epididimo (4-5 m) e giungere nell’eiaculato In questo percorso le cellule diventano mobili e acquistano la capacità di fecondare l’ovulo Gli spermatozoi prelevati direttamente dal testicolo non sono fertili In assenza di eiaculazione, essi degenerano e si liquefanno (non passano mai nelle vescicole seminali)
 e giungere nell’eiaculato. In questo percorso le cellule diventano mobili e acquistano la capacità di fecondare l’ovulo. Gli spermatozoi prelevati direttamente dal testicolo non sono fertili. In assenza di eiaculazione, essi degenerano e si liquefanno (non passano mai nelle vescicole seminali)")
45
Lo spermatozoo maturo Durante la maturazione, il nucleo diventa sempre più piccolo e compatto, andando a formare la testa dello spermio maturo Un piccolo corpo citoplasmatico, il centriolo,emette un fascio di fibre e, insieme al sottile velo di citoplasma che lo avvolge, diventa la coda Il citoplasma si riduce ad un velo che avvolge testa, tratto intermedio (con spirale di mitocondri = fonte energetica) e coda Nella testa è contenuto l’acrosoma (formatosi nell’apparato di Golgi) contenente enzimi proteolitici e la ialuronidasi, che liquefa l’ac. ialuronico del muco cervicale dell’utero e quello della zona pellucida dell’uovo
 e coda. Nella testa è contenuto l’acrosoma (formatosi nell’apparato di Golgi) contenente enzimi proteolitici e la ialuronidasi, che liquefa l’ac. ialuronico del muco cervicale dell’utero e quello della zona pellucida dell’uovo.")
46
Il movimento La coda dello spermatozoo è composta da 9 fibrille disposte attorno ad altre 2 centrali: insieme formano l’assonema Solo le fibrille esterne si contraggono con movimenti flagellari (velocità = 20 μm/sec = 70 mm/h) L’energia è fornita dall’ATP sintetizzato dai mitocondri
 L’energia è fornita dall’ATP sintetizzato dai mitocondri.")
47
Il liquido seminale Poiché gli spermi hanno scarso citoplasma, il liquido seminale fornisce loro il nutrimento (la concentrazione di fruttosio è maggiore della concentrazione di glucosio ematico) Impartisce mobilità Liquefa il muco del canale cervicale dell’utero Sensibilizza la muscolatura dell’utero grazie al suo contenuto in prostaglandine prodotte dalla prostata Protegge gli spermi dall’alta temperatura delle vie genitali femminili che ne eleva l’attività ma ne abbrevia la vita perché ne innalza il metabolismo. Nelle vie genitali maschili gli spermatozoi sopravvivono per settimane, ma in quelle femminili, per non più di 24-48h. A temperature più basse lo sperma può essere conservato per settimane o, sotto i 100°, anche per anni
 Impartisce mobilità. Liquefa il muco del canale cervicale dell’utero. Sensibilizza la muscolatura dell’utero grazie al suo contenuto in prostaglandine prodotte dalla prostata. Protegge gli spermi dall’alta temperatura delle vie genitali femminili che ne eleva l’attività ma ne abbrevia la vita perché ne innalza il metabolismo. Nelle vie genitali maschili gli spermatozoi sopravvivono per settimane, ma in quelle femminili, per non più di 24-48h. A temperature più basse lo sperma può essere conservato per settimane o, sotto i 100°, anche per anni.")
48
Il liquido seminale E’ formato da spermatozoi e liquido proveniente da deferenti, prostata, gh. Di Cowper ma soprattutto dalle vescicole seminali (ca 60%). Questo è l’ultimo ad essere eiaculato e lava via il materiale spermatico dal dotto eiaculatore e dall’uretra Il liquido prostatico gli conferisce pH alcalino (7,5), aspetto lattiginoso, proprietà coagulanti: l’enzima coagulante agisce sul fibrinogeno emesso dalle vescicole seminali dopo ca 5 min dall’eiaculazione, formando un debole coagulo che serve a trattenere lo sperma nel collo dell’utero. Il coagulo si dissolve dopo ca 15 min (fibrinolisina, dalla profibrinolisina prostatica) e gli spermatozoi diventano più mobili Il vol medio di una eiaculazione nell’adulto è di 2-4 ml con milioni di spermatozoi. Nella sua vita l’uomo produce ca 400 miliardi di spermatozoi contro le 400 uova della donna
. Questo è l’ultimo ad essere eiaculato e lava via il materiale spermatico dal dotto eiaculatore e dall’uretra. Il liquido prostatico gli conferisce pH alcalino (7,5), aspetto lattiginoso, proprietà coagulanti: l’enzima coagulante agisce sul fibrinogeno emesso dalle vescicole seminali dopo ca 5 min dall’eiaculazione, formando un debole coagulo che serve a trattenere lo sperma nel collo dell’utero. Il coagulo si dissolve dopo ca 15 min (fibrinolisina, dalla profibrinolisina prostatica) e gli spermatozoi diventano più mobili. Il vol medio di una eiaculazione nell’adulto è di 2-4 ml con milioni di spermatozoi. Nella sua vita l’uomo produce ca 400 miliardi di spermatozoi contro le 400 uova della donna.")
49
Composizione dello sperma

