Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
NATURA CHIMICA DELL’INFORMAZIONE GENETICA ovvero Come si è arrivati a dimostrare che l’informazione nelle cellule è contenuta nella molecola del DNA

2
L’ESPERIMENTO DI GRIFFITH (1928)
")
3
Che cosa dimostrava l’esperimento di Griffith? che esiste un “principio” trasformante in grado di passare da batteri morti a batteri vivi, trasformandoli dalla forma R (non capsulata e non virulenta) alla forma S (capsulata e virulenta) che tale principio trasformante, una volta acquisito dalle cellule, viene trasmesso anche alla progenie
 alla forma S (capsulata e virulenta) che tale principio trasformante, una volta acquisito dalle cellule, viene trasmesso anche alla progenie.")
4
In che cosa consiste questo “principio trasformante”? L’esperimento di Griffith non dice NULLA sulla natura chimica di questo “principio”, ma è evidente che doveva trattarsi di qualcosa di chimico, cioè una molecola Inoltre, poiché i batteri “trasformati” acquistano una nuova capacità (quella di costruirsi la capsula) e sono anche in grado di trasmetterla alla progenie, la molecola trasmessa deve avere un contenuto “informativo”
 e sono anche in grado di trasmetterla alla progenie, la molecola trasmessa deve avere un contenuto informativo .")
5
In altre parole, non è la torta che viene passata, ma la ricetta per farla ! Tra le molecole che potevano considerarsi adatte (per varieta’ di composizione) a contenere e trasmettere informazione si conoscevano : gli acidi nucleici (DNA e RNA) e le proteine: a) degli acidi nucleici si sapeva che nella loro struttura vi sono quattro varietà di “mattoni” (i nucleotidi), che tuttavia si supponeva si susseguissero nel polimero sempre nello stesso ordine; b) delle proteine, invece, si sapeva che nella loro composizione entravano in gioco venti diversi aminoacidi
 a contenere e trasmettere informazione si conoscevano : gli acidi nucleici (DNA e RNA) e le proteine: a) degli acidi nucleici si sapeva che nella loro struttura vi sono quattro varietà di mattoni (i nucleotidi), che tuttavia si supponeva si susseguissero nel polimero sempre nello stesso ordine; b) delle proteine, invece, si sapeva che nella loro composizione entravano in gioco venti diversi aminoacidi.")
6
Quale dei due tipi di molecole era passato dai batteri S uccisi ai batteri R vivi? L’esperimento di “trasformazione” di Griffith venne ripetuto da altri ricercatori (Avery, Mc Leod, Mc Carthy, 1944) dopo aver trattato gli estratti dei batteri S uccisi, rispettivamente: a) con enzimi in grado di degradare le proteine (proteasi) b) con enzimi in grado di degradare l’RNA (RNAasi) c) con enzimi in grado di degradare il DNA (DNAasi)
 dopo aver trattato gli estratti dei batteri S uccisi, rispettivamente: a) con enzimi in grado di degradare le proteine (proteasi) b) con enzimi in grado di degradare l’RNA (RNAasi) c) con enzimi in grado di degradare il DNA (DNAasi).")
7
L’esperimento di Avery, Mc Leod, Mc Carthy (1944) LA CAPACITA’ TRASFORMANTE ERA CONSERVATA SOLO NEGLI ESTRATTI IN CUI ERA STATO MANTENUTO INTATTO IL DNA
 LA CAPACITA’ TRASFORMANTE ERA CONSERVATA SOLO NEGLI ESTRATTI IN CUI ERA STATO MANTENUTO INTATTO IL DNA")
8
I risultati dell’esperimento di Avery, Mc Leod e Mc Carthy avrebbero dovuto mettere fine alla discussione e stabilire definitivamente che la molecola portatrice di informazione nelle cellule era il DNA…… …...MA NON FU COSI’….

