Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
L’esportazione delle proteine verso la membrana esterna

2
Esperimento della fusione malE-LacZ
LacZ è una proteina citoplasmatica lacZ lacZ Esperimento della fusione malE-LacZ malE lacZ IM malE lacZ malE lacZ malE lacZ malE lacZ malE lacZ malE lacZ lacZ TA malE lacZ M In presenza di maltosio si ha una produzione elevata della fusione malE-lacZ che porta a morte la cellula . Si possono quindi selezionare i Mal resistenti

3
Gli attori del sistema Sec
SecB è una chaperonina, ovvero interagisce con la sequenza segnale della proteina nella fase di traduzione mantenendola in uno stato strutturale non corretto. SecA è una proteina con attività ATPasica e che è in grado di interagire con SecB quando questa ha catturato una proteina SecYEG formano un complesso trans-membranario che permette l’esportazione della proteina attraverso la membrana esterna (YEG sono proteina transmembrana SecDFYaiC coinvolte nel rilascio della proteina esportato e nella regolazione delle interazioni di SecA con la membrana (D e F sono proteina della membrana interna ma rivolte verso lo spazio periplasmatico)
")
4
I componenti del sistema SEC
Proteine citoplasmatiche * Sec A * SecB Proteine della membrana interna: * SecYEG formano il canale attraverso il quale passerà la proteina da esportare * SecDFYajC si associano a SecYEG e sono coinvolte nel rilascio delle proteine esportate dalla membrana e nel riciclo di SecA

5
Il sistema SEC : General Export Pathway (GEP )
")
6
La proteina SecA è un ATP binding protein ( dotata di un motivo ABC) che fornisce l’energia per il passaggio attraverso la membrana consumando ATP
 che fornisce l’energia per il passaggio attraverso la membrana consumando ATP")
7
SecB è una ciaperonina citoplasmatica che entra in contatto con la proteina da esportare in modo che questa non assuma una conformazione inadatta all’esportazione. E’ cruciale che la sequenza segnale rimanga esposta in modo che possa interagire con il complesso di esportazione

8
La proteina SecB interagisce con il segnale peptide della proteina da esportare durante la sua traduzione. La trasporta al complesso SecA+SecYEG e l’affida a SecA. SecA, attraverso il consumo di ATP inserisce e spinge la proteina attraverso il complesso SecYEG. Appena superato questo complesso una peptidasi (Lep) proteolizza il segnale dal resto del peptide attivando la proteina
 proteolizza il segnale dal resto del peptide attivando la proteina.")
9
Dal citoplasma allo spazio periplasmatico: il sistema Sec
SecYEG

10
Modello per l’attraversamento Sec dipendente delle proteine
D=ADP T=ATP 4. La perdita di ADP da parte di SecA favorisce l’entrata di SecA nella membrana 8. L’entrata della sequenza segnale nel canale di SecY apre il tappo lasciando il canale disponibile alla fuoriuscita della proteina 4-11. Questo ciclo deve essere ripetute perché ogni volta passano circa 30 AA della preproteina

11
Il sistema SRP (Signal Recognition Particle )
Nel caso del sistema SRP il processo è molto simile ma cambiano gli “attori”. In questo caso non abbiamo più le proteine SecB ma una piccola nucleoproteina (SRP) composta da una proteina (codificata dal gene ffh) e un piccolo frammento di RNA (4.5S). Questo RNA non appartiene a nessuna delle classiche classi di RNA (tRNA, mRNA o rRNA) Questo complesso sostituisce funzionalmente (pur non agendo da chaperonina) la proteina SecB.
 composta da una proteina (codificata dal gene ffh) e un piccolo frammento di RNA (4.5S). Questo RNA non appartiene a nessuna delle classiche classi di RNA (tRNA, mRNA o rRNA) Questo complesso sostituisce funzionalmente (pur non agendo da chaperonina) la proteina SecB.")
12
Ruolo di SRP Il meccanismo ricalca sostanzialmente quello visto per il sistema SecA/B

13
E.coli 114 nucleotides RNA

14
Il complesso SRP+preproteina viene diretto verso la membrana interna dove interagisce con la proteina di membrana FtsY. Questo nuovo complesso costituito d FtsY/SRP/preproteina interagisce con SecYEG e permette l’esportazione della proteina attraverso il complesso trans-membranario.