50
Cause di sterilità dello sperma
Contiene meno di 20 milioni di spermatozoi Il 20% degli spermatozoi sono anormali (testa o coda) Scarsa motilità Temperatura dei testicoli troppo alta (causa temporanea o conseguente a criptorachidismo, che può essere dovuto a insufficiente quantità di testosterone) Distruzione dell’epitelio dei tubuli seminiferi (parotite)
 Scarsa motilità. Temperatura dei testicoli troppo alta (causa temporanea o conseguente a criptorachidismo, che può essere dovuto a insufficiente quantità di testosterone) Distruzione dell’epitelio dei tubuli seminiferi (parotite)")
52
Capacitazione degli spermatozoi I
Gli spermatozoi acquistano capacità di fecondazione solo nel tratto genitale femminile mediante i processi di capacitazione: Le secrezioni uterine e delle tube di Falloppio asportano i fattori inibitori che avevano represso l’attività degli spermatozoi nelle vie genitali maschili Nei tubuli seminiferi sono presenti vescicole flottanti contenenti colesterolo che viene continuamente ceduto alla membrana dell’acrosoma, rendendola più resistente. Dopo l’eiaculazione, nelle vie genitali femminili, gli spermi si allontanano dalle vescicole e perdono il colesterolo in eccesso, e la membrana sull’acrosoma diventa molto meno resistente

53
Capacitazione degli spermatozoi II
La membrana della testa diventa più permeabile agli ioni Ca++, che rendono più efficaci ed energici i movimenti del flagello Gli ioni Ca++ modificano la membrana dell’acrosoma che si fonde con quella della testa dello spermatozoo L’acrosoma potrà liberare i suoi enzimi quando penetrerà nella zona pellucida dell’uovo Entro 30 min la membrana dello spermatozoo e dell’oocita si fondono e avviene la fecondazione

54
Gli organi sessuali secondari
Gli organi sessuali secondari (vie spermatiche con le loro ghiandole annesse e genitali esterni) sono quindi coinvolti nella produzione, maturazione e trasporto degli spermatozoi L’attività endocrina dei testicoli promuove la maturazione degli organi sessuali secondari e dei caratteri sessuali primari e secondari sino alla pubertà e li mantiene fino alla vecchiaia
 sono quindi coinvolti nella produzione, maturazione e trasporto degli spermatozoi. L’attività endocrina dei testicoli promuove la maturazione degli organi sessuali secondari e dei caratteri sessuali primari e secondari sino alla pubertà e li mantiene fino alla vecchiaia.")
55
I caratteri sessuali I caratteri sessuali primari sono rappresentati dagli organi sessuali interni e dai genitali esterni I caratteri sessuali secondari sono quei tratti che distinguono i maschi dalle femmine: la forma del corpo, la distribuzione dei peli corporei, l’ispessimento delle corde vocali, la struttura delle ossa del bacino….

56
I caratteri sessuali sono anch’essi sotto controllo ormonale, quindi, la castrazione effettuata
prima della pubertà comporta regressione atrofica degli organi e modificazione dell’accrescimento corporeo (massa muscolare ridotta, tessuto adiposo più abbondante, ginecomastia). dopo la pubertà determina regressione dei caratteri sessuali secondari
. dopo la pubertà determina regressione dei caratteri sessuali secondari.")
57
Attività steroidogenetica
La secrezione ormonale è quasi del tutto a carico delle cell. di Leydig integrata da quella dei tubuli seminiferi e delle cell. del Sertoli

58
La secrezione endocrina del testicolo
La secrezione interna del testicolo comprende Androgeni (testosterone, diidrotestosterone, androstenedione) Estrogeni (17β-estradiolo ed estrone) In condizioni fisiologiche prevale la secrezione di androgeni Le cellule del Sertoli producono La inibina che inibisce la secrezione di FSH La proteina che lega gli androgeni (PLA o ABP) E contengono l’aromatasi, enzima che converte gli androgeni in estrogeni
 Estrogeni (17β-estradiolo ed estrone) In condizioni fisiologiche prevale la secrezione di androgeni. Le cellule del Sertoli producono. La inibina che inibisce la secrezione di FSH. La proteina che lega gli androgeni (PLA o ABP) E contengono l’aromatasi, enzima che converte gli androgeni in estrogeni.")
59
Il testosterone E’ prodotto, a partire dall’acetato attraverso il colesterolo, dalle cellule di Leydig (che costituiscono il 20% della massa testicolare dell’adulto) Queste cell. sono abbondanti nell’adulto e nel neonato (dove sono stimolate dalle gonadotropine corioniche) mentre sono scarse nel bambino. La sintesi è uguale a quella che avviene nel surrene e nell’ovaio Nei tessuti bersaglio il testosterone viene convertito in diidrotestosterone, più attivo
 Queste cell. sono abbondanti nell’adulto e nel neonato (dove sono stimolate dalle gonadotropine corioniche) mentre sono scarse nel bambino. La sintesi è uguale a quella che avviene nel surrene e nell’ovaio. Nei tessuti bersaglio il testosterone viene convertito in diidrotestosterone, più attivo.")
60
aromatasi 5α-reduttasi

61
Caratteristiche ormonali
La secrezione è di ca 7mg/die nell’uomo e di ca 0,15 mg/die nella donna E’ max nelle prime ore del mattino e nei mesi di maggio e novembre Circola legato per il 40-45% alla SHBG (sex hormon binding protein), per il 50-55% ad albumine, α1-glicoproteina acida, transcortina, mentre il 2% è libero Emivita: 30 min libero, 1h in forma legata
, per il 50-55% ad albumine, α1-glicoproteina acida, transcortina, mentre il 2% è libero. Emivita: 30 min libero, 1h in forma legata.")
62
Recettore: nucleare per la forma libera, che dopo penetrazione viene convertita nella forma più attiva (diidrotestosterone, DHT), membranario (con cAMP come II messaggero) per la forma legata all’SHBG Degradazione: quello che non si fissa nei tessuti, nel fegato viene trasformato in androsterone ed eziocolanolone, che vengono escreti nella bile o nell’urina dopo essere stati glucurono – o solfo- coniugati