9
Nel 1952 gli esperimenti dei biologi Alfred Hershey e Martha Chase dimostrarono che alcuni virus sono in grado di riprogrammare le cellule ospiti per produrre nuovi virus, iniettando il proprio DNA dentro le cellule. Testa Coda Fibre della coda DNA 300 000

10
Ciclo litico fago T2

11
Fago Batterio Proteina radioattiva DNA DNA del fago Involucri proteici vuoti Radioattività nel liquido Precipitato Si centrifuga Ceppo 1 Proteina radioattiva Ceppo 2 DNA radioattivo DNA radioattivo Si centrifuga Precipitato Radioattività nel precipitato Si mescolano i fagi marcati radioattivamente con i batteri. I fagi infettano le cellule batteriche. 1 Si utilizza un frullatore per separare i fagi esterni ai batteri dalle cellule batteriche e dal loro contenuto. 2 Si centrifuga la miscela. 3 Si misura la radioattività nel precipitato e nel liquido. 4

12
Dunque, il materiale genetico, cioè la molecola capace di dare alle cellule le informazioni necessarie a costruire tutte le strutture e a svolgere le funzioni metaboliche è il DNA

13
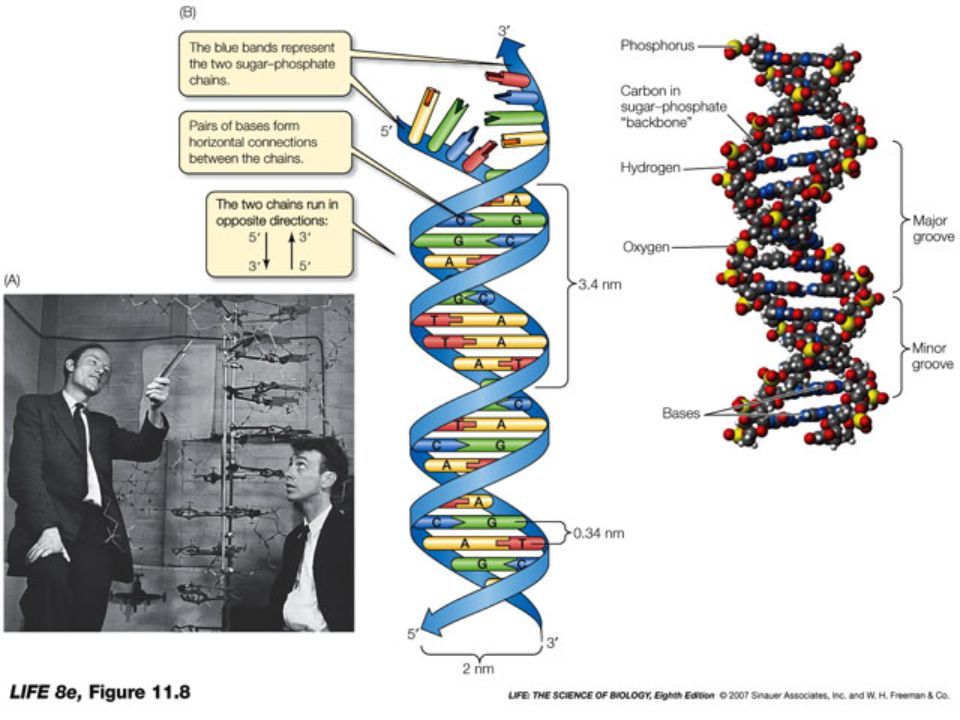
DNA: acido nucleico depositario dell’informazione genetica Nel 1953 Watson e Crick dimostrano che il DNA si duplica da una molecola “stampo” e propongono il primo modello strutturale del DNA.

