Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Andrea Camperio Ciani dip di Psicologia Generale Universita’ di Padova
Identita’ di genere basi neurobiologiche biologiche della sessualita’ e dell’orientamento sessuale Andrea Camperio Ciani dip di Psicologia Generale Universita’ di Padova

2
L’ACCOPPIAMENTO NEI RATTI
I ratti sono attratti l’un l’altro prevalentemente attraverso gli odori. Le femmine sono ovulatrici spontanee, cioè, anche quando sono lasciate sole esse ovulano ogni quattro o cinque giorni. Per alcune ore durante ciascun ciclo, la femmina cerca un maschio e mostra comportamenti procettivi, che incitano il maschio a montarla. Se la femmina è recettiva, sta ferma e assume una postura che favorisce l’intromissione: alza la testa e la groppa e sposta la coda da un lato; questa posizione è chiamata lordosi.

3
CIRCUITI NEURALI NELLA RIPRODUZIONE DEI RODITORI
Nella femmina, l’ipotalamo ventromediale (VMH) è cruciale per la lordosi, poiché le lesioni in tale sede aboliscono la risposta. L’VMH invia assoni alla sostanza grigia del mesencefalo, i neuroni della sostanza grigia periacqueduttale proiettano alla formazione reticolare del midollo allungato, che a sua volta proietta al midollo spinale attraverso il tratto reticolospinale. Il ruolo dell’VMH è di monitorare la concentrazione di ormone steroideo e, al momento giusto dell’ovulazione, attivare una via multisinaptica che induca il midollo spinale a mostrare una risposta di lordosi altrimenti assente.
 è cruciale per la lordosi, poiché le lesioni in tale sede aboliscono la risposta. L’VMH invia assoni alla sostanza grigia del mesencefalo, i neuroni della sostanza grigia periacqueduttale proiettano alla formazione reticolare del midollo allungato, che a sua volta proietta al midollo spinale attraverso il tratto reticolospinale. Il ruolo dell’VMH è di monitorare la concentrazione di ormone. steroideo e, al momento giusto dell’ovulazione, attivare una via multisinaptica che induca il midollo spinale a mostrare una risposta di lordosi altrimenti assente.")
4
CIRCUITI NEURALI NELLA RIPRODUZIONE DEI RODITORI
Nel maschio, l’area preottica mediale dell’ipotalamo (POA) è ricca di neuroni sensibili agli steroidi e le lesioni in tale area aboliscono il comportamento copulatorio nei maschi. I neuroni di quest’area mandano i loro assoni al mesencefalo mediale attraverso il fascio mediale del proencefalo, da qui l’informazione va ai nuclei della base e, attraverso una via multisinaptica, al midollo spinale che, a sua volta, media i vari riflessi di intromissione ed eiaculazione. Quando il testosterone attiva il POA in un ratto maschio adulto, lo fa dopo essere stato convertito in estrogeno, che agisce poi sui recettori per gli estrogeni. Nella periferia e nel midollo spinale i riflessi di intromissione ed eiaculazione sono indotti dall’attivazione dei recettori per gli androgeni. L’organo vomeronasale del maschio è in grado di rilevare i feromoni delle femmine recettive che eccitano il maschio. L’informazione vomeronasale raggiunge l’amigdala mediale che manda a sua volta assoni al POA.
 è ricca di neuroni sensibili agli steroidi e le lesioni in tale area aboliscono il comportamento copulatorio nei maschi. I neuroni di quest’area mandano i loro assoni al mesencefalo mediale attraverso il fascio mediale del proencefalo, da qui l’informazione va ai nuclei della base e, attraverso una via multisinaptica, al midollo spinale che, a sua volta, media i vari riflessi di intromissione ed eiaculazione. Quando il testosterone attiva il POA in un ratto maschio adulto, lo fa dopo essere stato convertito in estrogeno, che agisce poi sui recettori per gli estrogeni. Nella periferia e nel midollo spinale i riflessi di. intromissione ed. eiaculazione sono indotti dall’attivazione dei recettori per gli androgeni. L’organo vomeronasale del maschio è in grado di rilevare i feromoni delle femmine recettive che eccitano il maschio. L’informazione vomeronasale raggiunge l’amigdala mediale che manda a sua volta assoni al POA.")
5
ANDROGENI E COMPORTAMENTO COPULATORIO MASCHILE
Gli ormoni giocano un ruolo importante nei comportamenti di accoppiamento del ratto. Il testosterone ha un effetto di attivazione sul comportamento, ma il livello ormonale da solo non determina completamente la quantità del comportamento sessuale. In seguito alla castrazione l’attività sessuale dei porcellini d’India maschio decresce; quindi viene somministrata a ciascun animale la stessa quantità di testosterone. Ogni gruppo è tornato al livello di attività sessuale che mostrava prima della castrazione, ma in nessun gruppo aumenta l’attività di accoppiamento in seguito al raddoppio della dose di ormone. Qualcos’altro sembra essere responsabile delle differenze individuali nell’attività di accoppiamento (stalloni e mezze cartucce). “stalloni” (maggiore frequenza di accoppiamento) “mezze cartucce” (minore frequenza di accoppiamento)
. stalloni (maggiore frequenza di accoppiamento) mezze cartucce (minore frequenza di accoppiamento)")
6
IL CICLO DELL’OVULAZIONE NEL RATTO
L’estrogeno secreto all’inizio del ciclo di ovulazione del ratto femmina facilita il suo comportamento procettivo e la susseguente produzione di progesterone accresce il comportamento procettivo e permette la lordosi. Se ad una femmina adulta vengono rimosse le ovaie, essa mostrerà recettività solo se trattata prima con estrogeno e poi con progesterone, perché l’estrogeno stimola il cervello ad attivare i recettori del progesterone; senza il pretrattamento con estrogeno, il cervello non può rispondere al progesterone.

