Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
consisting of a long α he- lix (α2) flanked on each side by a random coil loop (L1 or L2) and a short α helix (α1 or α3), long α helix (α2) flanked on each side by a random coil loop (L1 or L2) and a short α helix (α1 or α3),
 flanked on each side by a random coil loop (L1 or L2) and a short α helix (α1 or α3),")
3
HISTONE VARIANTS H2A.Z has apparently contradictory roles in gene activation and silencing; nucleosome turnover; DNA repair; heterochromatin H2A.Z differs from H2A and around the L1–α2 and α2–L2 junctions and in the C-terminal 'docking domain' that contacts H3 H2A.Z-containing nucleosomes are found on either side of a 'nucleosome-free region' (NFR) at transcription start sites (TSSs)6, where they promote efficient RNA polymerase II (RNAPII) recruitment in both yeast and human cell H2A.Z function are located in its C-terminus interaction of H2A.Z with the evolutionary conserved SWR-1 (Swi2/Snf2-related ATPase-1) chromatin remodelling complex important for H2A.Z H2A.Z is present at most promoters in yeast whether they are active promoters or not6. in uomo H2A.Z is found at enhancers and in heterochromatin and its presence at promoters correlates with RNA polymerase II (RNAPII) occupancy, suggesting it is only recruited to promoters when needed Saccharomyces cerevisiae H2A.Z (Htz1) is enriched on repressed promoters, whereas Htz1 acetylated on Lys14 is enriched on active promoters H2A.Z in human heterochromatic regions is under-acetylated. Acetylation of H2A.Z is necessary for its association with inducible promoters increased acidic patch(interazione con altri istoni) on its surface is observed for the H2A.Z nucleosome, suggesting an influence on internucleosomal interactions Possible implications of the H2A.Z ubiquitination correlates with localization to the inactive X chromosome (Xi) in mammals , whereas N-terminal acetylation leads to nucleosome destabilization H2A.Z has apparently contradictory roles in gene activation and silencing; nucleosome turnover; DNA repair; heterochromatin
 at transcription start sites (TSSs)6, where they promote efficient RNA polymerase II (RNAPII) recruitment in both yeast and human cell. H2A.Z function are located in its C-terminus interaction of H2A.Z with the evolutionary conserved SWR-1 (Swi2/Snf2-related ATPase-1) chromatin remodelling complex important for H2A.Z. H2A.Z is present at most promoters in yeast whether they are active promoters or not6. in uomo H2A.Z is found at enhancers and in heterochromatin and its presence at promoters correlates with RNA polymerase II (RNAPII) occupancy, suggesting it is only recruited to promoters when needed. Saccharomyces cerevisiae H2A.Z (Htz1) is enriched on repressed promoters, whereas Htz1 acetylated on Lys14 is enriched on active promoters. H2A.Z in human heterochromatic regions is under-acetylated. Acetylation of H2A.Z is necessary for its association with inducible promoters. increased acidic patch(interazione con altri istoni) on its surface is observed for the H2A.Z nucleosome, suggesting an influence on internucleosomal interactions. Possible implications of the H2A.Z ubiquitination correlates with localization to the inactive X chromosome (Xi) in mammals. , whereas N-terminal acetylation leads to nucleosome destabilization. H2A.Z has apparently contradictory roles in gene activation and silencing; nucleosome turnover; DNA repair; heterochromatin.")
4
Histone fold proteins H2A H2B NF-YB NF-YC NC2b NC2a YEATS2 Pole4
CHRAC14 CHRAC16 Pole3 TAF12 NF-Y fattore trascizionale NC2 repressore Remodelling complex chrac che oltre ad una subunita’ iswi TAF TBP associated factor che compongono I vari TFs (accessory factors)che sono GTFs che interagiscono con tbp per legare TATAbox e attivare trascriz consisting of a long α he- lix (α2) flanked on each side by a random coil loop (L1 or L2) and a short α helix (α1 or α3), TAF4 TAF4b TAF3 TAF10 TAF11 TAF13 H3 H4 TAF9 TAF6 TAF6L
che sono GTFs che interagiscono con tbp per legare TATAbox e attivare trascriz. consisting of a long α he- lix (α2) flanked on each side by a random coil loop (L1 or L2) and a short α helix (α1 or α3), TAF4. TAF4b. TAF3. TAF10. TAF11. TAF13. H3. H4. TAF9. TAF6. TAF6L.")