14
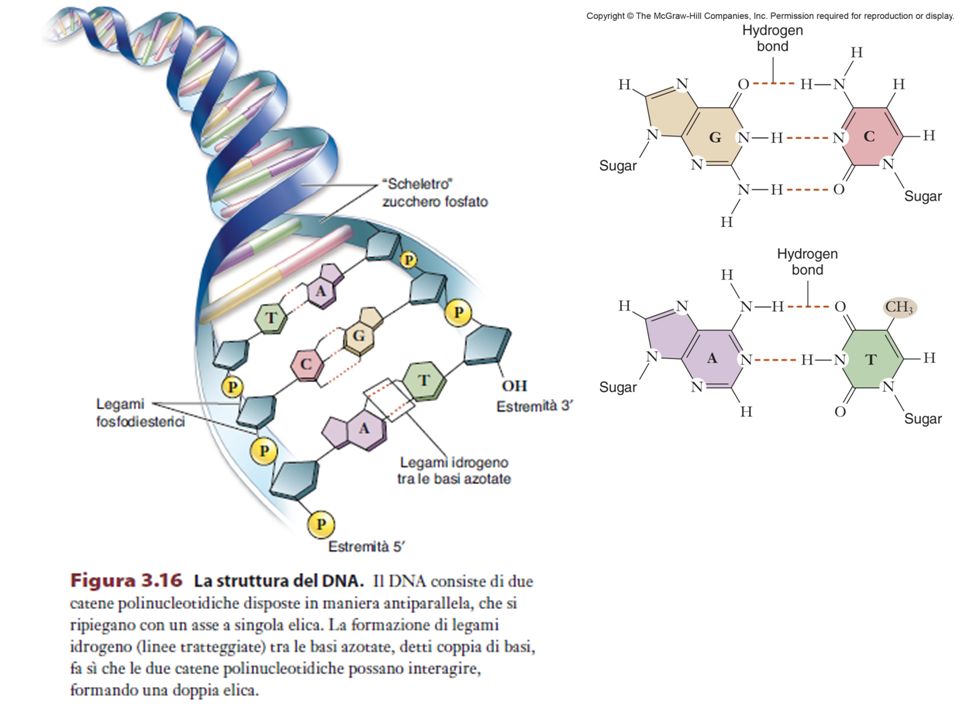
STRUTTURA DEL DNA Dal nucleotide alla doppia elica di Watson e Crick

15
Il DNA è una molecola polimerica i cui monomeri sono i NUCLEOTIDI Ogni NUCLEOTIDE è formato da uno zucchero (un pentoso, cioè uno zucchero a 5 atomi di carbonio), il DESOSSIRIBOSIO, da un gruppo FOSFATO, e da una BASE AZOTATA
, il DESOSSIRIBOSIO, da un gruppo FOSFATO, e da una BASE AZOTATA")
17
Nell’RNA:Nel DNA: Lo zucchero

18
Le basi azotate sono di cinque tipi diversi, e appartengono a due classi di molecole: le purine e le pirimidine: A, G, T, C sono presenti nel DNA A, G, U, C sono presenti nell’RNA La base azotata

19
Organismo Escherichia coli Mycobact. tuberc. Lievito Bue Maiale Uomo A 26.0 15.1 31.7 29.0 29.8 30.4 G 24.9 34.9 18.3 21.2 20.7 19.9 C 25.2 35.4 17.4 21.2 20.7 19.9 T 23.9 14.6 32.6 28.7 29.1 30.1 A/T 1.08 1.03 0.97 1.01 1.02 1.01 G/C 0.99 1.05 1.00 G+C 50.1 70.3 35.7 42.4 41.4 39.8 Py/Pu 1.04 1.00 1.01 Composizione in basi del DNA di varie specie Regola di Chargaff: A=T e G=C