7
ANATOMIA DELL’APPARATO RIPRODUTTIVO UMANO
Nelle donne le ovaie producono uova più o meno ogni 28 giorni. L’uovo rilasciato cade nell’adiacente tromba di falloppio e da qui all’utero (fig. a). Nei maschi gli spermatozoi vengono prodotti nei testicoli e maturano nell’adiacente epididimo (fig. b).
. Nei maschi gli spermatozoi vengono prodotti nei testicoli e maturano nell’adiacente epididimo (fig. b).")
8
I CICLI DELLA RISPOSTA SESSUALE UMANA e sua variabilita’
Masters e Johnson (1965) hanno riassunto i modelli delle risposte tipiche degli uomini e delle donne come consistenti in quattro fasi: 1. Crescita dell’eccitamento 2. Plateau 3. Orgasmo 4. Risoluzione Mentre gli uomini hanno un solo modello fondamentale (fig. a), le donne hanno tre modelli tipici (fig. b). Questi modelli sono molto schematici e variano da un individuo all’altro. Il tipico modello maschile ha un periodo refrattario assoluto successivo all’orgasmo, mentre le donne possono avere orgasmi multipli in rapida successione.
 hanno riassunto i modelli delle risposte tipiche degli uomini e delle donne come consistenti in quattro fasi: 1. Crescita dell’eccitamento. 2. Plateau. 3. Orgasmo. 4. Risoluzione. Mentre gli uomini hanno un solo modello fondamentale (fig. a), le donne hanno tre modelli tipici (fig. b). Questi modelli sono molto schematici e variano da un individuo all’altro. Il tipico modello maschile ha un periodo refrattario assoluto successivo all’orgasmo, mentre le donne possono avere orgasmi multipli in rapida successione.")
9
DETERMINAZIONE DEL GENERE SESSUALE
La differenziazione sessuale è il processo attraverso il quale gli individui sviluppano corpi e comportamenti o maschili o femminili. Il sesso nei mammiferi è determinato al momento del concepimento, quando uno spermatozoo penetra l’uovo e offre un cromosoma Y o X. Il maschio è eterogametico perché ha due cromosomi diversi (XY), mentre la femmina è omogametica perché ha due cromosomi uguali (XX).
, mentre la femmina è omogametica perché ha due cromosomi uguali (XX).")
10
LA DIFFERENZIAZIONE SESSUALE
Nelle fasi iniziali dello sviluppo ciascun individuo ha un paio di gonadi indifferenziate. La determinazione sessuale, cioè i primi eventi che decidono se l’individuo diventerà maschio o femmina, inizia con l’espressione o non della proteina Sry, prodotta dalla gonade indifferenziata alla presenza del gene Sry, contenuto nel cromosoma Y. Con la produzione della proteina Sry la gonade indifferenziata si trasforma in testicolo; in assenza della proteina Sry la gonade indifferenziata si trasforma in ovaia.

11
Processo di differenziazione sessuale durante l’ontogenesi

12
MECCANISMI GENETICI E ORMONALI DELLA DIFFERENZIAZIONE SESSUALE

13
EVENTI DELLA DIFFERENZIAZIONE SESSUALE
I testicoli in sviluppo producono diversi ormoni, mentre le ovaie inizialmente producono pochi ormoni. Se le cellule dell’embrione ricevono gli ormoni testicolari, cominciano a sviluppare caratteri maschili; se le cellule non sono esposte agli ormoni testicolari, sviluppano caratteri femminili. Quindi, il ruolo dei cromosomi è quello di determinare il sesso delle gonadi, mentre gli ormoni delle gonadi poi guidano la differenziazione sessuale del resto del corpo.

14
L’IPOTESI DELL’AROMATIZZAZIONE
Nell’aromatizzazione, l’enzima aromatasi converte il testosterone (androgeno) in estradiolo (estrogeno). Le ovaie ed il cervello normalmente contengono una grande quantità di aromatasi. Secondo l’ipotesi dell’aromatizzazione, gli androgeni testicolari entrano nel cervello e sono convertiti in estrogeni che a loro volta mascolinizzano il sistema nervoso in via di sviluppo. La proteina alfa-fetoproteina chela solo gli estrogeni e ne previene l’ingresso nel cervello (non gli androgeni) ; quindi gli androgeni (testosterone) prodotti dai testicoli entrano nel cervello, vengono convertiti dall’aromatasi in estrogeni e si legano ai suoi recettori (Re) regolando l’espressione genica in modo maschile.
 in estradiolo (estrogeno). Le ovaie ed il cervello normalmente contengono una grande quantità di aromatasi. Secondo l’ipotesi dell’aromatizzazione, gli androgeni testicolari entrano nel cervello e sono convertiti in estrogeni che a loro volta mascolinizzano il sistema nervoso in via di sviluppo. La proteina alfa-fetoproteina chela solo gli estrogeni e ne previene l’ingresso nel cervello (non gli androgeni) ; quindi gli androgeni (testosterone) prodotti dai testicoli entrano nel cervello, vengono convertiti dall’aromatasi in estrogeni e si legano ai suoi recettori (Re) regolando l’espressione genica in modo maschile.")
15
UN FENOTIPO “INTERSESSUALE”
Alcune persone hanno un solo cromosoma sessuale, un singolo X; tale struttura genetica (XO) ha come risultato la sindrome di Turner: un individuo di aspetto femminile con ovaie poco sviluppate, ma riconoscibili. Nell’iperplasia surrenale congenita (CAH), le ghiandole surrenali, che normalmente producono piccole quantità di steroidi androgeni, producono invece una quantità di androgeno considerevole, solitamente intermedi tra i livelli dei maschi e delle femmine normali, provocando un aspetto “intersessuale” (vedi figura).
 ha come risultato la sindrome di Turner: un individuo di aspetto femminile con ovaie poco sviluppate, ma riconoscibili. Nell’iperplasia surrenale congenita (CAH), le ghiandole surrenali, che normalmente producono piccole quantità di steroidi androgeni, producono invece una quantità di. androgeno considerevole, solitamente intermedi tra i livelli dei maschi e delle femmine normali, provocando un aspetto intersessuale (vedi figura).")
16
L’INSENSIBILITA’ ALL’ANDROGENO
Nell’insensibilità all’androgeno, un individuo XY, ha il gene del recettore dell’androgeno difettoso, quindi è incapace di produrre il recettore dell’androgeno e di conseguenza è incapace di rispondere agli ormoni androgeni. Il suo corpo presenta i testicoli, ma l’insensibilità all’androgeno sviluppa un aspetto femminile (vedi figura).
.")
17
un paradosso darwiniano
L’omosessualità: un paradosso darwiniano Prof. Andrea Camperio Ciani Facoltà di Psicologia -Università di Padova-

18
Comportamenti omosessuali sono stati riscontrati in oltre 450 specie animali (Bagemihl, 1999)
Alcuni esempi….. UCCELLI Nei Pipistrelli vampiro sono state osservate erezioni durante “grooming” e pulizia reciproca tra soggetti dello stesso genere sessuale MAMMIFERI Nei leoni, sono stati spesso osservati comportamenti omosessuali e promiscui. Nelle giraffe i maschi mostrano prolungate manifestazioni di affetto reciproco attraverso lo strofinamento dei colli, spesso poi si montano reciprocamente e sembrano raggiungere l’orgasmo