5
TFIID Histon foldi protein fanno parte di complessi che servono per reclutare I GTF insieme a TBP sul tata box vicino al tSS Si legano a certe sequenze caratteristiche del core promoter TBP o TBP subunita del TFIID si lega alla tata box TFIID che contiene TAF1 e TAF2 si legano alla seq inr(initiator element) sul tss che con tbp o meno fanno partire la trascriz TAF9-TAF6 riconoscono la sequenza DPE(distal promoter element) BRE element riconosciuto da TFIIB indipendentemente da TBP nel primo e dipendente nel secondoaiuta TFIID DCE downstream core element dal TAF1
 sul tss che con tbp o meno fanno partire la trascriz. TAF9-TAF6 riconoscono la sequenza DPE(distal promoter element) BRE element riconosciuto da TFIIB indipendentemente da TBP nel primo e dipendente nel secondoaiuta TFIID. DCE downstream core element dal TAF1.")
6
In general, TFIIA and TFIIB enhance TBP binding to the TATA box and further stabilize the TBP-promoter complex via distinct mechanisms TFIIA increases the likelihood of precise promoter recogni- tion by TBP via promoting the dissociation of TBP dimers and thus facilitating the loading of monomeric TBP onto the TATA box (Coleman et al., 1999; see Figure 5B). TFIIA also competes with the inhibitory do- main of TAF1 for overlapping binding regions on TBP, hence alleviating TAF1-mediated inhibition of TATA recognition (Kokuba et al., 1998). Furthermore, incor- poration of TFIIA into the TBP-TATA complex renders the ternary complex resistant to BTAF1-mediated dis- sociation of the TBP-TATA complex (Auble and Hahn, 1993). Similarly, TFIIB can enhance TBP binding to the TATA box (Imbalzano et al., 1994b) and also sta- bilize the bent TBP-TATA complex (Zhao and Herr, 2002), thereby reducing the dissociation rate of TBP from the promoter region (Wolner and Gralla, 2 Besides protein-protein interaction observed between TFIIB and TBP, additional DNA contacts provided by TFIIB-BREu and TFIIB-BREd likely contribute to this stabilizing effect (Figure 5C). chromatin-modifying en- zymes play a key role in enhancing TBP access to its recognition sequence by altering chromatin structure surrounding the TATA box
. TFIIA also competes with the inhibitory do- main of TAF1 for overlapping binding regions on TBP, hence alleviating TAF1-mediated inhibition of TATA recognition (Kokuba et al., 1998). Furthermore, incor- poration of TFIIA into the TBP-TATA complex renders the ternary complex resistant to BTAF1-mediated dis- sociation of the TBP-TATA complex (Auble and Hahn, 1993). Similarly, TFIIB can enhance TBP binding to the TATA box (Imbalzano et al., 1994b) and also sta- bilize the bent TBP-TATA complex (Zhao and Herr, 2002), thereby reducing the dissociation rate of TBP from the promoter region (Wolner and Gralla, 2. Besides protein-protein interaction observed between TFIIB and TBP, additional DNA contacts provided by TFIIB-BREu and TFIIB-BREd likely contribute to this stabilizing effect (Figure 5C). chromatin-modifying en- zymes play a key role in enhancing TBP access to its recognition sequence by altering chromatin structure surrounding the TATA box.")