20
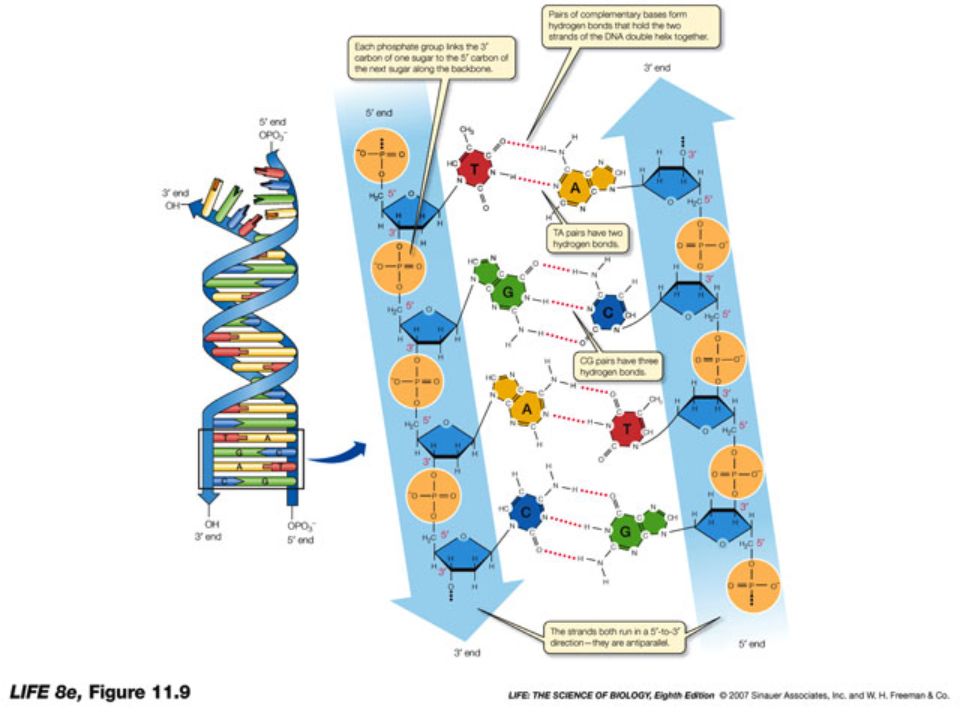
La catena che viene così a formarsi ha una “polarità”, nel senso che le due estremità non sono equivalenti (5’ fosfato da una parte, 3’OH dall’altra)
")
22
Legami idrogeno tra le coppie di basi nel DNA

23
DNAds conservare l’informazione genetica e trasmetterla alla generazione successiva RNAss usare tale informazione per specificare una sequenza amminoacidica

24
Diffrazione dei raggi X del DNA

25
Nel 1953 Watson e Crick propongono il primo modello strutturale del DNA

26
Il DNA è costituito da due catene polinucleotidiche avvolte a formare una doppia elica destrorsa 10 paia di basi

27
Legame forte: nucleotidi concatenati da un legame fosfodiesterico tra il fosfato in posizione 5’ di un nucleotide e il gruppo alcolico in posizione 3’ del nucleotide adiacente Legame debole: legame idrogeno tra basi azotate