19
PRIMATI comportamenti omosessuali sono stati osservati nelle macache (monte reciproche tra maschi, intromissioni anali e sesso orale), nei gorilla (corteggiamenti reciproci tra individui dello stesso sesso, più spesso omosessualità femminile), nei bonobo (tra femmine: strofinamento reciproco dei genitali), negli scimpanzé (comune sia tra maschi che tra femmine) e nelle macache Rhesus (monte reciproche tra individui dello stesso sesso, sia maschi che femmine)
, nei gorilla (corteggiamenti reciproci tra individui dello stesso sesso, più spesso omosessualità femminile), nei bonobo (tra femmine: strofinamento reciproco dei genitali), negli scimpanzé (comune sia tra maschi che tra femmine) e nelle macache Rhesus (monte reciproche tra individui dello stesso sesso, sia maschi che femmine)")
20
Ipotesi fisiologiche Disequilibrio ormonale Studi di Resko (1996)
Deficit nella produzione di ormoni (testosterone, estrogeno, estradiolo) o ridotta biosintesi di tali ormoni possono portare all’espressione del comportamento omosessuale Regione Cerebrale L’Area preottico- anteriore dell’Ipotalamo è legata al comportamento riproduttivo del maschio in molti vertebrati. Studi su gatti,cani, capre e scimmie confermano che una lesione di tale area può provocare deficit nel comportamento copulatorio e comportamenti omosessuali
 o ridotta biosintesi di tali ormoni possono portare all’espressione del comportamento omosessuale. Regione Cerebrale. L’Area preottico- anteriore dell’Ipotalamo è legata al comportamento riproduttivo del maschio in molti vertebrati. Studi su gatti,cani, capre e scimmie confermano che una lesione di tale area può provocare deficit nel comportamento copulatorio e comportamenti omosessuali.")
21
Ipotesi Sociali Le ipotesi sociali avanzate per spiegare l’omosessualità animale si possono suddividere in due grandi tipologie: Quelle che partono dal presupposto che l’omosessualità sia un comportamento finalizzato al benessere ed allo sviluppo del singolo individuo Quelle che spiegano l’omosessualità come un comportamento del singolo individuo finalizzato al benessere del gruppo (GROUP SELECTION) Queste due tipologie di ipotesi risultano essere, comunque, complementari, reciprocamente legate e non mutuamente esclusive
 Queste due tipologie di ipotesi risultano essere, comunque, complementari, reciprocamente legate e non mutuamente esclusive.")
22
Omosessualità: Come “fase di apprendimento”, finalizzata all’acquisizione della pratica sessuale eterosessuale. Questo spiegherebbe perché risulta essere più frequente negli individui giovani Come “risultato dell’imprinting nella sfera sessuale”. Un animale potrebbe nascere senza preferenze sessuali ed aver bisogno di una forma di imprinting. Esperienze precoci con partner dello stesso sesso potrebbero favorire una preferenza omosessuale Come “imitazione del comportamento eterosessuale” e finalizzato al comportamento eterosessuale. In questo modo l’individuo potrebbe stimolare sessualmente individui del sesso opposto Come “gioco promiscuo” tra maschi e femmine che potrebbe poi trasformarsi in attività sessuale eterosessuale Come “modalità per ottenere piacere sessuale” Come “fonte di distensione” finalizzato limitare gli effetti distruttivi della competizione e limitare i comportamenti aggressivi

23
Differenze di genere nelle fantasie sessuali La prospettiva della psicologia evoluzionistica: i transessuali?

24
Ipotesi darwiniana della selezione sessuale
I maschi competono fra loro per accoppiarsi con le femmine (selezione intrasessuale) Le femmine scelgono i maschi migliori selezione intersessuale…. Quindi gli uomini sono piu’ promiscui e le femmine piu’ selettive La poliginia avvantaggia i maschi la monogamia avvantaggia le femmine I maschi se devono scelgono le femmine sulla base delle caratteristiche fisiche che richiamano la giovinezza la salute e quindi la fecondita’ Le femmine scelgono i maschi in base ai loro buoni geni o alle prospettive di investire al meglio nella famiglia
 Le femmine scelgono i maschi migliori selezione intersessuale…. Quindi gli uomini sono piu’ promiscui e le femmine piu’ selettive. La poliginia avvantaggia i maschi la monogamia avvantaggia le femmine. I maschi se devono scelgono le femmine sulla base delle caratteristiche fisiche che richiamano la giovinezza la salute e quindi la fecondita’ Le femmine scelgono i maschi in base ai loro buoni geni o alle prospettive di investire al meglio nella famiglia.")
25
Differenze legate al genere nei contenuti delle fantasie sessuali(Ellis e Symons ’90)
Varieta’ di partner sessuali:gli uomini piu’ delle donne coinvolgono partners sconosciuti e piu’ partners II maschi preferiscono immagini visive sessualmente esplicite verso immagini emotivo-romantiche Le femmine preferiscono un ruolo passivo piuttosto che attivo nella fantasia Le fantasie di sottomissione sono piu’ frequenti nelle femmine, negli uomini quelle di dominanza

28
Il nostro studio replica di Ellis e Symons ’90
Anche per il campione italiano sia adulto che giovane presenta gli stessi risultati Sia per la varieta’ sessuale Sia per le immagini visive verso contatto fisico, importanza del contesto e caratteristiche personali

29
Differenze di genere nella qualificazione dell’investimento parentale (Kenrick et al, ’90)
Gli autori suggeriscono di qualificare l’investimento parentale in rapporto a diversi livelli di relazione ipotizzando che: Al primo appuntamento l’investimento parentale atteso dovrebbe essere basso per entrambi i sessi a livello di relazione occasionale l’investimento potenziale della donna dovrebbe crescere incrementando la selettivita’ A livello di relazione stabile anche l’investimento dell’uomo dovrebbe crescere sostanzialmente con un incremento della selettivita’ A livello di matrimonio o convivenza l’investimento parentale diverrebbe molto consistente alzando la selettivita’ in entrambi

30
*

31
* ** *femmine italiane meno selettive negli incontri occasionali rispetto alle attese ** femmine italiane piu’ selettive dei maschi negli incontri occasionali *

32
**

33
Confronto adulti studenti
**

34
* **

35
*

36
La nostra ricerca Contrariamente a quanto riscontrato nel campione US (Kenrick et al. ’90) Negli italiani non si rivela l’atteso incremento della selettivita’ delle donne, sia giovani che adulte, a livello di relazione occasionale, se non per salute ed attrattiva fisica Per gli altri risultati non ci sono differenze con il campione US
 Negli italiani non si rivela l’atteso incremento della selettivita’ delle donne, sia giovani che adulte, a livello di relazione occasionale, se non per salute ed attrattiva fisica. Per gli altri risultati non ci sono differenze con il campione US.")
37
Identita’ di genere nei transessuali MvF
Introduzione Differenze di genere fra maschi e femmine Livelli di investimento nei due sessi Criteri di scelta del partner nei due sessi Definizione di transessuale MvF come viene visto nella psicologia dinamica (una psiche femminile imprigionata in un corpo maschile)
")
38
L’indagine sul campo 272 studenti italiani (146 femmine e 126 maschi) tutti eterosessuali 30 soggetti transessuali MvF due questionari: selettivita’ nella scelta del partner in base al livello di investimento fantasie sessuali