63
Meccanismo d’azione Il recettore nucleare è una proteina codificata da un gene del cromosoma X Al contatto con l’ormone (testosterone o DHT), il recettore dimerizza e si lega ai geni bersaglio attivando la trascrizione Sintesi proteica (ormone anabolizzante) Il testosterone viene trasformato dall’enzima aromatasi in estradiolo che si lega al recettore per gli estrogeni determinando effetti importanti sulla maturazione dell’osso. L’assenza di aromatasi e la mancata formazione di estradiolo può comportare una severa osteopenia e aspetto eunucoide
, il recettore dimerizza e si lega ai geni bersaglio attivando la trascrizione. Sintesi proteica (ormone anabolizzante) Il testosterone viene trasformato dall’enzima aromatasi in estradiolo che si lega al recettore per gli estrogeni determinando effetti importanti sulla maturazione dell’osso. L’assenza di aromatasi e la mancata formazione di estradiolo può comportare una severa osteopenia e aspetto eunucoide.")
64
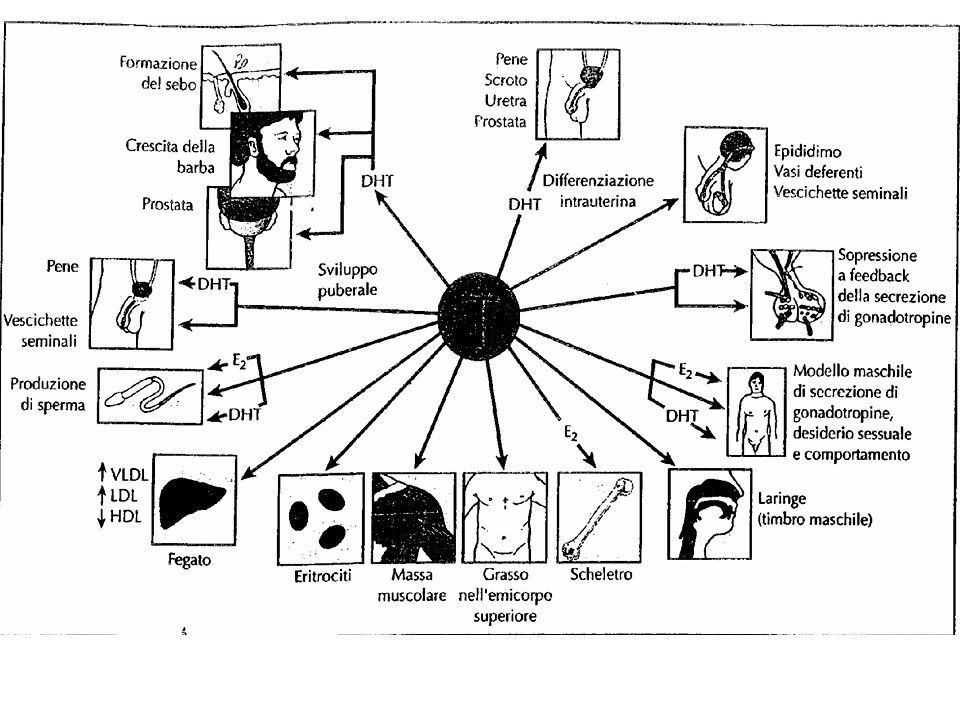
Effetti biologici Effetti durante l’embriogenesi
Gli effetti indotti sono diversi a seconda degli stadi di maturazione sessuale della vita dell’uomo Effetti durante l’embriogenesi E’ importante per lo sviluppo dei caratteri sessuali primari (genitali interni ed esterni)
")
65
Effetti alla nascita Aumento della secrezione che ha effetto permissivo (priming) sul tratto urogenitale maschile, facilitando l’ingrandimento del pene
 sul tratto urogenitale maschile, facilitando l’ingrandimento del pene.")
66
Effetti alla pubertà E’ responsabile della comparsa dei caratteri sessuali secondari Induce l’accrescimento dell’osso e la ritenzione di Ca Aumenta il metabolismo basale del 5-10% Determina un cambiamento comportamentale, anche sessuale, stimolando la libido, dando inizio alla spermatogenesi e determinando la capacità di fecondare Aumenta l’aggressività Stimola la eritropoiesi e induce l’aumento dell’ematocrito Riduce le HDL che trasportano il colesterolo

68
Effetti nell’età adulta
Mantenimento: delle secrezioni prostatiche e delle vescicole seminali della densità ossea, della massa muscolare, della distribuzione del pelo della libido, della potentia generandi e della potentia coeundi dell’azione sull’eritropoiesi e sulle HDL