28
LEGAMI DEBOLI CHE STABILIZZANO LA DOPPIA ELICA · LEGAMI IDROGENO FRA LE BASI DI OGNI COPPIA: (C-G, tre legami idrogeno, è una coppia più stabile di A-T, due legami idrogeno) · INTERAZIONI IDROFOBICHE: i costituenti idrofili delle basi restano esposti al solvente acquoso solo nei due solchi della doppia elica · FORZE DI VAN DER WAALS fra gli anelli sovrapposti delle basi; si tratta di legami di per sè piuttosto deboli ma cooperativi · INTERAZIONE CON CATIONI: le numerose cariche negative dei fosfati tenderebbero a respingersi rappresentando una causa di instabilità: grazie invece all’interazione con cationi (ioni positivi, cioè, soprattutto Mg 2+, la molecola del DNA, che è unpolianione, viene stabilizzata TUTTI QUESTI LEGAMI, PUR ESSENDO DEBOLI, SONO COOPERATIVI: PERCIO’ LA DOPPIA ELICA DEL DNA E’ UNA STRUTTURA MOLTO STABILE
 · INTERAZIONI IDROFOBICHE: i costituenti idrofili delle basi restano esposti al solvente acquoso solo nei due solchi della doppia elica · FORZE DI VAN DER WAALS fra gli anelli sovrapposti delle basi; si tratta di legami di per sè piuttosto deboli ma cooperativi · INTERAZIONE CON CATIONI: le numerose cariche negative dei fosfati tenderebbero a respingersi rappresentando una causa di instabilità: grazie invece all’interazione con cationi (ioni positivi, cioè, soprattutto Mg 2+, la molecola del DNA, che è unpolianione, viene stabilizzata TUTTI QUESTI LEGAMI, PUR ESSENDO DEBOLI, SONO COOPERATIVI: PERCIO’ LA DOPPIA ELICA DEL DNA E’ UNA STRUTTURA MOLTO STABILE")
29
DENATURAZIONE DEL DNA IN ALCUNI PROCESSI BIOLOGICI FONDAMENTALI, COME LA REPLICAZIONE (O DUPLICAZIONE) E LA TRASCRIZIONE DEL DNA, LA DOPPIA ELICA DEVE APRIRSI, CIOE' LE DUE CATENE DEVONO SEPARARSI. CIO' PUO' ESSERE OTTENUTO ANCHE SPERIMENTALMENTE, SOTTOPONENDO IL DNA A UN TRATTAMENTO CHIMICO O FISICO CHE DETERMINI LA ROTTURA DEI LEGAMI IDROGENO TRA LE BASI DI OGNI COPPIA (ES.: RISCALDANDO IL DNA). QUESTO PROCESSO E' DETTODENATURAZIONE DEL DNA. ESSO PORTA DALLA STRUTTURA ORDINATA A DOPPIA ELICA A UNA STRUTTURA DISORDINATA IN CUI LE DUE CATENE RISULTANO AGGROVIGLIATE IN MODO CASUALE.
. QUESTO PROCESSO E DETTODENATURAZIONE DEL DNA. ESSO PORTA DALLA STRUTTURA ORDINATA A DOPPIA ELICA A UNA STRUTTURA DISORDINATA IN CUI LE DUE CATENE RISULTANO AGGROVIGLIATE IN MODO CASUALE..")
30
Il DNA esiste in diversi tipi di conformazione. Esse sono denominate A-DNA, B-DNA, C-DNA, D-DNA, E-DNA, H- DNA, L-DNA, P-DNA e Z-DNA. Solo A-DNA, B-DNA e Z-DNA sono stati osservati in sistemi biologici naturali. A B Z La forma A è un'ampia spirale destrorsa, con un solco minore largo ma poco profondo ed un solco maggiore più stretto e profondo. Tale conformazione è presente in condizioni non fisiologiche. La conformazione Z è si manifesta invece quando le sequenze che presentano modificazioni chimiche come la metilazione. Essa è caratterizzata da un andamento sinistrorso, opposto rispetto alla conformazione B. Queste strutture inusuali possono essere riconosciute da specifiche proteine, con effetti sulla regolazione della trascrizione.

31
NAD FAD Nicotinammide adenina dinucleotide Flavina adenina dinucleotide Trasportatori di elettroni in molti processi cellulari

32
Per riassumere………

33
1. Struttura del DNA La molecola del DNA è formata da due catene polimeriche, i cui monomeri sono i NUCLEOTIDI Ogni nucleotide è formato da uno zucchero, il 2’- desossiribosio, da una base azotata e da un gruppo fosfato Il gruppo fosfato è legato con un legame estereo all’ossidrile del carbonio 5’ del desossiribosio