39
Principali differenze fra sessi nelle fantasie sessuali emerse nello studio

40
Principali differenze fra sessi nella selettivita’ emerse nello studio

41
Confronto fra maschi femmine e transessuali nella Selettivita’ nella scelta del partner

42
Confronto fra maschi femmine e transessuali nella Selettivita’ nella scelta del partner

43
Confronto fra maschi femmine e transessuali nella Selettivita’ nella scelta del partner

44
I transessuali MvF erano dunque donne intrappolate in un corpo maschile?

45
Dove esiste uno stereotipo noto come nella selezione del partner i transessuali si adeguano al profilo femminile dove lo stereotipo non e’ noto i transessuali sembrano mantenere l’identita’ di genere originaria e maschile

46
Quali fattori influenzano l’orientamento sessuale nell’essere umano
Ipotesi psicosociali: Teoria dell’effetto contagio (Archer 1997) Contatti ed abusi in fase evolutiva precoce con Uomini omosessuali adulti Evitamento maestri e pediatri Si prevede alto tasso di abusi o contatti sessuali omosessuali precoci con adulti Ipotesi di Storms (1981) maturazione sessuale anticipata Gli omosessuali adulti sembrano essere, nel periodo infantile, più precoci nella sfera sessuale e riportano più contatti precoci con compagni dello stesso sesso Si prevede che riportino elevato numero di contatti sessuali prima dei dieci anni rispetto agli eterosessuali
 Contatti ed abusi in fase evolutiva precoce con Uomini omosessuali adulti. Evitamento maestri e pediatri. Si prevede alto tasso di abusi o contatti sessuali omosessuali precoci con adulti. Ipotesi di Storms (1981) maturazione sessuale anticipata. Gli omosessuali adulti sembrano essere, nel periodo infantile, più precoci nella sfera sessuale e riportano più contatti precoci con compagni dello stesso sesso. Si prevede che riportino elevato numero di contatti sessuali prima dei dieci anni rispetto agli eterosessuali.")
47
le indagini genetiche:
Esistono basi biologiche che influenzano l’orientamento omosessuale? (parte seconda) le indagini genetiche: Bailey e la familiarita’ Hamer il gene Xq28, e la linea materna la Fisiologia dello sviluppo l’ipotesi dell’ordine di nascita di Blanchard
 le indagini genetiche: Bailey e la familiarita’ Hamer il gene Xq28, e la linea materna. la Fisiologia dello sviluppo. l’ipotesi dell’ordine di nascita di Blanchard.")
48
L’Ipotesi della familiarita’ di Bailey (‘91,’95,’98)
Studi sui gemelli omozigoti, dizigoti, e fratelli Gli omozigoti hanno maggiori concordanze di omosessualità rispetto agli altri Si prevedono alti tassi di omosessualità nelle famiglie dei soggetti omosessuali indistintamente fra linea materna e linea paterna

49
Ipotesi genetica di Hamer (‘95)
Mediante lo studio della familiarita’verifica un maggior tasso di omosessualita’ maschile nella linea materna degli omosessuali Attraverso DNA linkage analysis verifica un possibile legame fra orientamento omosessuale e la regione Xq28 nel cromosoma X Si prevede un piu’ alto tasso di omosessualita’ lungo la linea materna nei soggetti omosessuali

50
L’ipotesi immunologica di Blanchard (’96)
reazione immunologica della madre nei confronti del H-Y antigene presentato da ciascun feto di sesso maschile Memoria immunologica accentua la reazione dopo ogni gravidanza maschile, riducendo il processo di mascolinizzazione cerebrale nel feto nella fase critica dello sviluppo Si prevede che gli omossessuali riportino un ordine di nascita piu’ elevato rispetto agli eterosessuali Possiedono un numero di fratelli maschi maggiori superiori alla media degli eterosessuali

51
Lo studio di LeVay (1991) Attraverso uno studio su un campione di donne, uomini eterosessuali ed omosessuali, LeVay riportò che l’area Preottica degli esseri umani contiene un nucleo, il terzo nucleo interstiziale dell’ipotalamo anteriore (INAH-3), che è più grande negli uomini che nelle donne ed è più grande negli uomini eterosessuali che negli uomini omosessuali Non è stato, da LeVay, chiarito se tale porzione fosse più piccola già dalla nascitaaspetto innato dell’omosessualità O se le esperienze sessuali da adulto influenzassero la struttura dell’INAH-3omosessualità legata all’esperienza
, che è più grande negli uomini che nelle donne ed è più grande negli uomini eterosessuali che negli uomini omosessuali. Non è stato, da LeVay, chiarito se tale porzione fosse più piccola già dalla nascitaaspetto innato dell’omosessualità. O se le esperienze sessuali da adulto influenzassero la struttura dell’INAH-3omosessualità legata all’esperienza.")
52
DIMORFISMO SESSUALE NELL’IPOTALAMO
Nell’area preottica (POA) dell’ipotalamo sono state identificate differenze sessuali relativamente al numero di sinapsi. Il dimorfismo sessuale nel sistema nervoso riguarda differenze relative al numero, grandezza e forma dei neuroni e al numero di sinapsi. Nei ratti il nucleo sessualmente dimorfico del POA (SDN-POA) è tanto più ampio nei maschi rispetto alle femmine.
 dell’ipotalamo sono state identificate differenze sessuali relativamente al numero di sinapsi. Il dimorfismo sessuale nel sistema nervoso riguarda differenze relative al numero, grandezza e forma dei neuroni e al numero di sinapsi. Nei ratti il nucleo sessualmente dimorfico del POA (SDN-POA) è tanto più ampio nei maschi rispetto alle femmine.")
53
CHE COSA DETERMINA L’ORIENTAMENTO SESSUALE ?
L’area preottica (POA) degli esseri umani contiene un nucleo (INAH-3) che è più grande negli uomini che nelle donne ed è più grande negli uomini eterosessuali che negli uomini omosessuali. Gli psicobiologi, sanno che l’esperienza precoce può alterare la struttura del cervello per avere un’influenza sul comportamento successivo. E’ possibile che questa esperienza sociale influisca sullo sviluppo dell’INAH-3 per determinare più tardi l’orientamento sessuale. Le stime indicano che la variabilità nell’orientamento sessuale è dovuta solo per metà a differenze nel genoma, il che lascia ampio spazio alle prime influenze sociali. Quindi, sia le influenze genetiche che quelle ambientali contribuiscono a determinare l’orientamento sessuale degli esseri umani.
 degli esseri umani contiene un nucleo (INAH-3) che è più grande negli uomini che nelle donne ed è più grande negli uomini eterosessuali che negli uomini omosessuali. Gli psicobiologi, sanno che l’esperienza precoce può alterare la struttura del cervello per avere. un’influenza sul comportamento successivo. E’ possibile che questa esperienza sociale influisca sullo sviluppo dell’INAH-3 per determinare più tardi l’orientamento sessuale. Le stime indicano che la variabilità nell’orientamento sessuale è dovuta solo per metà a differenze nel genoma, il che lascia ampio spazio alle prime influenze sociali. Quindi, sia le influenze genetiche che quelle ambientali contribuiscono a determinare l’orientamento sessuale degli esseri umani.")
54
(i dati raccolti da LeVay sono stati messi in discussione perché il campione era costituto da cadaveri di uomini omosessuali morti di AIDS - il virus stesso potrebbe aver modificato le dimensioni di diverse aree cerebrali) INAH-3 in uomo eterosessuale INAH-3in uomo omosessuale