70
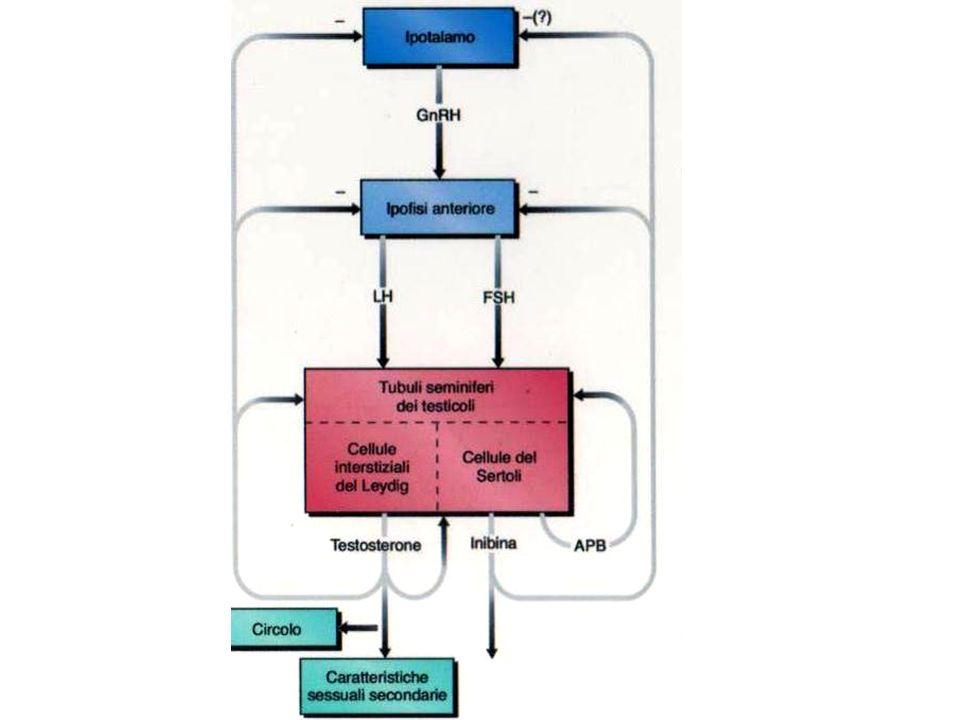
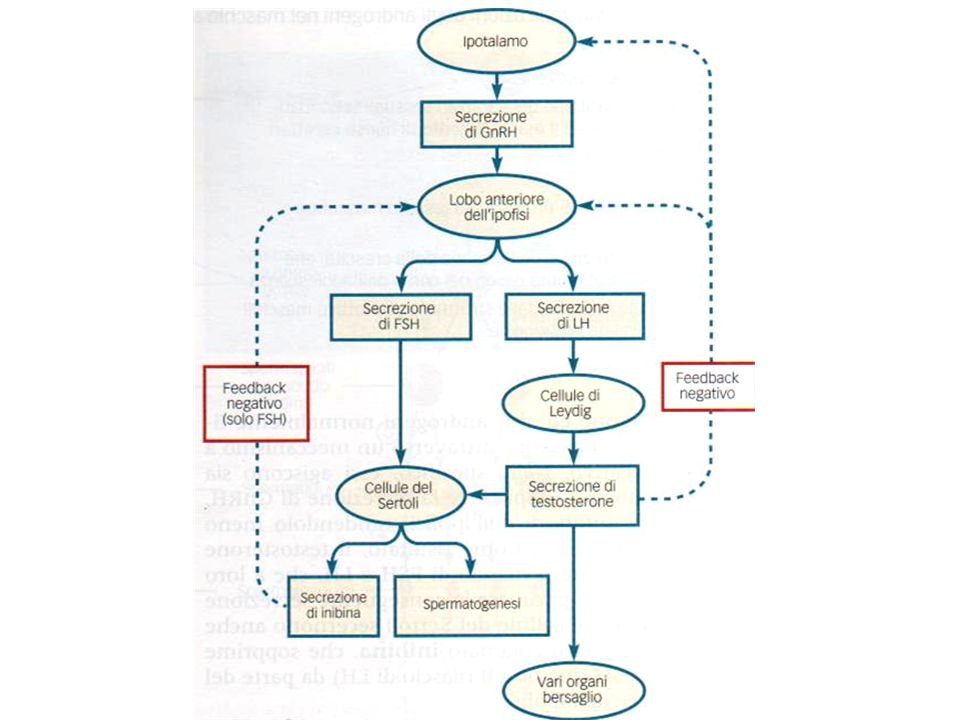
Controllo della funzione testicolare
L’ipotalamo rilascia l’ormone liberante le gonadotropine (GnRH) che stimola l’ipofisi L’adenoipofisi secerne l’FSH e l’LH L’FSH ha azione spermatogenetica insieme al testosterone e stimola la secrezione di inibina che agisce a feedback negativo sull’FSH L’LH (o ICSH interstitial cells stimulating hormon) stimola la secrezione di testosterone dalle cell di Leydig. Il testosterone agisce con feedback negativo sulla secrezione di LH
 che stimola l’ipofisi. L’adenoipofisi secerne l’FSH e l’LH. L’FSH ha azione spermatogenetica insieme al testosterone e stimola la secrezione di inibina che agisce a feedback negativo sull’FSH. L’LH (o ICSH interstitial cells stimulating hormon) stimola la secrezione di testosterone dalle cell di Leydig. Il testosterone agisce con feedback negativo sulla secrezione di LH.")
72
Apparato riproduttivo femminile
Il sistema riproduttivo maschile ha solo il compito di produrre lo sperma e depositarlo nel sistema riproduttivo femminile Il sistema riproduttivo femminile è responsabile: Della produzione delle cellule uovo Del trasporto delle cellule uovo Di tutto ciò che occorre per la nascita di un nuovo essere umano