34
2. Struttura del DNA La base azotata (purinica: adenina o guanina, oppure pirimidinica: citosina o timina) è legata con legame -N- glicosidico al carbonio 1’ del desossiribosio tramite l’azoto (N) 1 (per le pirimidine) oppure 9 (per le purine) I nucleotidi sono legati tra loro attraverso un legame FOSFODIESTEREO: ogni gruppo fosfato, cioè, forma un secondo legame estere con l’OH del carbonio 3’ di un altro nucleotide: si forma così una catena polinucleotidica ASIMMETRICA, poiché a un’estremità termina con il gruppo 5’-fosfato, e all’altra con il 3’-OH
 è legata con legame -N- glicosidico al carbonio 1’ del desossiribosio tramite l’azoto (N) 1 (per le pirimidine) oppure 9 (per le purine) I nucleotidi sono legati tra loro attraverso un legame FOSFODIESTEREO: ogni gruppo fosfato, cioè, forma un secondo legame estere con l’OH del carbonio 3’ di un altro nucleotide: si forma così una catena polinucleotidica ASIMMETRICA, poiché a un’estremità termina con il gruppo 5’-fosfato, e all’altra con il 3’-OH.")
35
3. Struttura del DNA In tutte le cellule, nei mitocondri e in alcuni virus il DNA è formato da due catene polinucleotidiche antiparallele avvolte a formare una doppia elica destrorsa Le basi planari sono rivolte verso il centro della doppia elica, quasi perpendicolari all’asse dell’elica; zucchero e fosfato, legati con legame fosfodiestereo, costituiscono lo “scheletro” esterno della doppia elica. Se paragoniamo la doppia elica del DNA a una scala a chiocciola, i corrimano sono costituiti dallo scheletro zucchero- fosfato, mentre gli scalini sono formati dalle coppie di basi delle due catene che si trovano allo stesso livello Le basi di ogni coppia sono legate tra loro da legami a idrogeno, che per la geometria dell’elica possono di fatto formarsi soltanto tra le coppie A-T (due legami) e C-G (tre legami). Le due catene risultano perciò COMPLEMENTARI, nel senso che data la sequenza di basi su una catena, quella dell’altra catena è necessariamente determinata.
 e C-G (tre legami). Le due catene risultano perciò COMPLEMENTARI, nel senso che data la sequenza di basi su una catena, quella dell’altra catena è necessariamente determinata..")
37
4. Struttura del DNA La doppia elica può assumere diverse conformazioni: un’elica sinistrorsa, un’elica molto compatta oppure allungata: in realtà il DNA assume quella che risulta essere la conformazione più stabile per la sua composizione e sequenza di basi e per le condizioni (temperatura, forza ionica ecc) in cui si trova. In natura, la conformazione più comune del DNA è quella B, cioè quella descritta da Watson e Crick: una doppia elica destrorsa, con passo d’elica di 3.4 nm e circa 10 coppie di basi per ogni giro completo d’elica (“spessore” di ogni coppia di basi: 0.34 nm) Esistono però altre conformazioni del DNA, con caratteristiche diverse da quelle della conformazione B. Alcune vengono assunte soltanto in determinate condizioni sperimentali, mentre altre (come il DNA Z) sono compatibili con l’ambiente biologico e possono perciò trovarsi, in zone limitate del genoma, dove probabilmente hanno una funzione “regolativa”
 in cui si trova. In natura, la conformazione più comune del DNA è quella B, cioè quella descritta da Watson e Crick: una doppia elica destrorsa, con passo d’elica di 3.4 nm e circa 10 coppie di basi per ogni giro completo d’elica ( spessore di ogni coppia di basi: 0.34 nm) Esistono però altre conformazioni del DNA, con caratteristiche diverse da quelle della conformazione B. Alcune vengono assunte soltanto in determinate condizioni sperimentali, mentre altre (come il DNA Z) sono compatibili con l’ambiente biologico e possono perciò trovarsi, in zone limitate del genoma, dove probabilmente hanno una funzione regolativa .")
39
Topologie del DNA

40
Il DNA può essere distorto come avviene per una corda attraverso un processo definito superavvolgimento. Quando il DNA è in uno stato rilassato, un filamento percorre un giro completo intorno all'asse ogni 10.4 paia di basi. Se invece il DNA è distorto, il numero di basi può aumentare o diminuire. Lo stato di superavvolgimento in cui si trova una molecola di DNA è definito topologia.

41
Se il DNA si avvolge nella direzione dell'elica, si parla di superavvolgimento positivo, con le basi strette tra di loro in modo più marcato. In caso contrario, si parla di superavvolgimento negativo. In natura, la maggior parte delle molecole di DNA presentano un lieve superavvolgimento negativo, introdotto da enzimi definiti topoisomerasi. Questi enzimi sono anche necessari in processi come la trascrizione e la replicazione del DNA, dal momento che sono in grado di risolvere gli stress topologici indotti dai processi stessi.