55
L’indagine sul campo 98 soggetti omosessuali maschi e tutti i loro familiari per due generazioni 100 soggetti eterosessuali di identica struttura e provenienza e tutti i familiari per due generazioni questionari: orientamento sessuale (scala Kinsey) ed ordine di nascita numerosita’ della linea materna ed orientamento sessuale numerosita’ della linea paterna ed orientamento sessuale
 ed ordine di nascita. numerosita’ della linea materna ed orientamento sessuale. numerosita’ della linea paterna ed orientamento sessuale.")
56
Risultati Come riportato da Kinsey (’48) l’orientamento sessuale maschile tende a riprodurre un bipolarismo (omosessualita’- eterosessualita’) ed a mostrare basse percentuali di bisessualita’ 0 eterosessuale omosessuale 6
 l’orientamento sessuale maschile tende a riprodurre un bipolarismo (omosessualita’- eterosessualita’) ed a mostrare basse percentuali di bisessualita’ 0 eterosessuale 3 4 omosessuale 6.")
57
Risultati si conferma l’ipotesi di Hamer - il problema della sovrastima o sottostima nel campione riportato (Bailey) ** P<0.001 **

58
Risultati II Si conferma l’ipotesi di Blanchard
P<.02 con Italia P<.05 con il controllo

59
L’analisi delle frequenze di abusi e di approcci sessuali precoci con adulti non confermano l’ipotesi dell’effetto contagio di Archer Abusi subiti L’analisi della frequenza degli approcci intimosessuali avuti prima dei dieci anni conferma l’ipotesi della maturazione sessuale anticipata di Storms Approcci precoci con chi eta’ Con chi sesso

60
Ma quale fattore conta maggiormente nell’influenzare l’orientamento sessuale?
- Nota Il modello spiega solo il 21% della varianza il resto e’ da spiegare

61
Il paradosso: come si mantiene nella popolazione un carattere parzialmente genetico che riduce la fecondita’ di chi lo possiede?

62
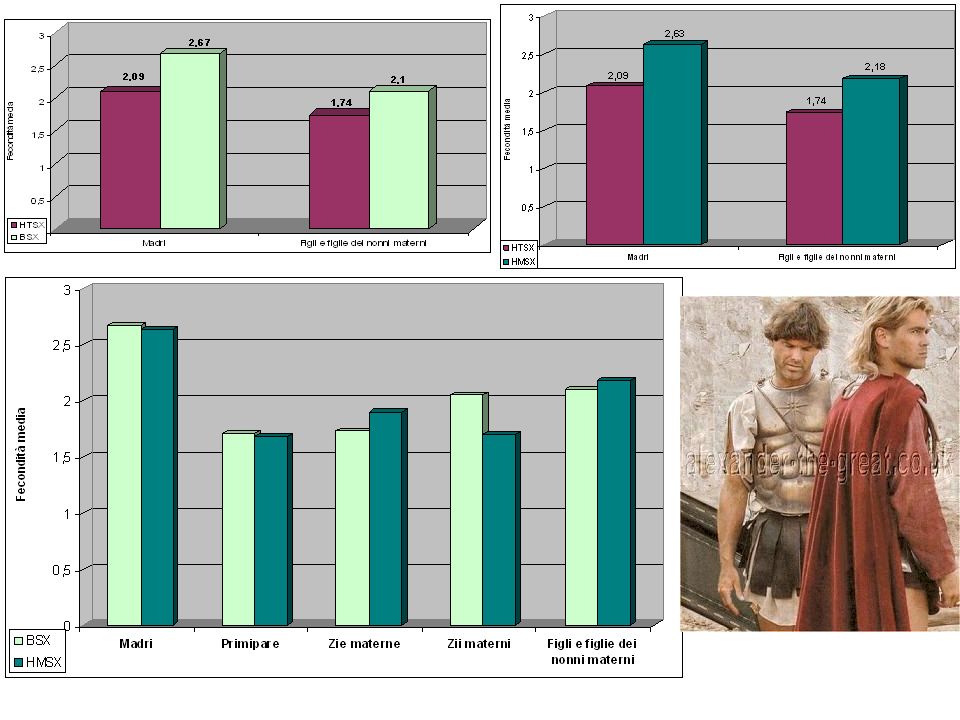
Aumentando la fecondita’ delle femmine in linea materna portatrici del fattore genetico
** P<.04 * P<.02 ** P<.001

63
Quale modello genetico spiega meglio i risultati ottenuti sulle basi genetiche dell’omosessualita’?

64
Quindi….. L’orientamento sessuale non modifica completamente le caratteristiche dell’identita’ del genere biologico esistono delle basi genetiche e fisiologiche dell’omosessualita’ che si mantengono nella popolazione grazie anche ad un bilanciamento della fitness dovuto ad un aumento della fecondita’ in linea materna queste basi biologiche spiegano tuttavia per il momento solo il 15% della varianza dell’orientamento sessuale

65
Che cosa succede per i bisessuali
Difficile identificarli, non emergono alla scala Kinsey Realizzare una nuova scala Discriminare meglio Omosessuali esclusivi e bisessuali Campione studiato 250 soggetti Replicazione studio precedente 98 htsx e 152 hmsx

66
Il nuovo campione secondo la scala Kinsey
(Mancano sempre i bisessuali!)
")
67
Anche per il nuovo campione si conferma, anche in modo piu’ marcato, la differenza di fecondita’ per le femmine della linea materna degli omosessuali

68
ordine di nascita del soggetto
L’ordine di nascita di omosessuali ed eterosessuali conferma l’ipotesi di Blanchard: ci sono significativamente piu’ eterosessuali primogeniti che omosessuali ordine di nascita del soggetto Primogeniti Secondogeniti Terzogeniti e più HTSX f. osservata 56 25 17 f. attesa 43,5 31,4 23,1 % entro gruppo 57,1 25,5 17,3 HMSX 55 42 67,5 48,6 35,9 36,2 27,6

69
numero di fratelli maggiori
Anche il numero di fratelli maschi maggiori e’ significativamente superiore negli omosessuali come previsto da Blanchard numero di fratelli maggiori Zero Uno Due o più HTSX f. osservata 77 16 5 f. attesa 63,1 23,9 11,0 % entro gruppo 78,6 16,3 5,1 HMSX 88 45 23 97,9 37,1 17,0 55,3 29,6 15,1