73
Caratteristiche del sistema riproduttivo femminile
Cambiamenti ciclici di attività: ogni 28 giorni cambiamenti strutturali e funzionali (ciclo mestruale) sono accompagnati da cambiamenti della secrezione di ormoni ipotalamici, ipofisari e ovarici ed iniziano con la mestruazione, perdita di sangue e tessuto dalla superficie della parete uterina Periodo di fertilità ristretto: a metà del ciclo mestruale viene rilasciato un ovulo (ovulazione), prerequisito per la fecondazione. La donna è fertile solo durante i giorni del ciclo che coincidono all’incirca con l’ovulazione Produzione limitata di gameti: il numero delle cellule germinali è fisso alla nascita (ca 2 milioni), ma il numero di ovuli potenziali diminuisce nel corso della vita perché la maggior parte delle cellule germinali degenera (atresia). Solo circa 400 cellule uovo vengono rilasciate nel corso della vita di una donna
 sono accompagnati da cambiamenti della secrezione di ormoni ipotalamici, ipofisari e ovarici ed iniziano con la mestruazione, perdita di sangue e tessuto dalla superficie della parete uterina. Periodo di fertilità ristretto: a metà del ciclo mestruale viene rilasciato un ovulo (ovulazione), prerequisito per la fecondazione. La donna è fertile solo durante i giorni del ciclo che coincidono all’incirca con l’ovulazione. Produzione limitata di gameti: il numero delle cellule germinali è fisso alla nascita (ca 2 milioni), ma il numero di ovuli potenziali diminuisce nel corso della vita perché la maggior parte delle cellule germinali degenera (atresia). Solo circa 400 cellule uovo vengono rilasciate nel corso della vita di una donna.")
74
Sistema riproduttivo femminile
Utero: corpo e cervice che immette nel canale cervicale, diretto alla vagina Tube uterine o di Falloppio: terminano con l’infundibulo (imbutiforme) frangiato da fimbrie che racchiudono parzialmente l’ovaio del rispettivo lato
 frangiato da fimbrie che racchiudono parzialmente l’ovaio del rispettivo lato.")
75
La parete uterina Strato esterno di cell epiteliali e tessuto connettivo, perimetrio Strato intermedio, spesso, di muscolatura liscia, miometrio Strato interno di cell epiteliali e connettivo, endometrio, che contiene ghiandole secernenti un fluido che begne la parete uterina

76
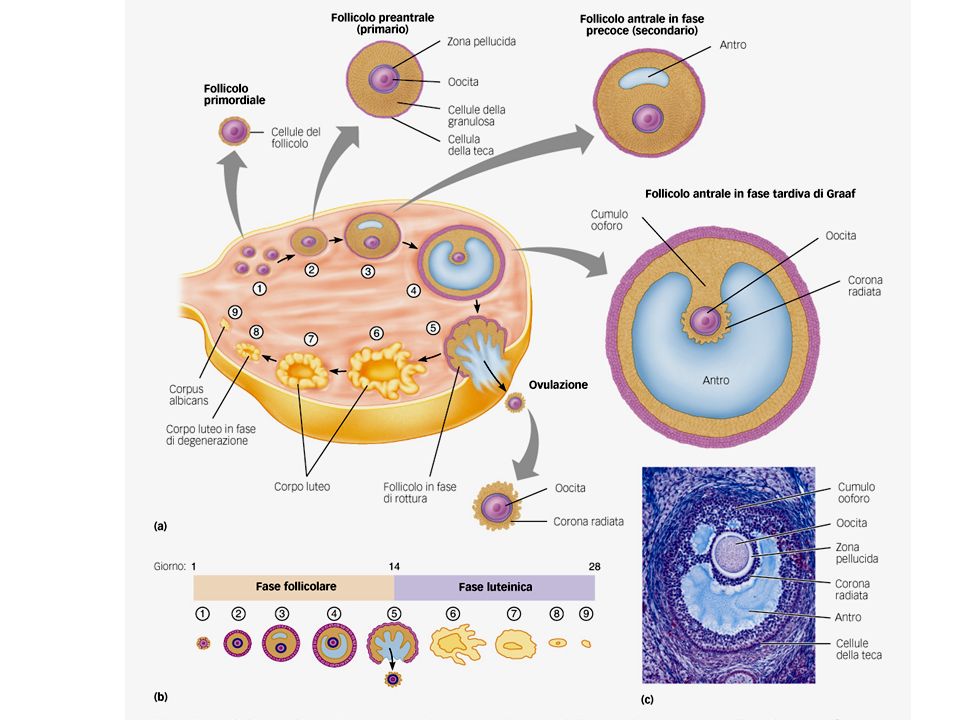
L’ovaio Struttura ovoidale di 2-4 cm costituita da tessuto connettivo (stroma). Si distinguono: una parte corticale spessa che contiene i follicoli ovarici in vari stati di sviluppo una parte midollare centrale con vasi sanguigni e nervi La gametogenesi avviene nell’ovaio

77
I genitali esterni Costituiti dalla vulva che comprende:
Grandi e piccole labbra, che circondano il meato vaginale Vestibolo, che conduce a vagina e uretra Clitoride, piccolo organo erettile che si trova di fronte al vestibolo Ghiandole vestibolari, che secernono fluido lubrificante per facilitare la penetrazione del pene nella vagina Imene, sottile membrana contenente piccoli vasi sanguigni che nelle donne vergini occlude parzialmente il meato vaginale

78
Lo sviluppo dei caratteri sessuali primari della donna è controllato dagli estrogeni
Gli estrogeni controllano anche i più evidenti caratteri sessuali secondari femminili : lo sviluppo delle mammelle, la distribuzione del tessuto adiposo su fianchi e cosce Altre caratteristiche sessuali secondarie femminili sono regolate dagli androgeni prodotti dalla corteccia surrenale : la crescita dei peli pubici e ascellari e anche la libido

79
La gametogenesi femminile
Entro il sesto mese di sviluppo embrionale: le cellule germinali, oogoni, completano la divisione mitotica e la profase della meiosi. La mitosi delle cellule germinali termina e non possono essere prodotti altri oociti (ca 7 milioni) Alla nascita: l’ovaio contiene circa 2 milioni di oociti primari Alla pubertà: gli oociti saranno ridotti a circa Nel periodo fertile: circa oociti maturano e vengono espulsi o fecondati Al climaterio: solo pochi follicoli primitivi saranno rimasti nelle ovaie, la riproduzione ha termine (menopausa)
 Alla nascita: l’ovaio contiene circa 2 milioni di oociti primari. Alla pubertà: gli oociti saranno ridotti a circa Nel periodo fertile: circa oociti maturano e vengono espulsi o fecondati. Al climaterio: solo pochi follicoli primitivi saranno rimasti nelle ovaie, la riproduzione ha termine (menopausa)")
80
Periodo fertile Durante il periodo fertile, ogni mese, l’ovaio rilascia uno o due cellule uovo (ovulazione). Con la I divisione meiotica, ogni oocita primario si divide in due cellule, un grande ovulo o oocita secondario e un piccolo primo corpo polare, entrambi con corredo diploide. Il primo corpo polare degenera. Con la II divisione meiotica, se l’ovulo non viene fecondato, i cromatidi fratelli si separano ma restano nella stessa cellula e l’ovulo si disintegra o viene espulso; se l’ovulo viene fecondato, la meiosi si completa, i cromatidi fratelli vanno a cellule diverse, uno zigote e un secondo corpo polare che degenera. Ogni oocita primario produce quindi un solo ovulo Prima della nascita