7
NC2-TBP-TATA NC2b NC2a A third TBP-interacting factor able to regulate TBP binding to the promoter region is negative cofactor 2 (NC2), which is conserved in eukaryotes Human NC2 consists of NC2α (Drap1; 22 kDa) and NC2β (Dr1; 20 kDa) that interact with each other through histone fold motifs, resembling the dimerization domain found in histones H2A and H2B The structural analysis of NC2-TBP-TATA ternary complex,, shows that the N-terminal regions of both NC2 subunits bind DNA on the underside of the preformed TBP-TATA complex and the C-terminus of NC2β additionally contacts the convex surface of TBP, hence giving NC2 the appearance of a molecular “clamp” that grips both upper and lower surfaces of the TBP-TATA complex this molecular clamp is able to block PIC assembly by inhibiting TFIIA and TFIIB binding to the upper side of TBP NC2 can also function as a coactivator in stimulating transcription from some TATA-less promoters NC2 works in conjunction with TFIID through the DPE to enhance TATA-less gene transcription
, which is conserved in eukaryotes. Human NC2 consists of NC2α (Drap1; 22 kDa) and NC2β (Dr1; 20 kDa) that interact with each other through histone fold motifs, resembling the dimerization domain found in histones H2A and H2B. The structural analysis of NC2-TBP-TATA ternary complex,, shows that the N-terminal regions of both NC2 subunits bind DNA on the underside of the preformed TBP-TATA complex and the C-terminus of NC2β additionally contacts the convex surface of TBP, hence giving NC2 the appearance of a molecular clamp that grips both upper and lower surfaces of the TBP-TATA complex. this molecular clamp is able to block PIC assembly by inhibiting TFIIA and TFIIB binding to the upper side of TBP. NC2 can also function as a coactivator in stimulating transcription from some TATA-less promoters. NC2 works in conjunction with TFIID through the DPE to enhance TATA-less gene transcription.")
8
Repression by NC2 NC2 binding precludes recruitment of TFIIB and TFIIA to a preformed TBP-DNA complex. Molecular surface representations of NC2 (blue mesh), TFIIB (solid gray), and TFIIA (red mesh) bound to TBP (green) and DNA (yellow/orange ribbons). NC2 recognizes the pre- formed TBP-TATA element binary complex, making sig- nificant protein-protein contacts with both domains of TBP (sites 1 and 2; Figure 2) that appear to preclude recruitment of both TFIIB and TFIIA to the core promoter.
, TFIIB (solid gray), and TFIIA (red mesh) bound to TBP (green) and DNA (yellow/orange ribbons). NC2 recognizes the pre- formed TBP-TATA element binary complex, making sig- nificant protein-protein contacts with both domains of TBP (sites 1 and 2; Figure 2) that appear to preclude recruitment of both TFIIB and TFIIA to the core promoter.")
9
TFIID TAF1 Histone acetylation TAF12 TAF4 TAF4b TAF9 TAF6 TAF3 TAF10
four of them (TAF6, TAF9, TAF12, and TAF4/TAF4b) share sequence similarity to core histones H4, H3, H2B, and H2A, respectively (Gangloff et al., 2001). A histone fold motif, is organized in the following order: the histone folds in TAFs are likely to be important for protein-protein interactions for maintaining the structural integrity of TFIID, TAF1 in TFIID not only modulates DNA binding by TBP, but also confers on TFIID multiple enzymatic activities to post-translationally modify histones and transcription factors, thus allowing TFIID to serve as a core promoter-binding factor in the con- text of chromatin and as a coactivator TAF1, a TFIID-specific subunit, is known to function as a histone acetyltransferase (HAT) in acetylating histones H3 and H4 in vitro TAF1 has also been shown to act as a kinase in phosphorylating histone H2B (Maile et al., 2004) and as a histone-specific ubiquitin-activating/conjugating en- zyme in mediating monoubiquitination of linker hi- stone H1 TAF12 TAF4 TAF4b TAF9 TAF6 TAF3 TAF10 TAF11 TAF13
 share sequence similarity to core histones H4, H3, H2B, and H2A, respectively (Gangloff et al., 2001). A histone fold motif, is organized in the following order: the histone folds in TAFs are likely to be important for protein-protein interactions for maintaining the structural integrity of TFIID, TAF1 in TFIID not only modulates DNA binding by TBP, but also confers on TFIID multiple enzymatic activities to post-translationally modify histones and transcription factors, thus allowing TFIID to serve as a core promoter-binding factor in the con- text of chromatin and as a coactivator. TAF1, a TFIID-specific subunit, is known to function as a histone acetyltransferase (HAT) in acetylating histones H3 and H4 in vitro. TAF1 has also been shown to act as a kinase in phosphorylating histone H2B (Maile et al., 2004) and as a histone-specific ubiquitin-activating/conjugating en- zyme in mediating monoubiquitination of linker hi- stone H1. TAF12. TAF4. TAF4b. TAF9. TAF6. TAF3. TAF10. TAF11. TAF13.")