70
La nuova suddivisione di Htsx, Hmsx e Bsx: alla ricerca del Bisessuale
Autoidentificazione sulla scala Kinsey Variabili socioanagrafiche: Stato civile e paternita’ Occorrenza di rapporti sessuali con il genere sfavorito Possibilita’ di ripetere l’esperienza sessuale con il genere sfavorito in un contesto favorevole

71
La nuova classificazione
Eterosessuali Bisessuali Omosessuali Esclusi

72
Kruskal-Wallis test and Mann-Whitney test
Confronto con i bisessuali (Camperio Ciani, Blecher, Iemmola 2006) La linea discontinua indica approssimativamente il campione bisessuale Confronto della fecondita’ delle parenti in linea materna dei probandi **P<.01 **P<.01 *P<.03 Text at top right – change to: The discontinuous red line approximately indicates the bisexual sample. Re lower section of the slide: Upper case letters for first words in ordinate (Mean...) and abscissa (Mothers, First born etc. and Heterosexuals, Bisexuals etc. Correct spelling of Whitney. Also, this lower half needs a caption – it is not at all clear what these histograms are supposed to show – e.g. are the frequencies of htsx, bisx and hmsx in each histogram the offspring of, respectively, all mothers, first born mothers etc.? If so, I suggest a caption such as: Frequency of sexuality of offspring distributed by status of parents. And if this is what is meant, I’d suggest that the words “of probands” be inserted after Mothers, Maternal aunts and the next two. Kruskal-Wallis test and Mann-Whitney test Eterosessuali (N=86) Bisessuali (N=88) Omosessuali(N=65)
 La linea discontinua indica approssimativamente il campione bisessuale. Confronto della fecondita’ delle parenti in linea materna dei probandi. **P<.01. **P<.01. *P<.03. Text at top right – change to: The discontinuous red line approximately indicates the bisexual sample. Re lower section of the slide: Upper case letters for first words in ordinate (Mean...) and abscissa (Mothers, First born etc. and Heterosexuals, Bisexuals etc. Correct spelling of Whitney. Also, this lower half needs a caption – it is not at all clear what these histograms are supposed to show – e.g. are the frequencies of htsx, bisx and hmsx in each histogram the offspring of, respectively, all mothers, first born mothers etc. If so, I suggest a caption such as: Frequency of sexuality of offspring distributed by status of parents. And if this is what is meant, I’d suggest that the words of probands be inserted after Mothers, Maternal aunts and the next two. Kruskal-Wallis test and Mann-Whitney test. Eterosessuali (N=86) Bisessuali (N=88) Omosessuali(N=65)")
74
Replicazione su 448 soggetti italiani e francesi (Camperio Ciani & Iemmola 2006)
Upper case for first word in ordinate and abscissa. Correct the spelling of Whitney. Mann-Whitney test Eterosessuali (N=198) Omosessuali (N=250) Soggetti N=448
 Omosessuali (N=250) Soggetti N=448.")
75
Se i Bisessuali, esprimono fenotipicamente la propria identita’ di genere in maniera diversa dagli Omosessuali pur essendo influenzati dai fattori genetici che si comportano in maniera identica….. ….Quindi uno stesso genotipo puo’ produrre due fenotipi diversi, ovvero che l’espressione fenotipica viene influenzata dai geni ma modulata da altre cause esterne al genotipo (ambiente di sviluppo, pre- e post-natale, educazione, esperienze soggettive, non shared environment). Non esistono i geni dell’Omosessualita’ ma solo geni che influenzano l’identita’ di genere insieme ad altre cause ambientali. G A B O
. Non esistono i geni dell’Omosessualita’ ma solo geni che influenzano l’identita’ di genere insieme ad altre cause ambientali. G. A. B. O.")
76
Quanti sono, Dove si trovano e come si trasmettono questi geni?
Il modello matematico di diffusione e mantenimento deve soddisfare i seguenti requisiti empirici: Universalita’ Omosessuali presenti in tutte le popolazioni Bassa frequenza Omosessuali presenti ovunque con frequenze variabili ma sostanzialmente basse (.5 – 7 %) Asimmetria di orientamento Maggiore incidenza di omosessuali nella linea materna dei soggetti omosessuali Asimmetria di fecondita’ Fecondita’ maggiore nelle femmine in line materna dei soggetti omosessuali
 Asimmetria di orientamento Maggiore incidenza di omosessuali nella linea materna dei soggetti omosessuali. Asimmetria di fecondita’ Fecondita’ maggiore nelle femmine in line materna dei soggetti omosessuali.")
77
Modelli di selezione diretta, divergente progressiva e stabilizzante vengono esclusi subito perche’ non prevedono mantenimenti a basse frequenze ne asimmetrie di distribuzione fra sessi. I meccanismi selettivi che rimangono sono: 1)Overdominance 2)L’effetto materno 3) L’Imprinting genomico 4)Selezione sessualmente antagonistica Possiamo analizzare modelli: 1) ad un singolo locus 2) a piu’ loci (due per semplicita’)
Overdominance. 2)L’effetto materno. 3) L’Imprinting genomico. 4)Selezione sessualmente antagonistica. Possiamo analizzare modelli: 1) ad un singolo locus. 2) a piu’ loci (due per semplicita’)")
78
Quale modello genetico spiega meglio i risultati ottenuti sulle basi genetiche dell’omosessualita’?
Nei modelli a singolo locus: Tutti i modelli a singolo locus falliscono perche’ non rappresentano la stabilita’: o si estingue il tratto nella popolazione o in altre condizioni si diffonde in tutta la popolazione, contrariamente ai dati empirici di universalita’

79
Nei modelli a due o piu’ loci:
Il solo modello vincente e’ a selezione sessualmente antagonistica Con due fattori sessualmente antagonistici con vantaggio femminile uno sul cromosoma X l’altro indifferentemente (per ora) sul cromosoma X o sugli autosomi Questo modello soddisfa tutte le evidenze empiriche e ne suggerisce una nuova che e’ stata da noi poi verificata e confermata (le zie paterne degli eterosessuali sono piu’ feconde)! Tutti gli altri modelli falliscono perche’ non spiegano tutte le evidenze di universalita’ bassa frequenza e asimmetrie trovate.
 sul cromosoma X o sugli autosomi. Questo modello soddisfa tutte le evidenze empiriche e ne suggerisce una nuova che e’ stata da noi poi verificata e confermata (le zie paterne degli eterosessuali sono piu’ feconde)! Tutti gli altri modelli falliscono perche’ non spiegano tutte le evidenze di universalita’ bassa frequenza e asimmetrie trovate.")
80
Possibili meccanismi causali nelle femmine
Maggiore fertilita’ Minori patologie riproduttive o correlate ad esse Minori problemi di parto Diverso profilo di personalita’ Diverso approccio alla contraccezione Maggiore promisquita’