81
Cambiamenti ciclici nella donna
Il processo ciclico mensile che porta all’ovulazione (ciclo mestruale) implica numerosi cambiamenti: Cambiamenti della struttura e della funzione delle ovaie (ciclo ovarico) Cambiamenti della struttura e della funzione dell’utero (ciclo uterino) Cambiamenti ciclici della secrezione degli ormoni ovarici, ipotalamici e ipofisari
 implica numerosi cambiamenti: Cambiamenti della struttura e della funzione delle ovaie (ciclo ovarico) Cambiamenti della struttura e della funzione dell’utero (ciclo uterino) Cambiamenti ciclici della secrezione degli ormoni ovarici, ipotalamici e ipofisari.")
82
Il ciclo ovarico E’ diviso in tre fasi
Fase follicolare: periodo di crescita dei follicoli nell’ovaio Ovulazione: quando uno o più follicoli sono giunti a maturazione, e l’ovaio li rilascia Fase luteale o luteinica: fase postovulatoria, in cui il follicolo si trasforma in corpo luteo, così chiamato per i suoi depositi gialli di pigmento e lipidi. Secerne ormoni per preparare alla gravidanza, ma se questa non si instaura, il corpo luteo cessa di funzionare dopo due settimane ed il ciclo ovarico ricomincia

83
Il ciclo uterino E’ regolato da ormoni ovarici. Si suddivide in:
Mestruo: sanguinamento uterino corrispondente alla fase follicolare Fase proliferativa: l’endometrio si ispessisce in preparazione di una eventuale gravidanza in corrispondenza dell’ultima parte della fase follicolare Fase secretoria: corrisponde alla fase luteale, quando gli ormoni prodotti dal corpo luteo convertono l’endometrio ispessito in una struttura secernente. Se non si ha gravidanza, gli strati superficiali dell’endometrio vengono persi durante la mestruazione

84
I cicli ormonali I cicli ovarico ed uterino sono sotto il controllo di diversi ormoni: GnRH rilasciato dall’ipotalamo FSH ed LH rilasciati dall’adenoipofisi Estrogeni, progesterone ed inibine prodotti dall’ovaio Durante la fase follicolare prevalgono gli estrogeni, durante la fase luteinica prevale il progesterone

85
Il ciclo ovarico: la fase follicolare
Nell’utero ha inizio la mestruazione. Nell’ovaio, alcuni dei follicoli primitivi formati da un oocita (cell. uovo) circondato da cellule follicolari epiteliali specializzate, iniziano a svilupparsi sotto l’influenza dell’FSH. Si formano i follicoli preantrali o primari in cui le cellule epiteliali proliferano formando più strati (cellule della granulosa) e lo strato più esterno formerà le cellule della teca (LH). Le cell della granulosa secernono una membrana (zona pellucida) -che isola l’oocita dagli altri tessuti- e formano delle estensioni citoplasmatiche che raggiungono l’oocita per assicurarne il nutrimento. La teca produce androgeni che diffondono verso le cell della granulosa che li convertono in estrogeni. Gli estrogeni esercitano un feedback negativo su FSH ed LH, impedendo lo sviluppo di altri follicoli nello stesso ciclo, e positivo sulle cell della granulosa che secernono altri estrogeni
 circondato da cellule follicolari epiteliali specializzate, iniziano a svilupparsi sotto l’influenza dell’FSH. Si formano i follicoli preantrali o primari in cui le cellule epiteliali proliferano formando più strati (cellule della granulosa) e lo strato più esterno formerà le cellule della teca (LH). Le cell della granulosa secernono una membrana (zona pellucida) -che isola l’oocita dagli altri tessuti- e formano delle estensioni citoplasmatiche che raggiungono l’oocita per assicurarne il nutrimento. La teca produce androgeni che diffondono verso le cell della granulosa che li convertono in estrogeni. Gli estrogeni esercitano un feedback negativo su FSH ed LH, impedendo lo sviluppo di altri follicoli nello stesso ciclo, e positivo sulle cell della granulosa che secernono altri estrogeni.")
86
Controllo ormonale