15
Dal citoplasma verso la membrana interna: strategie diverse verso il sistema Sec
3’

17
I componenti del sistema Sec di E.coli

18
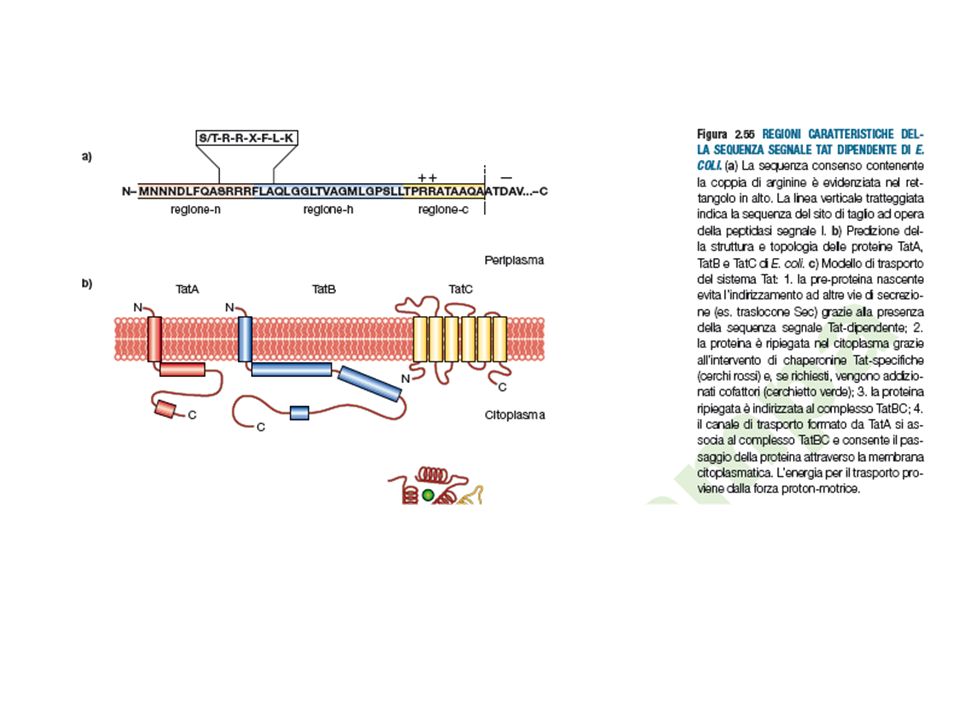
Twin Arginine Traslocase
Il Sistema TAT Twin Arginine Traslocase Le proteine trasportate dal sistema Sec sono ripiegate dopo aver raggiunto lo spazio periplasmatico. Vi sono proteine che contengono nella loro struttura cofattori e sono correttamente assemblate nel citoplasma. Quindi vengono trasferite assieme al cofattore da un sistema diverso da Sec. Proteine ferro-zolfo proteine del sistema redox Proteine coinvolte nella biosintesi della OM (membrane esterna) La proteine sono dotate di una breve sequenza segnale contenente una coppia di residui d’arginina. La sequenza segnale sulla proteina è riconosciuta dalla proteina TatBC che conducono la proteina al trasportare di membrana TatA. L’energia viene fornita dalla forza proton motrice.
 La proteine sono dotate di una breve sequenza segnale contenente una coppia di residui d’arginina. La sequenza segnale sulla proteina è riconosciuta dalla proteina TatBC che conducono la proteina al trasportare di membrana TatA. L’energia viene fornita dalla forza proton motrice.")
19
Caratteristiche della sequenza segnale delle proteine esportate
dal sistema Sec dal sistema TAT Motivo per l’esportazione TAT= S-R-R-x-F-L-K (ser-arg-arg-x-fenil-leu-lis)
")
20
FIG 2 - Il ciclo inizia quando la sequenza segnale contenente le 2 arginine si lega al complesso di membrana TatBC. Il motivo RR è riconosciuto da TatC .Sfruttando la forza proton motrice TatA si dirige verso TatBC quando la preproteina è gia legata a TatBC. La proteina a questo punto viene traslocata attraverso il poro formato da TatA. Alla fine TatBC si dissociano da TatA.