10
TAF12 and TAF6 H2B-H4 interface
reconstruction of the interface between the hTAF4-hTAF12(H2A-H2B) and hTAF6- hTAF9 (H3-H4)complexes, modeled so as to maximize resemblance to the corresponding region in the structure of the nucleosome core particle (36). For clarity, only hTAF12 and hTAF6 are shown. Hydrophobic residues contributing to the interface are shown in green as a result of a difference in “twist” between the H3-H4 and TAF6-TAF9 heterodimers (42), the resulting octamer features a 20 Å gap (arrow) between the L1 loops of the two hTAF4-hTAF12 dimers, and forms an open spiral rather than a compact nucleosome-like particle. Because the structural similarity of the two TAFs to the corresponding histones is particularly strong in the interface regions (i.e. the 2-L2- 3 regions of TAF12 and TAF6), the resulting TAF-TAF interface (Fig. 5A) is remarkably similar to the H2B-H4 inter- face. Conserved hydrophobic residues occupy virtually identi- cal relative positions to the corresponding residues in the his- tone octamer. Thus, a cluster of planar side chains (His-105 of hTAF12, Phe-57, and Tyr-73 of hTAF6) can be formed that is highly similar to the aromatic core of the H2A-H4 interface. Additional hydrophobic residues (Leu-102 and Leu-118 of hTAF12, Met-60 and Leu-77 of hTAF6) occupy positions equiv- alent to similar or identical residues in H2A and H4.
 and hTAF6- hTAF9 (H3-H4)complexes, modeled so as to maximize resemblance to the corresponding region in the structure of the nucleosome core particle (36). For clarity, only hTAF12 and hTAF6 are shown. Hydrophobic residues contributing to the interface are shown in green. as a result of a difference in twist between the H3-H4 and TAF6-TAF9 heterodimers (42), the resulting octamer features a 20 Å gap (arrow) between the L1 loops of the two hTAF4-hTAF12 dimers, and forms an open spiral rather than a compact nucleosome-like particle. Because the structural similarity of the two TAFs to the corresponding histones is particularly strong in the interface regions (i.e. the 2-L2- 3 regions of TAF12 and TAF6), the resulting TAF-TAF interface (Fig. 5A) is remarkably similar to the H2B-H4 inter- face. Conserved hydrophobic residues occupy virtually identi- cal relative positions to the corresponding residues in the his- tone octamer. Thus, a cluster of planar side chains (His-105 of hTAF12, Phe-57, and Tyr-73 of hTAF6) can be formed that is highly similar to the aromatic core of the H2A-H4 interface. Additional hydrophobic residues (Leu-102 and Leu-118 of hTAF12, Met-60 and Leu-77 of hTAF6) occupy positions equiv- alent to similar or identical residues in H2A and H4.")
11
TAF11 TAF13

12
Fig 2 Schematic model for TFIID assembly. A The histone fold containing TAFs form heterodimers and associates with TAF5 to form the core complex. The ability of TAF5 to homodimerise is still open to question, b TAF5 and the histone-like heterodimers associate to form the core complex. c The core complex interacts with the TAF1-TAF7-TBP sub-module to form TFIID. TAF2 may associate independently with TFIID through interaction with TAF1, but may not be always present in TFIID Altogether, the higher resolution structure and the labelling studies strongly suggest that the 3D architecture of TFIID is composed of two subcomplexes: (1) a core complex containing TAF5 and most of the HFD-containing TAF heterodimers (TAF6/9, TAF4/12, TAF8/10, TAF11/13) that adopts a crescent-shaped twofold symmetric structure; (2) a subcomplex containing TAF1, TAF7, TAF2 and TBP that is recruited to the core complex to form full TFIID (Fig 1 Structural organisation of yeast TFIID; (a, b) two opposite views of the TFIID complex showing the location of TAF1, TAF7 and TBP (white), TAF2 (blue) and a quasi symmetric core module (red). c Schematic representation of the TAF localisation within TFIID. The approximate positions of the subunits are derived from antibody-labelling experiments. The size of the spheres is proportional to the molecular mass of the proteins. Black lines represent documented protein–protein interactions of histone fold domain containing TAFs
 a core complex containing TAF5 and most of the HFD-containing TAF heterodimers (TAF6/9, TAF4/12, TAF8/10, TAF11/13) that adopts a crescent-shaped twofold symmetric structure; (2) a subcomplex containing TAF1, TAF7, TAF2 and TBP that is recruited to the core complex to form full TFIID (Fig 1 Structural organisation of yeast TFIID; (a, b) two opposite views of the TFIID complex showing the location of TAF1, TAF7 and TBP (white), TAF2 (blue) and a quasi symmetric core module (red). c Schematic representation of the TAF localisation within TFIID. The approximate positions of the subunits are derived from antibody-labelling experiments. The size of the spheres is proportional to the molecular mass of the proteins. Black lines represent documented protein–protein interactions of histone fold domain containing TAFs.")