87
Il ciclo ovarico: la fase follicolare
I follicoli che continuano a svilupparsi formano una cavità piena di fluido (antro), che, con lo sviluppo, aumenta di dimensioni: fase antrale precoce Dopo circa 7 giorni solo uno di questi follicoli completa il suo sviluppo (follicolo dominante) mentre gli altri vanno in atresia Nell’utero termina la mestruazione e l’endometrio inizia a crescere sotto la spinta degli estrogeni follicolari Gli estrogeni inducono anche le ghiandole mucose della cervice uterina a produrre muco chiaro e acquoso
, che, con lo sviluppo, aumenta di dimensioni: fase antrale precoce. Dopo circa 7 giorni solo uno di questi follicoli completa il suo sviluppo (follicolo dominante) mentre gli altri vanno in atresia. Nell’utero termina la mestruazione e l’endometrio inizia a crescere sotto la spinta degli estrogeni follicolari. Gli estrogeni inducono anche le ghiandole mucose della cervice uterina a produrre muco chiaro e acquoso.")
88
Il ciclo ovarico: la fase follicolare
Nel follicolo dominante l’antro cresce e le cellule della granulosa formano la corona radiata e il cumulo ooforo (follicolo di Graaf) Si verifica la meiosi, l’oocita secondario si stacca dalla parete con la corona radiata e fluttua nell’antro. Il follicolo raggiunge i 2-2,5 cm prima dell’ovulazione
 Si verifica la meiosi, l’oocita secondario si stacca dalla parete con la corona radiata e fluttua nell’antro. Il follicolo raggiunge i 2-2,5 cm prima dell’ovulazione.")
89
Il ciclo ovarico: la fase follicolare
Quando la fase follicolare si avvicina al termine, la secrezione ovarica degli estrogeni raggiunge il picco Quando la fase follicolare termina, le cell della granulosa secernono inibina e progesterone, oltre agli estrogeni che ora vanno non ad inibire, bensì a stimolare la risposta dell’ipofisi alle GnRH. Di conseguenza si osserva il picco di secrezione dell’LH (essenziale per l’ovulazione). L’FSH raggiunge pure il suo picco, ma più basso perché viene soppresso dall’inibina e dagli estrogeni L’endometrio continua a crescere (3-4 mm) e la ghiandole cervicali secernono muco viscoso
. L’FSH raggiunge pure il suo picco, ma più basso perché viene soppresso dall’inibina e dagli estrogeni. L’endometrio continua a crescere (3-4 mm) e la ghiandole cervicali secernono muco viscoso.")
90
Controllo ormonale

91
Funzioni delle cellule della granulosa
Per molti versi, le funzioni delle cell della granulosa sono simili a quelle delle cell del Sertoli: Provvedono al nutrimento per lo sviluppo degli oociti Secernono sostanze paracrine che stimolano lo sviluppo del follicolo (sotto influenza di estrogeni ed FSH) Secernono inibina per l’inibizione della secrezione di FSH Secernono estrogeni che derivano dagli androgeni sintetizzati dalle cell della teca Secernono progesterone Secernono liquido antrale Formano una barriera per prevenire l’ingresso di sostanze che potrebbero danneggiare la cellula in via di sviluppo
 Secernono inibina per l’inibizione della secrezione di FSH. Secernono estrogeni che derivano dagli androgeni sintetizzati dalle cell della teca. Secernono progesterone. Secernono liquido antrale. Formano una barriera per prevenire l’ingresso di sostanze che potrebbero danneggiare la cellula in via di sviluppo.")
92
Il ciclo ovarico: l’ovulazione
Dopo h dal picco dell’LH si ha l’ovulazione Il follicolo maturo secerne collagenasi che degrada il collagene nel tessuto connettivo che tiene unite le cell follicolari. I prodotti della rottura del collagene determinano una risposta infiammatoria, richiamando leucociti secernenti prostaglandine all’interno del follicolo. Il follicolo si rompe, il fluido antrale trasporta l’uovo alla superficie dell’ovaio L’ovulo cade nelle tube di Falloppio ed è trasportato verso l’utero dove viene fecendato oppure degenera

93
Il ciclo ovarico: la fase luteale
Dopo la caduta dell’ovulo, il picco dell’LH determina la migrazione delle cell follicolari nello spazio antrale dove si mescolano a quelle della granulosa riempiendo la cavità. Entrambi i tipi di cell si trasformano in cell luteali (luteinizzazione) Le cell luteali accumulano gocce lipidiche e granuli di glicogeno nel citoplasma ed iniziano a secernere progesterone mentre la sintesi degli estrogeni diminuisce
 Le cell luteali accumulano gocce lipidiche e granuli di glicogeno nel citoplasma ed iniziano a secernere progesterone mentre la sintesi degli estrogeni diminuisce.")
95
Il ciclo ovarico: la fase luteale
Il corpo luteo secerne progesterone (ormone dominante in questa fase) ed estrogeni: entrambi, insieme all’inibina, esercitano un feedback negativo sulla secrezione di gonadotropine Il progesterone stimola l’endometrio: si formano nuovi vasi sanguigni, le ghiandole endometriali si avvolgono a spirale, nelle cell endometriali si accumulano lipidi e glicogeno che serviranno a nutrire l’embrione mentre si sta formando la placenta Il progesterone induce ispessimento del muco cervicale che crea un tappo che impedisce l’ingresso di batteri nell’utero Il progesterone ha azione termogenica: la temperatura corporea si alza di 0,3-0,5 gradi durante la fase luteale e resta elevata fino al mestruo. Questa variazione di temperatura permette di valutare se il ciclo è stato ovulatorio o anovulatorio
 ed estrogeni: entrambi, insieme all’inibina, esercitano un feedback negativo sulla secrezione di gonadotropine. Il progesterone stimola l’endometrio: si formano nuovi vasi sanguigni, le ghiandole endometriali si avvolgono a spirale, nelle cell endometriali si accumulano lipidi e glicogeno che serviranno a nutrire l’embrione mentre si sta formando la placenta. Il progesterone induce ispessimento del muco cervicale che crea un tappo che impedisce l’ingresso di batteri nell’utero. Il progesterone ha azione termogenica: la temperatura corporea si alza di 0,3-0,5 gradi durante la fase luteale e resta elevata fino al mestruo. Questa variazione di temperatura permette di valutare se il ciclo è stato ovulatorio o anovulatorio.")
96
Controllo ormonale