23
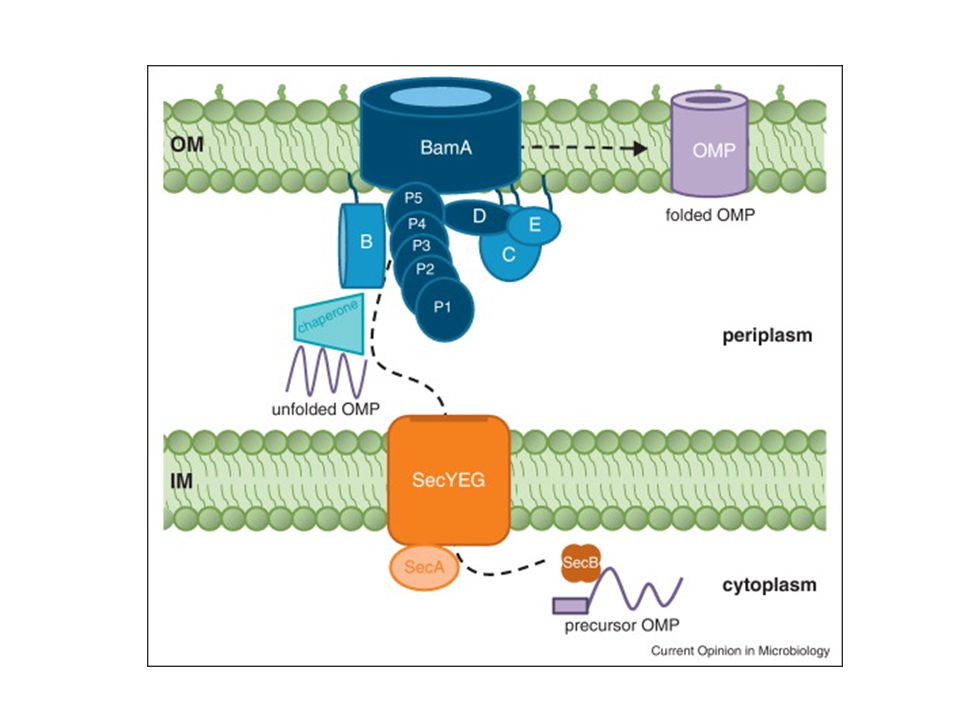
Come arrivano le proteine della OM ( OMPs) alla loro destinazione finale?
Tutte le OMPs contengono un numero pari di foglietti b che si ripiegano in una struttura a b barrel con i residui idrofobici rivolti verso l’esterno. Nello spazio periplasmatico si trovano: Skp ( 17 kb protein) : proteina che si lega alle OMPs non appena emergono dal canale SEC e prevengono il ripiegamento incorretto e l’aggregazione SurA proteina che contribuisce al corretto ripiegamento delle OMPs Dbs AC che induce la formazione dei ponti disolfuro nelle OMPs prima che siano inserite nella OM
 : proteina che si lega alle OMPs non appena emergono dal canale SEC e prevengono il ripiegamento incorretto e l’aggregazione. SurA proteina che contribuisce al corretto ripiegamento delle OMPs. Dbs AC che induce la formazione dei ponti disolfuro nelle OMPs prima che siano inserite nella OM.")
24
Arrivo delle OMP nella OM: ruolo di BamA
SurA contribuisce al corretto ripiegamento Skp previene il ripiegamento incorretto delle proteine che escono dal canale Sec