13
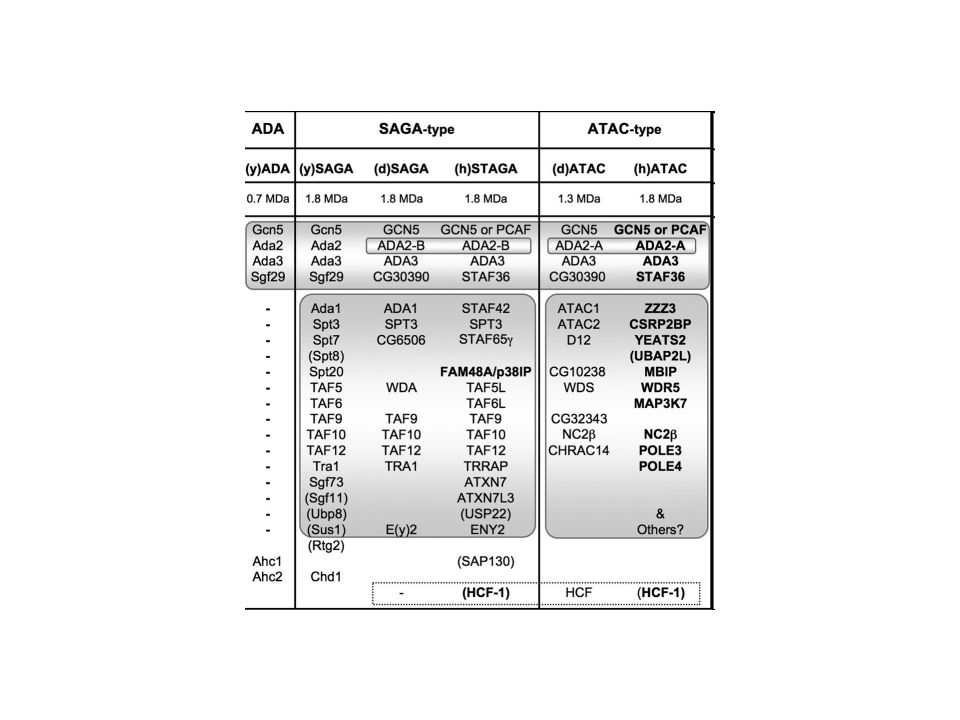
SAGA Histone acetylation ? PCAF GCN5 TAF4 TAF12 TAF10 TAF9 TAF6L
Spt-Ada-Gcn5-acetyltransferase TAF12 TAF4 The Taf (TBP-associated factor) subunits, shared with TFIID, occupy a central region in SAGA and form a similar structure in both complexes. The locations of two histone fold-containing core subunits, Spt7 and Ada1, are consistent with their role in providing a SAGA-specific interface with the Tafs. Three components that perform distinct regulatory functions, Spt3, Gcn5, and Tra1, are spatially separated, underscoring the modular nature of the complex. Our data provide insights into the molecular architecture of SAGA and imply a functional organization to the complex. This complex was therefore named SAGA (Spt-Ada-Gcn5-acetyltransferase). The Spt proteins known to be present in the SAGA complex include Spt3, Spt7, Spt8, and Spt20. Spt7 (20) and Spt20 (60) are apparently vital to the structure of SAGA ? TAF10 TAF9 TAF6L
 subunits, shared with TFIID, occupy a central region in SAGA and form a similar structure in both complexes. The locations of two histone fold-containing core subunits, Spt7 and Ada1, are consistent with their role in providing a SAGA-specific interface with the Tafs. Three components that perform distinct regulatory functions, Spt3, Gcn5, and Tra1, are spatially separated, underscoring the modular nature of the complex. Our data provide insights into the molecular architecture of SAGA and imply a functional organization to the complex. This complex was therefore named SAGA (Spt-Ada-Gcn5-acetyltransferase). The Spt proteins known to be present in the SAGA complex include Spt3, Spt7, Spt8, and Spt20. Spt7 (20) and Spt20 (60) are apparently vital to the structure of SAGA. TAF10. TAF9. TAF6L.")