97
Il ciclo ovarico: la fase luteale
Il corpo luteo sopravvive circa 12 gg, poi, se non inizia una gravidanza, va incontro ad apoptosi e diventa inattivo (corpo albicans) La secrezione di progesterone ed estrogeni diminuisce e la secrezione di FSH ed LH aumenta Con la diminuzione del progesterone, i vasi sanguigni dell’endometrio si contraggono e le cel superficiali degenerano, si desquamano e ha inizio la mestruazione Il flusso mestruale non presenta coaguli perché contiene plasmina che produce la lisi dei coaguli
 La secrezione di progesterone ed estrogeni diminuisce e la secrezione di FSH ed LH aumenta. Con la diminuzione del progesterone, i vasi sanguigni dell’endometrio si contraggono e le cel superficiali degenerano, si desquamano e ha inizio la mestruazione. Il flusso mestruale non presenta coaguli perché contiene plasmina che produce la lisi dei coaguli.")
98
Controllo ormonale

99
Il ciclo completo

100
Ormoni ovarici L’ovaio produce tre tipi di steroidi:
estrogeni, prodotti ciclicamente dalle cell follicolari progesterone, sintetizzato dal corpo luteo androgeni, prodotti in piccola quantità dalla zona midollare

101
Estrogeni Derivano dall’estrano: Estrone, Estriolo, !7β-estradiolo, il più attivo Il 17β-estradiolo circola in forma libera (20%) o legata ad albumina o a una proteina specifica Si lega a un recettore nucleare e avviene la dimerizzazione prima che venga stimolata l’attività dei fattori di trascrizione Effetti biologici: ipertrofia e iperplasia delle cellule e delle ghiandole endometriali e aumento della secrezione cervicale; sviluppo dei dotti galattofori; sviluppo delle vie genitali e dei caratteri sessuali femminili secondari; effetti sul metabolismo proteico (stimolando l’incremento dello sviluppo somatico); sono responsabili dell’arresto della crescita (facilitano la saldatura della cartilagine di coniugazione); aumentano la secrezione di angiotensinogeno e di molecole vettrici di ormoni, come la TBG, sostengono la libido
 o legata ad albumina o a una proteina specifica. Si lega a un recettore nucleare e avviene la dimerizzazione prima che venga stimolata l’attività dei fattori di trascrizione. Effetti biologici: ipertrofia e iperplasia delle cellule e delle ghiandole endometriali e aumento della secrezione cervicale; sviluppo dei dotti galattofori; sviluppo delle vie genitali e dei caratteri sessuali femminili secondari; effetti sul metabolismo proteico (stimolando l’incremento dello sviluppo somatico); sono responsabili dell’arresto della crescita (facilitano la saldatura della cartilagine di coniugazione); aumentano la secrezione di angiotensinogeno e di molecole vettrici di ormoni, come la TBG, sostengono la libido.")
102
Progesterone Deriva dal pregnano
Circola legato a proteine specifiche o all’albumina Ha recettori nucleari e la dimerizzazione consente il legame all’elemento di risposta ai geni bersaglio Effetti biologici: ipertrofia della mucosa uterina (per prepararla all’annidamento della cellula uovo) e mantenimento della gravidanza; iperplasia ed aumento dell’attività secretoria dell’endometrio in caso di fecondazione; inibizione delle contrazioni miometrali; proliferazione degli alveoli ghiandolari della mammella (come gli estrogeni); effetti termogenici (aumento della temperatura corporea durante l’ovulazione); riduce il riassorbimento renale di Na (e quindi la ritenzione idrica) con effetto opposto a quello degli estrogeni; diminuisce la sintesi del recettore degli estrogeni e facilita la loro inattivazione
 e mantenimento della gravidanza; iperplasia ed aumento dell’attività secretoria dell’endometrio in caso di fecondazione; inibizione delle contrazioni miometrali; proliferazione degli alveoli ghiandolari della mammella (come gli estrogeni); effetti termogenici (aumento della temperatura corporea durante l’ovulazione); riduce il riassorbimento renale di Na (e quindi la ritenzione idrica) con effetto opposto a quello degli estrogeni; diminuisce la sintesi del recettore degli estrogeni e facilita la loro inattivazione.")
103
Androgeni ovarici Derivano dall’androstano: androstenedione che viene convertito in testosterone nel fegato e nel tessuto adiposo, e che, a sua volta, viene convertito in diidrotestosterone (più attivo) nei tessuti bersaglio (follicoli piliferi, ghiandole sebacee, genitali esterni) Nelle femmine gli androgeni sono prodotti collaterali della secrezione dell’estradiolo (ovaio) e del cortisolo (surrene) pertanto non riconoscono uno specifico meccanismo ormonale di regolazione e non si può definire un meccanismo di controllo a feedback da parte dell’ipofisi Circola legato ad albumine, o a proteine specifiche Recettori nucleari che dimerizzano per stimolare i fattori di trascrizione
 nei tessuti bersaglio (follicoli piliferi, ghiandole sebacee, genitali esterni) Nelle femmine gli androgeni sono prodotti collaterali della secrezione dell’estradiolo (ovaio) e del cortisolo (surrene) pertanto non riconoscono uno specifico meccanismo ormonale di regolazione e non si può definire un meccanismo di controllo a feedback da parte dell’ipofisi. Circola legato ad albumine, o a proteine specifiche. Recettori nucleari che dimerizzano per stimolare i fattori di trascrizione.")
104
Controllo dell’attività secernente dell’ovaio
Le cell della teca interna forniscono androgeni alle cell della granulosa che producono estrogeni circolanti, e inibiscono la secrezione di GnRH, FSH, ed LH. L’inibina delle cell della granulosa inibisce la secrezione di FSH. L’LH regola le cell della teca, mentre le cell della granulosa sono regolate sia da LH sia da FSH

Presentazioni simili























 PER CONTROLLARE L’ATTIVITA’ DI ALTRI ORGANI AD UNA CERTA DISTANZA L’ORMONE: PRODOTTO.>")