81
Possibili meccanismi attraverso i quali le donne di questo gruppo fanno piu’ figli
meccanismi fisiologici che riducono i problemi di gravidanza meccanismi che facilitano il parto aspetti di personalita’ come estroversione e scarsa coscenziosita’ diversi atteggiamenti e valori attribuiti all’amore ed alla famiglia maggiore predisposizione ai rapporti sessuali minore importanza attribuita alla contraccezione maggior numero di partner

82
Il nuovo campione di donne che hanno accettato di rispondere al questionario sulle cause della fecondita’ N 106, relative ad omosessuali 37 relative ad eterosessuali 69

83
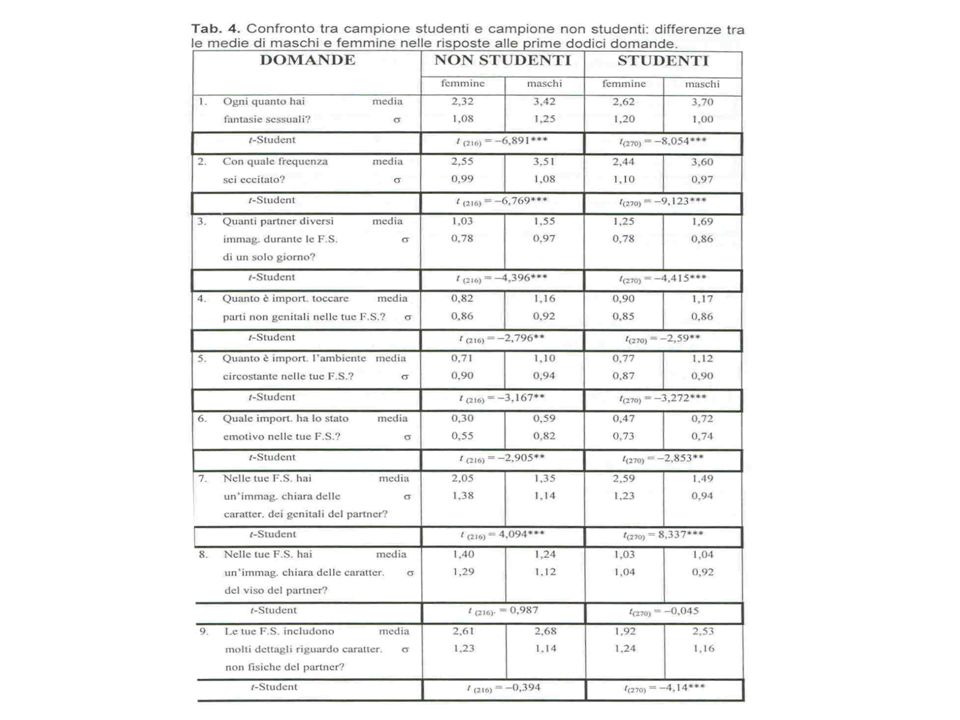
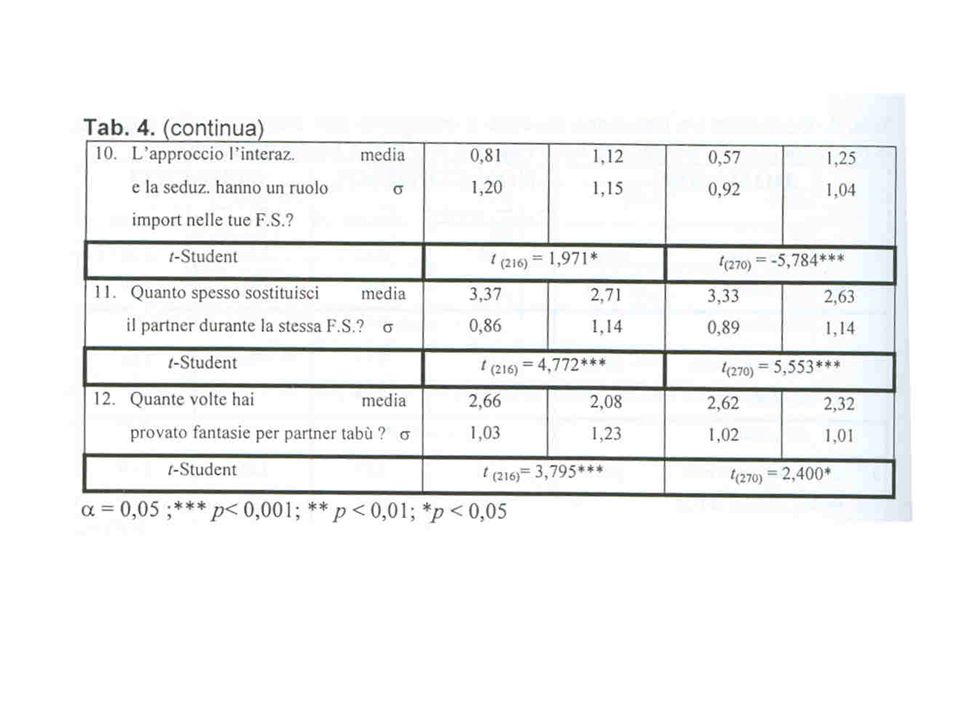
La fecondita’ media di madri e zie materne confrontata fra due nuovi gruppi di omosessuali ed eterosessuali nuovamente le indagini precedenti Parente Gruppo di controllo Gruppo sperimentale p N Fec. Media SD Madre 40 2.08 0.69 29 2.62 0.82 0.01 Zia materna 16 1.06 1.00 7 2.14 1.35 0.06 Madri + Zie materne 56 1.79 0.91 36 2.53 0.94 0.00

84
Le donne del gruppo sperimentale hanno meno disturbi di gravidanza in generale….
…Ma hanno piu’ disturbi legati al parto