25
BamA (Omp85) BamA è una proteina conservata in tutti i batteri Gram- e nei mitocondri (Tob55). Identificata in Neisseria con il nome di OMP85 BamA è costituita da 12 foglietti b disposti a b -barrel con una lunga estremità N terminale nello spazio periplasmatico (dominio POTRA). Il domino N terminale di BamA riconosce sequenze specifiche caratteristiche dell’estremità C-terminale di molte OMP. Phe(o Trp) al C poi residui idrofobici in posizione dal Cter.
. Il domino N terminale di BamA riconosce sequenze specifiche caratteristiche dell’estremità C-terminale di molte OMP. Phe(o Trp) al C poi residui idrofobici in posizione dal Cter.")
26
In E. coli e stato identificato e parzialmente caratterizzato un complesso multiproteico (Bam, β-barrel assembly machine) che comprende una proteina integrale di membrana (BamA) e quattro lipoproteine, e che costituisce la “macchina proteica” preposta a questo processo.
 che comprende una proteina integrale di membrana (BamA) e quattro lipoproteine, e che costituisce la macchina proteica preposta a questo processo..")
28
Questo dominio N-POTRA è coinvolto nel legame con la OMP mantenendola ancora accessibile ad altre ciaperonine che contribuiscono al corretto ripiegamento. La OMP successivamente si inserisce nel canale interno di BamA. Il complesso BamA-OMP si apre lateralmente per permettere il corretto inserimento della OMP nella M.E.

29
Polypeptide translocation associated motifs PROTRA
Omp85 =BamA

30
Alcune proteine richiedono l’assistenza di altre proteine definiti Ciaperoni per ripiegarsi in modo corretto. In E.coli vi sono 4 ciaperoni molto importanti DnaK, DnaJ GroEL and GroES

31
DnaK e DNAJ sono enzimi ATP-dipendenti che legano i polipeptidi neosintetizzati rallentandone il ripiegamento in modo da favorirne uno esatto. Se il complesso DnaKJ è incapace di far assumere alla proteina il corretto folding può trasferire la proteina parzialmente ripiegata ai due complessi proteici GroEL-GroES. La proteina entra dentro GroEL, una grande proteina a forma di barile che usando energia dall’idrolisi di ATP e coadiuvata dal complesso GroES, ripiega correttamente la proteina.

33
Le PORINE A differenza della MI la ME dei Gram– è parzialmente permeabile alle piccole molecole Grazie alla presenza di proteine della membrana esterna PORINE che svolgono la funzione di canali per entrata/uscita di sostanze idrofiliche a basso PM PORINE aspecifiche formano canali acquosi per il passaggio di piccole molecole PORINE specifiche per il passaggio di uno o un piccolo gruppo di composti

34
Le proteine della membrana esterna
Le porine specifiche o aspecifiche Proteine di membrana lipoproteine

35
La lipoproteina Sul lato interno della M.E. è presente un complesso lipoproteico costituito dalla LIPOPROTEINA. Questa proteina è una piccola proteina che svolge funzioni di ancoraggio tra M.E. e peptidoglicano

36
Le lipoproteine partecipano a numerose funzioni cellulari stabilizzazione delle strutture di superficie, resistenza ad antibiotici trasduzione del segnale. Sono noti circa 90 i tipi di lipoproteine diverse nella maggioranza associate all OM ( in E.coli tutte sul lato interno in altri batteri anche sul lato esterno). La lipoproteina di Braun è la più abbondante è una piccola proteina di 58 AA Con N terminale si ancora alla OM grazie alla modificazione lipidica della Cisteina al +1 con la lisina al C terminale forma un legame peptidico con il DAP

37
LIPOPROTEINE Le lipoproteine sono ancorate alla OM tramite l’estremità N-terminale della N-acyl diacylglyceril cisteina ( cisteina lipidata). Il processo di lipidazione e corretto ripiegamento ha luogo dopo che la lipoproteina ha attraversato la IM. Per attraversare lo spazio periplasmatico la lipoproteina ha bisogno del complesso LolCDE localizzato nella MI, che contiene un ATP binding cassette. L’energia derivante dall’idrolisi di LolD viene trasferita a LolC e LolE che la utilizzano per aprire la cavità di LolA in modo che possa legarsi la lipoproteina.