42
Queste due molecole di DNA sono uguali: hanno lo stesso numero e sequenza di nucleotidi, ma diverse caratteristiche “topologiche”: sono cioè due TOPOISOMERI della stessa molecola. quello a sinistra è però instabile, poiché in esso sono alterate le caratteristiche della doppia elica B

43
STRUTTURA DELLA CROMATINA Il DNA eucariotico è complessato con proteine basiche

44
Organizzazione del DNA eucariotico nei cromosomi I DNA batterici sono circolari ed hanno una sola origine di replicazione I cromosomi eucariotici sono costituiti ciascuno da una molecola di DNA lineare con molte origini di replicazione Il DNA batterico è associato con poliamine Il DNA eucariotico è associato con proteine (istoni) a formare la cromatina, e con proteine non istoniche
 a formare la cromatina, e con proteine non istoniche")
45
I nucleosomi sono complessi formati da un ottamero istonico e DNA 146 bp 60 bp Ottamero istonico formato da 4 coppie dell’istone H2A, H2B, H3 e H4 Istoni ricchi in Lys e Arg Sequenze molto conservate ruolo cruciale

46
Cromatina “a collana di perle” (fibra da 10 nm)
")
47
Il modello a “solenoide” della cromatina condensata (presente durante interfase)
")
48
Un modello per un’organizzazione gerarchica dei livelli di compattazione della cromatina nei cromosomi

49
Proteine non istoniche forniscono un’impalcatura strutturale per lunghe anse di cromatina nei cromosomi Un cromosoma è formato all’incirca da 40% DNA e 60% proteine

50
Nel cromosoma mitotico la lunghezza di ogni molecola di DNA è circa 10.000 volte inferiore

51
Morfologia ed elementi funzionali dei cromosomi eucariotici Il numero e la forma dei cromosomi sono caratteristici di ogni specie animale e vegetale Tre elementi funzionali sono richiesti per la replicazione e l’ereditabilità stabile dei cromosomi eucariotici: origini di replicazione centromero le due estremità (telomeri) centromero telomero
 centromero telomero")
52
Nelle nostre cellule il corredo genetico è diploide (2n) ed è formato da coppie di cromosomi omologhi, di origine materna e paterna, tranne che nei gameti dove il corredo è aploide (n) Per garantire che ogni cellula figlia riceva un set completo di cromosomi, i cromatidi fratelli che si formano dopo la replicazione rimangono uniti fino alla segregazione cromosomica.
 ed è formato da coppie di cromosomi omologhi, di origine materna e paterna, tranne che nei gameti dove il corredo è aploide (n) Per garantire che ogni cellula figlia riceva un set completo di cromosomi, i cromatidi fratelli che si formano dopo la replicazione rimangono uniti fino alla segregazione cromosomica.")
53
La cromatina è una struttura dinamica ed esiste in forme più o meno condensate. Durante l’interfase l’eucromatina non è condensata e viene espressa Cromatina “a collana di perle” o fibra da 10 nm Struttura a solenoide o fibra da 30 nm

54
L’eterocromatina è costituita da regioni che non si decondensano mai e non vengono mai trascritte (costitutiva). Alcune porzioni del DNA vengono trascritte solo in certi tipi cellulari e/o solo in alcuni momenti (eterocromatina facoltativa)
.")
55
DNA degli organelli I mitocondri contengono diverse molecole di DNA I geni contenuti nel DNA mitocondriale codificano per rRNA, tRNA mitocondriali e per alcune proteine mitocondriali Le dimensioni e il contenuto codificante dei DNA mitocondriali variano notevolmente nei diversi organismi I prodotti dei geni mitocondriali non vengono esportati (restano cioè nel mitocondrio)
")
Presentazioni simili









 è stato un biologo svizzero, che isolò per la prima volta gli acidi nucleici. Egli evidenziò.>")









. Chimica del DNA i). Forze che influenzano la stabilità della doppia elica del DNA interazioni idrofobiche - stabilizzano dentro idrofobiche e fuori.>")