85
I profili di personalita’ al BFQ ed al PDP non mostrano risultati significativi semmai inversi alle previsioni

86
numero medio di partner con cui si è avuta una relazione.
Valore del punteggio su scala Lickert Variabili indagate Gruppo sperimentale Gruppo di controllo Sig. (2-tailed ) P< α 0.05 N Media Dev st Numero di partner 32 2.13 1.58 68 1.54 0.92 .02 LEGENDA SCALA LICKERT: = da cinque a dieci 0 = zero partner 4 = da dieci a quindici 1 = un partner 5 = da quindici a venti 2 = da due a cinque 6 = più di venti partner
 P< α N. Media. Dev. st. Numero di partner LEGENDA SCALA LICKERT: 3 = da cinque a dieci. 0 = zero partner 4 = da dieci a quindici. 1 = un partner 5 = da quindici a venti. 2 = da due a cinque 6 = più di venti partner.")
87
Related Articles, Links
Heritable factors influence sexual orientation in women. Bailey JM, Pillard RC, Neale MC, Agyei Y. Arch Gen Psychiatry Mar;50(3): Department of Psychology, Northwestern University, Evanston, Ill Homosexual female probands with monozygotic cotwins, dizygotic cotwins, or adoptive sisters were recruited using homophile publications. Sexual orientation of relatives was assessed either by asking relatives directly, or, when this was impossible, by asking the probands. Of the relatives whose sexual orientation could be confidently rated, 34 (48%) of 71 monozygotic cotwins, six (16%) of 37 dizygotic cotwins, and two (6%) of 35 adoptive sisters were homosexual. Probands also reported 10 (14%) nontwin biologic sisters to be homosexual, although those sisters were not contacted to confirm their orientations. Heritabilities were significant using a wide range of assumptions about both the base rate of homosexuality in the population and ascertainment bias. The likelihood that a monozygotic cotwin would also be homosexual was unrelated to measured characteristics of the proband such as self-reported history of childhood gender nonconformity. Concordant monozygotic twins reported similar levels of childhood gender nonconformity.
: Department of Psychology, Northwestern University, Evanston, Ill Homosexual female probands with monozygotic cotwins, dizygotic cotwins, or adoptive sisters were recruited using homophile publications. Sexual orientation of relatives was assessed either by asking relatives directly, or, when this was impossible, by asking the probands. Of the relatives whose sexual orientation could be confidently rated, 34 (48%) of 71 monozygotic cotwins, six (16%) of 37 dizygotic cotwins, and two (6%) of 35 adoptive sisters were homosexual. Probands also reported 10 (14%) nontwin biologic sisters to be homosexual, although those sisters were not contacted to confirm their orientations. Heritabilities were significant using a wide range of assumptions about both the base rate of homosexuality in the population and ascertainment bias. The likelihood that a monozygotic cotwin would also be homosexual was unrelated to measured characteristics of the proband such as self-reported history of childhood gender nonconformity. Concordant monozygotic twins reported similar levels of childhood gender nonconformity.")
88
Influenza genetica nell'orientamento sessuale (Bailey)
Differenze nella distribuzione generale dei non -eterosessuali Gay e lesbiche: la sessualita' senza senza riproduzione perche' esiste? Influenza genetica nell'orientamento sessuale (Bailey) Teoria sintetica dell'evoluzione, e dell'adattamento: I caratteri genetici che non mantengono il successo riproduttivo dei portatori sono destinati a sparire. Il paradosso Darwiniano: Lesbiche e gay ci sono da sempre! Distribution of Kinsey scores for all men ( = 147) and women ( = 238) with nonzero Kinsey scores (i.e., who were not exclusively heterosexual), as their percentage of the entire within-sex sample: 1 = heterosexual with slight homosexual feelings; 2 =heterosexual with substantial homosexual feelings; 3 =equally heterosexual and homosexual; 4 =homosexual with substantial heterosexual feelings; 5 =homosexual with slight heterosexual feelings; 6 = completely homosexual. Bailey,2000: J Pers Soc Psychol, 78(3).524–536.
 Teoria sintetica dell evoluzione, e dell adattamento: I caratteri genetici che non mantengono il successo riproduttivo dei portatori sono destinati a sparire. Il paradosso Darwiniano: Lesbiche e gay ci sono da sempre! Distribution of Kinsey scores for all men ( = 147) and women ( = 238) with nonzero Kinsey scores (i.e., who were not exclusively heterosexual), as their percentage of the entire within-sex sample: 1 = heterosexual with slight homosexual feelings; 2 =heterosexual with substantial homosexual feelings; 3 =equally heterosexual and homosexual; 4 =homosexual with substantial heterosexual feelings; 5 =homosexual with slight heterosexual feelings; 6 = completely homosexual. Bailey,2000: J Pers Soc Psychol, 78(3).524–536.")
89
Patterson,2005, Camperio Ciani,2012
Soluzione: Soluzioni diverse per I due sessi: I gay fanno meno figli ma gli stessi fattori genetici che influenzano l'orientamento sessuale fanno fare piu' figli alle femmine che li portano (Selezione sessualmente antagonistica Camperio Ciani) Nelle lesbiche il successo riproduttivo cambia ma rimane equivalente perche' alla carente spinta erotica etero-sessuale si sostituisce l'istinto materno (Patterson, Camperio Ciani). Soluzione: l'omosessualita' evidenzia che gay e lesbiche sono due fenomeni apparentemente simili ma in realta' completamente distinti I gay esistono perche' i fattori che riducono la fecondita' dei gay maschi aumentano quella delle loro femmine materne. e le lesbiche esistono perche' il loro comportamento, in effetti, non riduce la loro fecondita'. Con buona pace per coloro che ancora credono che l'omosessualita' sia una malattia o un vizio da curare!! Etero n=198 Omo n=250 (Camperio Ciani et al, 2008,2013) Patterson,2005, Camperio Ciani,2012
 Nelle lesbiche il successo riproduttivo cambia ma rimane equivalente perche alla carente spinta erotica etero-sessuale si sostituisce l istinto materno (Patterson, Camperio Ciani). Soluzione: l omosessualita evidenzia che gay e lesbiche sono due fenomeni apparentemente simili ma in realta completamente distinti I gay esistono perche i fattori che riducono la fecondita dei gay maschi aumentano quella delle loro femmine materne. e le lesbiche esistono perche il loro comportamento, in effetti, non riduce la loro fecondita . Con buona pace per coloro che ancora credono che l omosessualita sia una malattia o un vizio da curare!! Etero n=198. Omo n=250 (Camperio Ciani et al, 2008,2013) Patterson,2005, Camperio Ciani,2012.")
90
Quindi…. L’orientamento sessuale non modifica completamente le caratteristiche dell’identita’ del genere biologico esistono delle basi genetiche e fisiologiche dell’omosessualita’ che si mantengono nella popolazione grazie anche ad un bilanciamento della fitness queste basi biologiche spiegano tuttavia solo il 15-20% della varianza dell’orientamento sessuale Le cause dell’aumento della fecondita’ nelle femmine di linea materna sono da approfondire ancora .

91
Conclusioni Secondo questi studi, l’omosessualita’ non e’ una malattia bensi’ l’effetto di una strategia genetica sessualmente antagonistica che ha come effetto finale l’aumento della fecondita’. Le componenti genetiche finora identificate dell’omosessualita’ e della bisessualita’sono solo una piccola parte di un quadro complesso di interazioni fra aspetti genetici e modulazioni ambientali che producono l’identita’ di genere sia sulle componenti genetiche che sugli effetti dell’ambiente molto ancora resta da capire Per quanto riguarda l’espressione dei geni che conducono all’omosessualita’ e come funziona l’Omosessualita’ femminile…. …Ve lo racconto la prossima volta. Grazie!

Presentazioni simili

 Dominanza incompleta>")

