38
Maturazione delle lipoproteine
Tutte le lipoproteine contengono una breve sequenza chiamata LIPOBOX . La lipobox è costituita -(leu, ala, val) -4 -leu-3-(ala, ser) -2-(gly, ala) -1↓ cys+1, E. coli Braun's lipoprotein (OM) MKATKLVLGAVILGSTLLAG↓CSSNA... E. coli lpp-28 (IM) MKLTTHHLRTGAALLLAGILLAG↓CDQSS... Prima che la sequenza segnale venga rimossa un enzima periplasmatico Lgt trasferisce un diacilglicerolo al gruppo SH del residuo di Cysteina ( in posizione +1). Una leader peptidasi specifica rimuove la sequenza segnale e successivamente l’enzima Lnt lega un acido grasso alla cisteina (+1) nei Gram- .
 -4 -leu-3-(ala, ser) -2-(gly, ala) -1↓ cys+1, E. coli Braun s lipoprotein (OM) MKATKLVLGAVILGSTLLAG↓CSSNA... E. coli lpp-28 (IM) MKLTTHHLRTGAALLLAGILLAG↓CDQSS... Prima che la sequenza segnale venga rimossa un enzima periplasmatico Lgt trasferisce un diacilglicerolo al gruppo SH del residuo di Cysteina ( in posizione +1). Una leader peptidasi specifica rimuove la sequenza segnale e successivamente l’enzima Lnt lega un acido grasso alla cisteina (+1) nei Gram- .")
39
Le lipoproteine possono essere associate o alla membrana interna (IN) o a quella esterna (OUT).
Il sistema LOL trasferisce alla OM solo quelle lipoproteine che non possiedono un residuo di acido aspartico (D) in posizione +2.
 in posizione +2.")
40
L’energia derivante dall’idrolisi di ATP mediata da LolD viene trasferita a LolC e LolE che la utilizzano per aprire la cavità di LolA in modo che possa legarsi la lipoproteina Il complesso LolA-lipoproteina attraversa il periplasma e raggiunge LolB, il recettore nella OM Grazie ad una maggiore affinità della lipoproteina per LolB, LolB lega la lipopreina e la inserisce nella ME. Le lipoproteine attraversano la membrana plasmatica grazie al traslocone Sec e subisconola modificazione lipidica a livello di un residuo di cisteina (C) all’N-terminale nel periplasma. Le lipoproteineche possiedono un residuo di aspartato(D) in posizione +2 restano nella membrana interna,mentre quelle che possiedono un altro residuo aminoacidico nella stessa posizione vengono indirizzate alla membrana esterna. L’energia derivata dall’idrolisi dell’ATP viene utilizzata dal complesso LolCDE per trasferire la lipoproteina a LolA. Quando il complesso LolA-lipoproteina interagisce con ilrecettore LolB, la lipoproteina viene trasferita a LolBe inserita sul lato periplasmatico della membrana esterna.
 all’N-terminale nel periplasma. Le lipoproteineche possiedono un residuo di aspartato(D) in posizione +2 restano nella membrana interna,mentre quelle che possiedono un altro residuo aminoacidico nella stessa posizione vengono indirizzate alla membrana esterna. L’energia derivata dall’idrolisi dell’ATP viene utilizzata dal complesso LolCDE per trasferire la lipoproteina a LolA. Quando il complesso LolA-lipoproteina interagisce con ilrecettore LolB, la lipoproteina viene trasferita a LolBe inserita sul lato periplasmatico della membrana esterna.")
Presentazioni simili








>")









 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")
 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")