14
ATAC Histone acetylation GCN5 ATAC2 NC2b YEATS2 Pole4 CHRAC14 Pole3
two acetyltransferases GCN5 and ATAC2 ATAC2 is depleted display reduced levels of acetylated histones H3 and H4 at specific lysines GCN5 ATAC2 Histone acetylation

16
hSNF2/ACF1 Nucleosome sliding hCHRAC Pole4 CHRAC16 CHRAC14
Iswi subunit SNF2 / ACF1rimodellatore della cromatina CHRAC14 CHRAC16 dimerizzano e interagiscono con il dna con histon fold hSNF2/ACF1 hCHRAC Nucleosome sliding

17
DNA Pol e DNA repair Pole4 CHRAC14 Pole3
Pol4 e pole 3 favoriscono interazione dna durante il proceso di replicazione CHRAC14 e DPB3 DNA Pol e DNA repair

18
ACF/ CHRAC DNA Pol e hACF1 hSNF2H DPB1 DPB2 DPB3 CHRAC14 CHRAC14

19
NF-Y NF-YB NF-YC

20
The crystal structure of NF-Y bound to a 25 bp CCAAT oligonucleotide shows that the HFD dimer binds to the DNA sugar-phosphate backbone, mimicking the nucleosome H2A/H2B-DNA assembly. NF-YA both binds to NF-YB/NF-YC and inserts an α helix deeply into the DNA minor groove, providing sequence-specific contacts to the CCAAT box Structural considerations and mutational data indicate that NF-YB ubiquitination at Lys138 precedes and is equivalent to H2B Lys120 monoubiquitination, important in transcriptional activation. Thus, NF-Y is a sequence-specific transcription factor with nucleosome-like properties eliminandolo si ha la scomparsa di alcune modificazioni istoniche attivtorie (H3K4me3 e H3K79me2) pioneer factor
 pioneer factor.")
21
TAF1 TAF7 TAF2 TAF5 NF-YA NF-YB NF-YC TAF12 TAF4 TAF10 TAF3 TAF9 TAF11
TBP Interagisce con molti gtf,tbp e coattivatori come pcaf e p300 TAF9-TAF6 interagiscono con il trimero TAF12,11,13 con il dimero yb-yc TAF13 NF-YA NF-YB NF-YC

24
Stem Cells. 2012 Nov;30(11):2450-9. doi: 10.1002/stem.1232.
Cell Jan 17;152(1-2): doi: /j.cell Sequence-specific transcription factor NF-Y displays histone-like DNA binding and H2B-like ubiquitination. Nardini M, Gnesutta N, Donati G, Gatta R, Forni C, Fossati A, Vonrhein C, Moras D, Romier C, Bolognesi M, Mantovani R. Source Dipartimento di BioScienze, Università degli Studi di Milano, Via Celoria 26, Milano, Italy Stem Cells Nov;30(11): doi: /stem.1232. The short isoform of NF-YA belongs to the embryonic stem cell transcription factor circuitry. Dolfini D, Minuzzo M, Pavesi G, Mantovani R. Source Dipartimento di Scienze Biomolecolari e Biotecnologie, Università degli Studi di Milano, Milano, Italy.
: doi: /j.cell Sequence-specific transcription factor NF-Y displays histone-like DNA binding and H2B-like ubiquitination. Nardini M, Gnesutta N, Donati G, Gatta R, Forni C, Fossati A, Vonrhein C, Moras D, Romier C, Bolognesi M, Mantovani R. Source. Dipartimento di BioScienze, Università degli Studi di Milano, Via Celoria 26, Milano, Italy. Stem Cells Nov;30(11): doi: /stem The short isoform of NF-YA belongs to the embryonic stem cell transcription factor circuitry. Dolfini D, Minuzzo M, Pavesi G, Mantovani R. Source. Dipartimento di Scienze Biomolecolari e Biotecnologie, Università degli Studi di Milano, Milano, Italy.")
Presentazioni simili


 Brussels, 26 settembre 2013.>")





 physician who has completed training as an intern and resident and has been granted a stipend and position allowing him or her to do further.>")












